рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Философия
- /
- Эволюционный ряд основных типов морфологической симметрии организмов.
Реферат Курсовая Конспект
Эволюционный ряд основных типов морфологической симметрии организмов.
Эволюционный ряд основных типов морфологической симметрии организмов. - раздел Философия, Эволюционные теории асимметризации организмов, мозга и тела Первые Организмы На Земле, Плавающие В Толще Воды Одноклеточные, Имели Максим...
Первые организмы на Земле, плавающие в толще воды одноклеточные, имели максимально возможную симметрию – шаровую, они появились примерно 3,5 млрд. лет назад. Спустя 2,5 млрд. лет возник половой процесс (оплодотворение). Это привело к вспышке разнообразия и почти одновременному появлению простейших многоклеточных ШС и малоподвижных или прикрепленных организмов радиальнойсимметрии. Следующие 80 млн. лет на Земле превалировала удивительная и своеобразная фауна Венда, характерные черты которой: многоклеточность, отсутствие скелета, гигантизм и радиальная симметрия. Это были двухслойные дисковидные организмы диаметром до 1,5 м. Билатеральносимметричные организмы, занимавшие в фауне Венда скромное место, господствуют последние 650–800 млн. лет. Это ракообразные, рыбы, все прогрессивные формы: млекопитающие, птицы, насекомые.
4.1. Симметрия и эволюционный прогресс.
Организмышаровой, радиальной и билатеральной симметрии широко распространены в животном мире и хорошо известны. Симметрия в этом ряду закономерно падает, а асимметрия и эволюционная прогрессивность – растут. В.Н. Беклемишев, считая амёбу полностью асимметричным организмом, более примитивным, чем одноклеточные ШС (радиолярии, вольвоксовые), поместил ее в начале ряда. Стало четыре типа: амёба (полная асимметрия), ШС, РС, БС (Беклемишев, 1964). Я высказал сомнение в правильности такой концепции и предложил новую (рис. 2) (Геодакян, 1993).

Если по каждой из осей трехмерного пространства симметрию организма обозначить с, асимметрию – а, то теоретически возможны четыре типа симметрии: ссс, асс, аас, ааа. Первые три, по времени появления в природе и возрастающей прогрессивности форм, укладываются в закономерный эволюционный ряд: ШС®РС®БС. Четвертый тип (ааа) – триаксиальной асимметрии – либо отсутствовал, либо его помещали в начале ряда. В любом случае “венцом” эволюции считали билатеральную симметрию. Число типов Беклемишев угадал правильно, но 4-ый тип определил и поместил неправильно. В чем его ошибки? 1. Симметрия–асимметрия признаки формы, а амёба – бесформенна и ей не место в классификации форм. 2. Триаксиальная асимметрия означает, что у организма есть разные (невзаимозаменяемые) спина–брюхо, голова–хвост, левая–правая сторона тела. У амёбы их нет. 3. Предложенный им ряд: ааа®ссс®асс ®аас, с “революционным” возникновением из полной асимметрии абсолютной симметрии и дальнейшей, уже эволюционной, её неполной утратой, алогичен (трудно представить результатом естественной эволюции немонотонный ряд для числа осей асимметрии: 3®0®1®2, или ряд геометрических понятий: объем®точка®линия®поверхность!). Странно, что с ним согласился, верно, с оговорками, такой блестящий теоретик, как А.А.Любищев (1982).
Согласно же эволюционной логике, тип ТА должен стоять не в начале, а в конце ряда. И это должны быть не самые примитивные организмы, а наоборот, самые прогрессивные, но в отличие от трех более древних и простых типов, широко распространенных на Земле, тип ТА новый, эволюционно самый молодой, поэтому пока еще редкий и неприметный. Он появляется на наших глазах, но это – закономерное будущее БС-организмов.
Латерализация начинается, видимо, у плацентарных млекопитающих, с появлением мозолистого тела(у опоссума его нет). Асимметрия максимально развита у человека, и, судя по половому диморфизму (Геодакян, 1980,1984,1987,1992,1993,1997,1998), продолжает расти. Следовательно, по асимметрии, венцом эволюции нужно считать не БС, а ТА. Тогда асимметризация организмов предстает единым, закономерно-прогрессивным процессом, по последовательному освоению асимметрией всех осей трехмерного пространства: 0®1®2®3 (рис. 3).
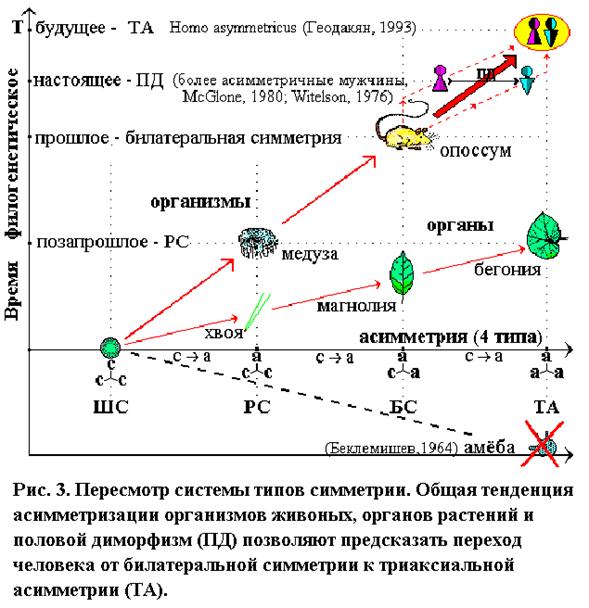
Таким образом, экстраполяция эволюционной логики существующего ряда: ссс®асс®аас; растущее число фактов латеральной асимметрии у высших форм (ФАМ и видовая праворукость человека, односторонняя овуляция и однополушарный сон дельфинов); тесные связи асимметрии с полом и направление полового диморфизма; позволили высказать гипотезу о дальнейшей асимметризации современных прогрессивных форм и закономерном их переходе от БС к последнму типу ТА (аас®ааа) такого же фундаментального эволюционного ранга, высшей (и по асимметрии, и по прогрессивности). Нас интересует именно этот переход. Предвижу, моя футуристическая гипотеза не понравится многим по чисто эстетическим соображениям. Но ведь вкусы тоже эволюционируют. И для “триаксиальных” вкусов, “билатеральная” красота, будет казаться несколько старомодной, как нам “радиальная” красота медузы.
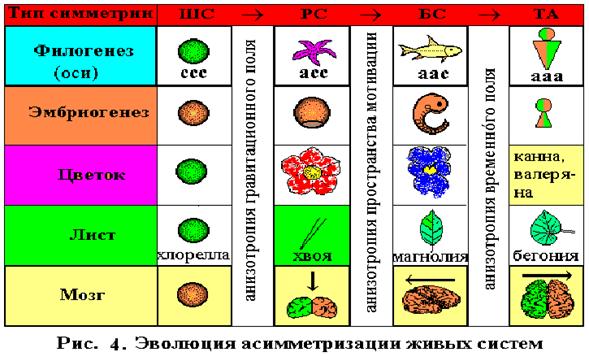
Та же тенденция наблюдается и в филогенезе органов растений(цветок, лист, плоды, семена), и вряд ли это случайно. Например, известно, что зигоморфные (БС) цветкидельфиниума и львиного зева, эволюционно более прогрессивны, чем актиноморфные (РС) цветки розы и лютика, но менее прогрессивны, чем ТА цветки канны и валерьяны. Ту же самую картину имеем и с морфологией листа в процессе эволюции: ШС хлореллы (почки), РС хвои, БС листа магнолиевых и ТА листа вязовых или бегониевых. Интересно отметить, что асимметрия органов растений более продвинута, чем организмов животных. Если существуют двудомные триаксиально асимметричные растения, то теория предсказывает у них наличие полового диморфизма по всем признакам, имеющим латеральный (правило сопряженных диморфизмов).
Вскрытая тенденция асимметризации в филогенезе, согласно закону рекапитуляции, прослеживается и в эмбриогенезе: ШС зиготы®РС гаструлы®БС эмбриона®ТА ребенка. Было показано, что в утробе матери эмбрион сосет большой палец (Hepper et al., 1991).
4.2. Асимметризация организмов – ответ на анизотропизацию среды..
В рамках адаптогенеза Дарвина признаки симметрии, как и все другие, определяются условиями среды. Поэтому развитие асимметрии в процессе земной эволюции связано с анизотропией среды обитания (см. рис. 3, 4). Асимметризация по первой оси (низ–верх) происходила, в результате взаимодействия организмов с гравитационным полем. Одни, становясь тяжелее воды, опускались на дно (скажем, морские звезды). Другие, наоборот, становясь легче, всплывали к поверхности (медузы). При этом и те, и другие оборачивались “лицом”, главными рецепторами к жизненно важным потокам информации от среды.
Асимметризация по второй оси (зад–перед) – происходила при взаимодействии с пространственным (мотивационным) полем, когда понадобилось быстрое, целенаправленное движение (спастись от хищника, догнать жертву). В результате этой асимметризации в передней части тела оказались главные рецепторы и мозг. Следовательно, адаптивность асимметризаций по этим осям понятна. Чего нельзя сказать про латеральную ось. Что дает латеральная асимметризация, и анизотропия какого фактора среды диктует ее развитие? Поскольку теория приписывает одинаковый эволюционный ранг всем 4-м типам симметрии, и в качестве первых двух полей анизотропии выступают фундаментальные физические факторы среды (гравитация и пространство), можно было думать, что поле анизотропии и по третьей оси также должно быть фундаментальным. По идее дихронизма таким полем является временнόе, т.к. левое полушарие “авангардное” (как бы, уже в будущем), а правое – “арьергардное” (еще в прошлом).
Следовательно, потоки информации от анизотропной среды, индуцируют в организмах и органах соответствующие градиенты информационного потенциала – асимметрию.
Конечно, дело не только в классификации типов симметрии, а в тех важных выводах, которые можно сделать из такого понимания. 1. Асимметризация не частное, сугубо-человеческое явление, а общебиологический, фундаментальный, эволюционный феномен, присущий всем живым системам. Поэтому все попытки объяснить асимметрию мозга, уникально-человеческими свойствами (речь, праворукость, самосознание), а так же, как патологию, аномалию развития или родовую травму – нужно отбросить. 2. Эволюция живых систем сопровождалась последовательной асимметризацией по всем трем координатным осям. 3. Латеральная асимметрия – последняя, эволюционно самая молодая. 4. Появляется новый тип триаксиально асимметричных систем (органов, организмов, популяций), внутри которых асимметризация продолжается (цветков и листьев, более асимметричный мозг у мужского пола!). 5. Асимметризация имеет адаптивное значение: она возникает в ответ на анизотропизацию среды. 6. Четыре типа симметрии и три перехода между ними составляют конститутивно-факультативный эволюционный ряд (когда наличие предшествующих непременное условие появления последующих), т.е. у ТА организмов сохраняются две предыдущие асимметризации. 7. Латеральная асимметризация, появляется на фоне предшествующих (спина–брюхо от медузы, нос–хвост от опоссума), поэтому должна распространяться спереди назад. 8. В асимметриях мозга так же можно считать, что из 3-х его асимметрий, самая древняя – кора–подкорка (от медузы), потом – лоб–затылок (от опоссума) и самая молодая – латеральная (наша). 9. По той же причине внутрипарнаяасимметрия парных органов ТА форм возникает на фоне, уже существующей у БС форм, межпарнойасимметрии (как бы реликтовой). 10. Латеральная асимметризация, как и половая дифференциация, идет “снизу–вверх” (функция®орган®организм®популяция). 11. Для её понимания достаточно рассмотреть только два последних типа симметрии и переход между ними, т.е. БС®ТА.
Кроме того, описанная картина глобально-перманентной панасимметризации, позволяет предположить. 1. Эта тенденция действует не только между типами, но и внутри них, не только на уровне организма, но и органов (непарных, парных), а также популяций, т.е. эволюция любых структур идет, как правило, от симметричных к асимметричным; 2. Парные органы БС-форм возникли позже непарных; при этом, и те и другие вначале были симметричны, а асимметризация происходила потом; 3. У ТА-форм, в процессе прогрессивной эволюции, происходит дальнейшая асимметризация, которая, в принципе, может привести к превращению парных органов, в разные органы.
В этом разделе мы рассмотрели проморфологию целых организмов: от одноклеточных до человека. Дальше речь пойдет о латеральной асимметрии человеке на уровне отдельных функций (элементарная единица асимметрии), парных органов (как и в ЭТП, вскрываются механизмы), организма (основной уровень) и популяции (элементарная единица эволюции).
5. Асимметрия функций и парных органов.
Естьорганы непарные и парные. Асимметрия первых может быть только морфологической: по их локализации, форме или ориентации. У вторых возможна еще асимметрия размеров и функциональная асимметрия – по эффективности выполнения функций, т.е. она может быть богаче и фигурировать, как полный морфо-функциональный латеральный диморфизм(аналог полового диморфизма)– отклонение от внутрипарной,зеркальной симметрии (одинаковости).
Среди парных органов человека особое место занимают полушария мозга, как управляющие, а из управляемых – руки, как самые многофункциональные из них, асимметричные и изученные. Поскольку, по сравнению с рукостью, вклад других парных органов в асимметрию целого организма мал, то упростив несколько проблему, рассмотрим только внутрипарные асимметрии полушарий, рук, и их межпарные отношения.
Внутрипарнаяасимметрия органов проявляется в виде доминантности одного и субординатности другого органа по данной функции. D-орган лучше выполняет функцию, поэтому для организма предпочтительный. Конечности умелые, сильнее, ловче, чуть больше размеры костей, ногтей и мышц; рецепторы чувствительнее.
5.1. Асимметрию создают не разные функции, а разные фазы их эволюции.
В настоящее время ФАМ твердо установленное явление. Новая теория рассматривает её в эволюционном плане.
Для чего возникла асимметрия? Почему одни функции управляются лучше левым полушарием, другие – правым? Что дает эта разница? Какова эволюционная логика, смысл этой асимметризации? Конечно, это специализация. Но, по какому принципу? Поиском этого принципа занимались многие, было предложено более двадцати гипотез. Безуспешно. Пришли даже к выводу, что его вообще не существует, или будет открыт в далеком будущем! Потому, что ищут: “какие функции в каком полушарии?” А факты говорят о том, что многие функций и там, и там (см. 2.1.). Но качество управления функций разное: почти у всех людей лучше левополушарное, примерно у 1% – наоборот, или одинаково.
В процессе адаптивной эволюции, при изменении среды, возникают новые проблемы и новые функции, для их решения. В дальнейшем среда может измениться так, что функция окажется ненужной или даже вредной, тогда она элиминируется отбором. Стало быть, любая функция имеет свою “филогенетическую жизнь”: где-то “рождается”, развивается, проходит становление, усовершенствование, где-то работает и где-то “умирает”.
Все существующие теории исходят из синхронной модели эволюции, поэтому трактуют асимметрию (специализацию) полушарий, как результат локализации в них центров разных функций. По новой же концепции, асимметрия – следствие асинхронной эволюции, поэтому полушария предназначены для разных фаз эволюции любых функции.
5.2. Транслокации и “возрастной принцип” специализации полушарий.
В отличие от моделей, “оседлых” (приписанных по полушариям) центров функций, новая концепция исходит из того, что центры функций “кочуют”, они появляется в левом полушарии (как гены в мужском геноме), проходят там становление, подвергаются апробации и отбору, после чего, оставаясь конечно в левом, попадают и в правое (женский геном). Работают, стареют и, если со временем становятся ненужными, утрачиваются, сначала левым полушарием, потом правым (рис 5).
Стало быть, в филогенезе любой функции можно выделить, три фазыпо эволюционному “возрасту” этой функции и, связанные с ними, три разные формыих локализации по полушариям. 1. Молодые (новые) функции только в левом полушарии (онтогенетическом, социокультурном. 2. Зрелыефункции, прошедшие апробацию, основная масса, актуальных, рабочих функций одновременно, и в левом(продвинутые “версии”) и в правом(ранние). 3. Старые функции (утраченные уже ЛП) – только в правом(биологическом). Это – “возрастной принцип” локализации центров по полушариям, как раз тот, который считали, что не существует, или будет открыт в далеком будущем (Геодакян, 1993).
Центры, которые только в ЛП соответствуют сугубо мужским генам (Y-хромосомным). Это – “карантинная служба” генома. Центры в обоих полушариях соответствуют локализации генов в аутосомах, где они проводят основную часть своего филогенеза, находясь в геномах и того и другого пола. Если же в дальнейшем, какие-то признаки становятся ненужными, их гены покидают аутосомы и становятся сугубо женскими, в виде “архивных” генов, кодирующих атавистические признаки. Их эволюционный смысл – в случае возврата старых условий вернуть эти признаки, не прибегая к сложной и долгой процедуре проверки в мужском геноме. Аналогично, при утрате функций, управляющие ими центры элиминируются в ЛП и остаются только в ПП, как “архивные”, и проявляются, как атавистические функции (например, хватательный рефлекс).

На рис. 5 показаны моменты и направления предсказанных транслокацийпри переходах между стадиями симметрии и асимметрии.
5.3. Доминирует не орган, а функция.
Существующие теории оперируют только одной формой асимметрии органов – внутрипарно-зеркальной и не учитывают межпарныхотношений, т.е. реликтовой асимметрии, и, возникающих на уровне двух пар органов нового вида асимметрии – цис-транс. Обычно доминантность приписывают органам. Тогда непонятно, почему у разных функций одного органа бывает разное направление доминирования? Если у органа не одна функция, их векторы доминантности могут иметь разные направленияивеличины, т.е. человек может быть по одной функции (скажем, писать) сильно праворук, по другой (хватать) – слабо леворук, а по третьей.– амбидекстр(симметричен). И так по всем органам.
Значит, элементарной единицей (носителем) асимметрии является не орган, а функция. А асимметрия органа – векторная сумма асимметрий разных его функций. Поэтому, доминирует не правый или левый орган, а функция справа или слева, т.е. доминантность – атрибут функций, а не органа!
Асимметрия организма такая же сумма асимметрий органов. Чтобы понять адаптивность, нужна еще популяционная асимметрия, т.е. асимметриячисленности, дисперсий и латерального диморфизма субпопуляций левшей и правшей. Тогда, полушарность, рукость, правша, левша – некие усредненные характеристики, определяемые суммарно по числу, направлениюистепени доминантности их функций. А из 4 уровней асимметрии: функция, орган, организм и популяция, в “чистом” виде доминантность только у функций, остальные – мозаики.
5.4. Что такое доминантность и субординатность?
Доминантность– это форма проявления внутрипарной зеркальной асимметрии, вектор, показывающий её направление и величину. Существующие теории не ставят вопроса: почему органы пары разные, и никак не трактуют смысл доминантности. Новая концепция трактует доминантную и субординатную формы функций, как разные фазы их эволюции по качеству выполнения.
Глубокий изоморфизм явлений пола и асимметрии позволяет думать, что понятия доминантности–рецессивности в генетике, описывающие проявление генов в фенотипе, могут помочь понять близкие явления доминантности–субординатности функций в нейробиологии. Впрочем, и в генетике пока нет четкого понимания природы доминантности.
В конце 30-х годов А.С. Серебровским и, независимо от него, Д.Д. Ромашовым была высказана мысль о доминировании более поздних признаков, приобретенных в процессе эволюции, по сравнению с ранними (Малиновский, 1970). Такая связь, а также связи между явлениями фило-онтогенеза, мутирования и доминирования были вскрыты в экспериментах по наследованию окраски цветков. Трактуя рецессивные мутации в гомозиготном состоянии (аа), как остановки развития признака на той или иной стадии его формирования, тем самым, выявляющие результаты действия предшествующих генов, а более редкие, доминантные мутации – как добавления к существующей цепи развития нового звена, было показано, что в онтогенезе действительно доминируют более, продвинутые в филогенезе признаки (Ратькин и др, 1988; Геодакян, 1984a).
Это можно понять и из более общих соображений. В процессе прогрессивной эволюции объем генетической информации неуклонно увеличивался. Если при оплодотворении, образовании гибридной молекулы ДНК, от отца и матери участвуют молекулы разной длины, то для новой молекулы возможны два варианта: либо она будет иметь длину более короткой цепочки (т.е. непарные звенья не достраиваются), либо – более длинной (достраиваются). Очевидно, что в первом случае полезные эволюционные приобретения не имеют шансов выжить, тогда как второй, наоборот, способствует прогрессу, т.к. доминирует эволюционно более продвинутая форма. Однако этот вывод справедлив, только для прогрессивной эволюции, когда появляется новый признак (более длинная молекула), а для регрессивной, при утрате признака, длиннее будет молекула старой формы. Это значит, что всегдадолжнадоминироватьформа, обладающая большей информацией. Ведь отсутствующая информация не может доминировать над присутствующей. Следовательно, доминантность определяется, прежде всего, количеством информации: обладающее большей информацией по данной функции – мажорное полушарие доминирует над, обладающим меньшей информацией, минорным.
– Конец работы –
Эта тема принадлежит разделу:
Эволюционные теории асимметризации организмов, мозга и тела
На сайте allrefs.net читайте: "Эволюционные теории асимметризации организмов, мозга и тела"
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Эволюционный ряд основных типов морфологической симметрии организмов.
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.03 сек.
Новости и инфо для студентов