рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Философия
- /
- Ископаемые человекообразные обезьяны
Реферат Курсовая Конспект
Ископаемые человекообразные обезьяны
Ископаемые человекообразные обезьяны - раздел Философия, АНТРОПОЛОГИЯ Ископаемые Человекообразное Обезьяны Известны Только В Старом Свете, Начиная ...
Ископаемые человекообразное обезьяны известны только в Старом Свете, начиная с нижнего олигоцена.
Древнейшим и самым примитивным их представителем является парапитек (Parapithecus ftaasi) из нижнего олигоцена Файюма (Египет). От него известна лишь одна нижняя челюсть длиною 3,6 см с сильно расходящимися половинками (угол=33°), что свидетельствовало о коротком лицевом отделе и ширине мозговой коробки; эти особенности сближают парапитека с долгопятами. Парапитек — исходная форма для позднее развившихся антропоидов, в том числе и человека. Ближайшим потоком его является проплиопитек.
В нижнем олигоцене Египта была найдена нижняя челюсть и другой обезьяны. В правой половине челюсти сохранились клык, два предкоренных и три коренных зуба. Эта обезьяна, получившая название проплиопитека (Propliopimecus), близка к гиббонам. Их сближает, в частности, расположение добавочного бугорка — гипоконулида, который находится точно посередине заднего края та-лонида. Имеются, однако, и отличия от современных гиббонов: так, челюсть менее крупная, чем у гиббона, первые премоляры развиты слабее. Клыки проплиопитека значительно меньше, чем у гиббона, но крупнее, чем у парапитека.
Примерно с 1962 г. Элвин Саймоне производил раскопки в Файюме (Египет), где нашел новые остатки проплиопитека, а кроме того, неизвестную форму примата, получившую название египтопитека (Aegyptopitnecus zeuxis). Египтопитек представлен нижней челюстью и зубами верхней челюсти. Египтопитек — это очень примитивный дриопитек. Предположительно к египтопитеку могут быть отнесены также крупная метатарзальная кость, несколько фаланг и каудальных позвонков. Интерес к египтопитеку возрос в связи с предположением, что он является предком рамапитека и обладает чертами гоминид.
Миоцен — период развития высших обезьян. Многочисленные остатки разных видов антропоморфных извест-
* 167


 ны из миоценовых отложений Европы, Индии, Экватори
ны из миоценовых отложений Европы, Индии, Экватори
альной Африки. •
Плиопитек (Pliopithecus Gervais) — самая распространенная обезьяна среднего миоцена Европы. Его остатки, представленные фрагментами нижних и верхних челюстей, найдены на территории Польши, Чехословакии, Швейцарии, Австрии, Германии, Франции. Плиопитек близок к гиббонам. Отличия его от современных гиббонов сводятся к несколько большей примитивности. Для плиопи-тека характерна менее удлиненная форма коронки предко-ренных и коренных зубов, более длинный симфиз нижней челюсти, более сильная дивергенция половинок нижней челюсти (25°; у гиббоновых —16—21°), наличие одного бугорка на нижнем переднем предкоренном вместо двух, имеющихся у гиббона, меньшие размеры клыков, наличие цингулюма, которого нет у гиббона.
До начала второй мировой войны в течение ряда лет велись раскопки под Нейдорфом (Чехословакия), которые привели к замечательному открытию костных остатков антропоморфной обезьяны, получившей название эпиплио-питека (Epipliopithecus vindobonensis). Были обнаружены многочисленные кости черепа, зубы, а также почти полные остатки посткраниального скелета, в том числе некоторые шейные, грудные и поясничные позвонки, плечевые кости, лучевая и локтевая, кости таза, бедренные кости, кости голени, стоп, «истей. Кости принадлежат трем особям и датируются средним миоценом. Наибольшее сходство эта обезьяна обнаружила с плиопитеком (P. antiguns). Цапфе (1960) отмечает следующие характерные черты в черепе этой находки: очень малая ширина нижнего отдела морды; относительно большая межглазничная ширина, несколько напоминающая полуобезьян, широконосых обезьян и антропоморфных; положение скуловой кости сходно с низшими обезьянами и гиббонами; удлиненность грушевидного отверстия и его суженность в нижней части напоминает низ-
ших обезьян и гиббонов. Отсутствие сагиттального гребня или очень слабое его развитие имеет аналогию с широконосыми и гиббонами; выраженность затылочного рельефа, напоминающая самцов низших узконосых и самцов некоторых широконосых; низкий верхнелицевой и высокий фронтобиорбйтальный указатель.
Цапфе (1960) сделал примерный подсчет степени сходства по форме черепа эшшлиопитека с другими приматами: 1 признак сходства с понгидами, 8 — с гиббонами, 7 — с низшими узконосыми, 8 — с широконосыми, 2— с полуобезьянами.
В позвоночнике эпиплиопитека 7 шейных, 12 или 13 грудных, 6 или 7 поясничных, 3 крестцовых и 3 хвостовых позвонка. Форма таза вполне сходна с той, что имеется у низших обезьян. Такое же смещение типических признаков, напоминающих то низших узконосых, то широконосых обезьян, то полуобезьян, наблюдается на кистях, стопах и длинных костях. Наиболее поразительной особенностью у эшшлиопитека, пожалуй, следует считать сочетание вполне выраженного сходства по строению зубов с антропоморфными обезьянами и резко выраженного сходства по пропорциям тела с низшими узконосыми.
По способу локомоции эпиплиопитек, без всякого сомнения, не был чисто древесным существом. У него отсутствуют признаки специализации как к брахиации, так и к наземному беганию на четырех конечностях. В этом отношении он может считаться «генерализованной» формой. Судя по остаткам сопровождавшей фауны, он жил в условиях сухого лесного биотопа.
Для проблемы антропогенеза дриопитеки Европы интересны тем, что представляют собой древнейшие остатки антропоморфных, весьма близких к африканским высшим обезьянам. Еще более велико их значение в том, что по некоторым особенностям отдельные формы более сходны с человеком, чем любая из ныне живущих антропоморфных обезьян.
'*■ 169

 В среднем миоцене в Западной Европе описано 2 вида дриопитека: дриопитек фонтанова (Dryopithecus fontani Lartet) и дриопитек дарвинов (D. darvini Abel). Остатки дриопитека фонтанова известны не только из среднего миоцена, но и из нижнего плиоцена. От дриопитека фонтанова сохранились четыре нижних челюсти, нижние коренные зубы, один верхний коренной и плечевая кость. Это была антропоморфная обезьяна ростом примерно с шимпанзе. Зубы, в особенности коренные, очень сходны с зубами гориллы; клыки крупные. Основными отличиями от зубов гориллы является меньшая высота бугорков, менее четко выраженный основной рельеф жевательной поверхности, относительно меньшие размеры резцов. Плечевая кость дриопитека фонтанова тонкая, лишена сильного мышечного рельефа, что отличает ее от соответствующей кости шимпанзе и гориллы. Хотя на плечевой кости не сохранились эпифизы, тщательный анализ привел некоторых палеонтологов к выводу, что она была сходной с плечевой костью карликового шимпанзе и резко отличалась от плеча как древесных, так и наземных церкопите-ковых.
В среднем миоцене в Западной Европе описано 2 вида дриопитека: дриопитек фонтанова (Dryopithecus fontani Lartet) и дриопитек дарвинов (D. darvini Abel). Остатки дриопитека фонтанова известны не только из среднего миоцена, но и из нижнего плиоцена. От дриопитека фонтанова сохранились четыре нижних челюсти, нижние коренные зубы, один верхний коренной и плечевая кость. Это была антропоморфная обезьяна ростом примерно с шимпанзе. Зубы, в особенности коренные, очень сходны с зубами гориллы; клыки крупные. Основными отличиями от зубов гориллы является меньшая высота бугорков, менее четко выраженный основной рельеф жевательной поверхности, относительно меньшие размеры резцов. Плечевая кость дриопитека фонтанова тонкая, лишена сильного мышечного рельефа, что отличает ее от соответствующей кости шимпанзе и гориллы. Хотя на плечевой кости не сохранились эпифизы, тщательный анализ привел некоторых палеонтологов к выводу, что она была сходной с плечевой костью карликового шимпанзе и резко отличалась от плеча как древесных, так и наземных церкопите-ковых.
Дриопитек дарвинов представлен несколькими зубами, из которых наиболее интересен нижний последний корзиной, весьма сходный с человеческим. Его отличает от соответствующего зуба человека наличие резко выра-жекного цингулюма.
Дриопитек был также найден в нижних и верхних слоях среднего миоцена (Нижний Сивалик) и в верхнем миоцене. К нижнесиваликским относятся виды: D. punjabicus, D. chinjiensis, D. pilgrimi, D. sivalensis. К более поздним формам относятся: D. gigantens, D. cantleyi, D. fricae. Bee эти виды, выделенные на основании фрагментов челюстей; с зубами или отдельных зубов, отличаются друг от друга по положению бугорков или по общей форме и ве-лкпгане зубов.
Одни из этих видов более сходны с гориллой, другие — с шимпанзе. D. chinjiensis, бесспорно, сходен с гориллой. D. pinjabicus напоминает и гориллу й шимпанзе. D. gigantens близок к шимпанзе, однако значительно превосходит его но своей величине. D. pilgrimi ближе всего к дриопитеку фонтанову Западной Европы, но несколько прогрессивнее.
Весьма интересная находка человекообразной обезьяны, датируемая верхним миоценом, была сделана на территории Грузии. В1939 г. в Удабно (Кахетия) в верхне-сарматских отложениях был обнаружен фрагмент верхней челюсти с двумя зубами — вторым предкоренным и первым коренным. В 1945 г. эта находка была детально описана Н.О. Бургак-Абрамовичем и Е.Г. Габашвили под названием удабнопитека (Udabnopimecus garedaensis).
Первый коренной зуб удабнопитека имеет много черт сходства с человеческим зубом в размерах, общем плане и рисунке жевательной поверхности. Наиболее существенные отличия от человека сказываются в следующих особенностях: у удабнопитека: латеральныеТ>угры и гребни тритона и его ямки значительно более выражены; на латеральной стенке коронки имеется рудиментарный цингу-люм; предкоренной имеет.менее округлые контуры, с менее расширенной латеральной частью коронки по сравнению с медиальной; разделенность щечного и язычного бугорков менее резкая и глубокая; на медиальной стенке имеется цингулюм, отсутствующий у человека. Зуб имеет три корня в отличие от человеческого, обладающего одним корнем. Большое значение этой находки состоит в том, что ранее ни одной человекообразной обезьяны на территории Восточной Европы и Западной Азии не было известно. Удабнопитек заполняет, таким образом, пробел в распространении высших приматов в миоцене и соединяет западноевропейскую и сиваликскую фауну.
В 1955 г. в Малой Азии недалеко от г. Анкары в ело-
v 171




 ях верхнего миоцена была обнаружена нижняя челюсть крупной, по-видимому, высшей обезьяны с зачаточным подбородочным выступом, получившей название анкара-питек.
ях верхнего миоцена была обнаружена нижняя челюсть крупной, по-видимому, высшей обезьяны с зачаточным подбородочным выступом, получившей название анкара-питек.
В нижних слоях среднего миоцена в Сиваликских холмах были найдены представители следующих высших обезьян: палеосимия (Palaeosimia Pilgrim), сивапитек (Siva-pithecus Pilgrim), дриопитек (Dryopithecus Lartet), сугрива-питек (Sugrivapithecus Lewis), брамапитек (Bramapithecus Lewis), рамапитек (Ramapithecus Lewis).
Палеосимия представлена одним верхним коренным
зубом. Общий вид и мелкая морщинистость жевательной
поверхности этого зуба более всего сближаются с зубом
оранга. Таким образом, некоторые характерные черты зуб
ной системы оранга уже отчетливо проявляются в ниж
них слоях среднего миоцена. ,
Сивапитек, включающий несколько видов, представлен главным образом хорошо сохранившимися и достаточно полными фрагментами нижних и верхних челюстей. Положение сивапитека среди высших обезьян не вполне ясно. Для него характерно сочетание признаков, сближающих его отчасти с орангом, отчасти с дриопитеком.
Сугривапитек представлен левой половиной нижней челюсти с ячейкой и корнями М3, вполне сохранными М2 и М, и вторым Р, сохранились также корни первого Р, клыка, второго резца и ячейки первого резца. Эта форма интересна многими чертами сходства с человеком. К ним относятся: зачатки подбородочного выступа, сходные с человеческим, строение двухбугоркового второго предкорен-ного, малый размер клыков и резцов, отсутствие диас-тем. Однако у сугривапитека имеются черты, которые резко отличают его от человека: предкоренные зубы, в особенности первый, имеют обезьяноподобное строение, первый коренной гораздо меньше по размеру, чем второй, причем очень узкие по форме.
Брамапитек, известный по фрагментам левой половины нижней челюсти с М3, М2, ячейкой и корнями Mj и частью ячейки и корня второго Р, представляет сочетание обезьяньих (преобладающих) и некоторых человеческих признаков. К обезьяньим чертам относятся крайние вы-ступание метаконида на М2 и на М3, довольно заметное развитие сингулюма на М3. Однако по ширине коронки коренных зубов брамапитек далеко превосходит всех ископаемых обезьян.
Рамапитек — одна из наиболее важных находок из числа миоценовых высших обезьян — представлен двумя видами: Ramapithecus hariensis Lewis, датируемый миоценом, и Ramapithecus brevirostris, относящийся к плиоцену.
Миоценовый рамапитек представлен несколькими находками. Впервые он был описан по фрагменту правой половины верхней челюсти с М, и М2. При общем сходстве с человеческими эти зубы обладают примитивными | чертами — остатками цингулюма на М, и малой величиной М, по сравнению с М2.
Позднее в среднемиоценовых слоях были обнаруже
ны новые остатки рамапитека — передняя часть нижней
челюсти с правой половиной тела и с зубами М^, М,, Рх,
Р2; кроме того, сохранились ячейки от С, Lj, I,. Зубы на
этой челюсти имеют черты сходства с человеческими, од
нако совершенно отличен от человеческого вытянутый в
длину Pj, а ячейка клыка гораздо больше ячейки латераль-
ногорезца. ^
В раннем миоцене в Кении (Восточная Африка) вместе с остатками лемуроидов, динотерия, трех родов крео-донтных, амфициона и других млекопитающих были обнаружены остатки трех родов высших обезьян: ксенопи-тека (Xenopimecus Hopwood), лимнопитека (Limnopithecus Hopwood), проконсула (Proconsul Hopwood).
Ксенопитек — род, положение которого среди других ископаемых и современных высших приматов еще не выяснено.

 Лимнопитек, от которого сохранились остатки зубов, челюстей и некоторых других частей скелета, представлен двумя видами, из которых один некоторые исследоватег ли сближают с гиббоном, а другой — с симфалангом. Подробное изучение материалов по лимнопитекам, сделанное Д. Ференбах, привело к существенно другим выводам. Оба вида (L. legetet и L. macinesi) не имеют никакого специфического сходства с гиббонами. L. legetet очень похож на Pongidae, более всего на шимпанзе. L. macinesi многими чертами отличается от L. legetet. Строение конечностей сближает егас шимпанзе и колобусом; нижняя челюсть и зубы — с шимпанзе, гориллой, орангом, колобусом и макаком. Важно заключение Д. Ференбах, что по своей способности к брахиации L. macinesi приближается к шимпанзе.
Лимнопитек, от которого сохранились остатки зубов, челюстей и некоторых других частей скелета, представлен двумя видами, из которых один некоторые исследоватег ли сближают с гиббоном, а другой — с симфалангом. Подробное изучение материалов по лимнопитекам, сделанное Д. Ференбах, привело к существенно другим выводам. Оба вида (L. legetet и L. macinesi) не имеют никакого специфического сходства с гиббонами. L. legetet очень похож на Pongidae, более всего на шимпанзе. L. macinesi многими чертами отличается от L. legetet. Строение конечностей сближает егас шимпанзе и колобусом; нижняя челюсть и зубы — с шимпанзе, гориллой, орангом, колобусом и макаком. Важно заключение Д. Ференбах, что по своей способности к брахиации L. macinesi приближается к шимпанзе.
Проконсул, представленный обломком левой верхней челюсти с рядом зубов от С до Щ, обломком нижней челюсти, несколькими изолированными зубами и рядом костей скелета, считается предковой формой шимпанзе, с которым обнаруживает много сходных черт. Однако он отличается от шимпанзе некоторыми признаками, в общем свидетельствующими о большей примитивности проконсула; так, первый предкоренной у него очень напоминает клык, цингулюм на коренных чрезвычайно сильно развит, морщинистость жевательных Поверхностей мала, весь ряд зубов прорезывался примерно в одно и то же время, между теьЛ как у шимпанзе сроки прорезывания Щ и М, разделены очень большим промежутком времени.
Находка почти полного черепа проконсула в миоценовых слоях, близ озера Виктория, обнаруживает следующее своеобразное сочетание признаков: при резко скошенной кзади профильной линии нижней челюсти, при сильном выступают морды кости черепа очень тонки, а надглазничный валик полностью отсутствует. Род проконсулов включал виды, сильно варьирующие по размерам тела примерно от величины гиббона до гориллы.
В1961 г. Л. Лики опубликовал заметку о новом роде
кениапйтеке (Kenyapithecus Wiekeri), который он сначала
датировал плиоценом, а затем верхним миоценом, при
чем подчеркнул сходство с рамапитеком. Кениапитек был
обнаружен в Кении и представлен фрагментом верхней
челюсти. Многие специалисты сочли возможным сбли
жение этих форм и допустили их принадлежность к очень
ранним гоминидам. Абсолютная древность кениапитека
оказалась равна 14 млн лет. ,
Самой замечательной находкой раннего плейстоцена (Южный Китай) являются три коренных зуба огромного размера, принадлежащие какой-то антропоморфной обезьяне, получившей название гигантопитека (Gigantopithecus blacki). В 1934—1939 гг. в аптекарских лавках Гонконга были обнаружены три зуба гигантопитека — нижний правый Mj, верхний Mj, нижний левый Mj, причем у первых двух отсутствовали корни, а у последнего сохранился только задний корень.
В этих зубах поразительна прежде всего их величина. Так, длина коронки левого М3 у гигантопитека равна 22,3 мм, а у современного человека не превосходит 11 мм, ширина ее соответственно 17,0 и 11,0 мм, показатель мощности (длина, умноженная на ширину) составляет у гигантопитека 355 мм2, а у современного человека — 115 мм2. По объему коренной зуб гигантопитека превосходит человеческий приблизительно в 6раз.
Особенности зубов, сближающие гигантопитека с человеком, — отсутствие диастем, двухбугровый тип пред-коренных, не имитирующий форму клыка, малая высота клыка, лишь немного превышающая высоту резцов. Однако клыки очень мощные и толстые. Кроме того, на коренных, предкоренных и клыках имеется поясок (цингу-люм). Наиболее резкой отличительной чертой этих челюстей можно назвать их огромный размер в сочетании с типичным для обезьян строением. Профиль переднего от-
% 175


 дела скошен назад, никакого следа подбородочного выступа не имеется.
дела скошен назад, никакого следа подбородочного выступа не имеется.
Начиная с 1924 г. в Южной Африке были обнаружены многочисленные остатки ископаемых антропоморфных, изучение которых привело к выводам исключительно важного значения для проблемы антропогенеза. Эти костные остатки принадлежат высшим приматам, объединенным в одно подсемейство австралопитековых (Austra-lopithecinae), куда входят три рода с пятью видами (рис. 37). Хронологическая последовательность этих находок такова:
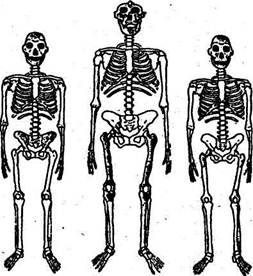
|

|
| Australopithecus afarensis |
| Australopithecus Australopithecus robustus Нолю africanus and/4, boisei habilis |
| Рис. 37. Реконструкция скелетов австралопитековых |
—Australopithecus africanus Dart (почти полный че
реп детеныша) обнаружен в 1924 г. в восточной
части пустыни Калахари.
—Ptesianthropus transvaalensis Broom (около 12 более или
менее полных черепов, свыше 150 отдельных зубов,
дистальный конец бедренной кости, часть фаланги
пальца руки, головчатая кость запястья, бедро, боль
шая берцовая, ребра, позвонки, лопатка, фрагменты
таза). Остатки обнаружены в 1936—1947 гг. в извест-
, коюй пещере к югу от г. Претории.
— Parantfiropus robustus Broom (фрагменты черепа, ди-
стального конца плечевой кости, проксимального
конца локтевой кости, фаланги кисти) найден в
1938—1939 гг. также близ г. Претории.
, — Australopithecus promentheus Dart (фрагменты черепа, крупные обломки таза, зубы) добыт в 1947— 1949 гг. в каменоломнях Макапанстада (центральный Трансвааль).
— Paranmropus crassidens Broom (фрагменты нижних
и верхних челюстей, зубы) открыт в 1948—1949 гг.
близ Стеркфонтейна в пещере Сворткранс.
Вопрос о древности южноафриканских австралопитеков еще окончательно не разрешен. Сначала возраст этих находок определяли как ранне- и даже среднеплейстоце-новый. Однако впоследствии были приведены доказательства их большей древности.
В целом все названные животные, несомненно, близкие родичи африканских высших обезьян, однако они обнаруживают значительно большее сходство с человеком, и по мнению большинства ученых, входят в семейство гоминид. Это сходство сказывается в приспособленности их тела к выпрямленному двуногому хождению, в строении зубов, в некоторых особенностях черепа. Двуногое хождение приписывается австралопитековым на основании следующих фактов:
1. Строение таза австралопитека прометеева, австралопитека трансваальского и парантропа крупнозубого весьма сходно с человеческим и отличает его от современных шимпанзе и гориллы. Подвздошная кость


 австралопитека и по абсолютным размерам, и по форме поразительно похожа на человеческую. Тазовые кости плезиантропа отличались от человеческих главным образом меньшими размерами и большей упло щенностью седалищных бугров.
австралопитека и по абсолютным размерам, и по форме поразительно похожа на человеческую. Тазовые кости плезиантропа отличались от человеческих главным образом меньшими размерами и большей упло щенностью седалищных бугров.
2. Характер поверхности затылочной кости свидетель
ствует о том, что у австралопитека прометеева чешуя
была более свободна от шейной мускулатуры, чем у
шимпанзе; по-видимому, его голова довольно хоро
шо балансировала на более или менее вертикально
направленном шейном отделе позвоночника.
3. У Paranmropus pobustus сочленовные отростки заты
лочной кости занимали более переднее положение,
чем у шимпанзе; то же, по-видимому, можно утвер
ждать и о положении его большого затылочного от
верстия.
4. Фауна, найденная вместе с австралопитековыми, сви
детельствует о том, что средой их обитания был не
лес, а открытые пространства. Наиболее частыми
«спутниками» австралопитековых были маленького
размера, ныне вымершие павианы. Остатки долгоно
са, очень сходного с кафрским долгоносом, которого
раньше зоологи относили к тушканчикам, говорят о
полупустынном ландшафте. Ботанические и геологи
ческие данные также свидетельствуют об отсутствии
лесов в центральной и западной областях Южной
Африки как в плейстоцене, так и в конце плиоцена.
5. При исследовании 58 черепов павианов, найденных
вместе с австралопитековыми, оказалось, что 80%
черепов несли на себе явные следы сильных раскалы
вающих ударов и окаменели уже после того, как эти
удары были нанесены. Многие следы ударов на чере
пах павианов имели вид двойных вмятин, причем рас
стояния между ними оказались в точности соответ-
ствующими ширине между мыщелками длинных костей копытных, найденных по соседству. По-видимому, австралопитеки охотились на павианов с помощью камней и плечевых костей копытных. Таким образом, австралопитеки использовали в качестве ударных х>рудий плечевые, бедренные и болыпебер-цовые бычьи кости; в качестве режущих пластин — нижние челюсти, безыменные кости и лопатки; в качестве колющих орудий — рога, костные стержни рогов, продольно расколотые кости; в качестве скребков кости неба.
Двуногая походка африканских обезьян отличалась от человеческой и была, без сомнения, менее совершенной. Этот вывод можно обосновать главным образом на строении таранной кости парантропа. Хотя по размеру она близка к таранной кости женщины маленького роста, однако отлична от нее по форме: суставная поверхность для сочленения с ладьевидной костью гораздо шире, более горизонтально расположена и заходит дальше внутрь, чем у человека; кроме того, шейка сильно отклонена внутрь, что свидетельствует о значительном отведении первого пальца. Имеется ряд отличий от человека и в строении бедренной кости.
Огромное сходство с человеком обнаружилось в строении постоянных и молочных зубов австралопитековых. Более всего приближаются к человеческим зубы парантропа и австралопитека. У парантропа, как и у человека, зубной ряд верхней и нижней челюстей не имеет диастем, клыки маленькие и не выдаются из уровня соседних зубов, предкоренные имеют по 2 округлых бугорка — язычный и щечный; по характеру поверхности коренные зубы сходны с человеческими, в особенности по плоской округленности жевательных бугорков.
Череп парантропа сочетает в себе признаки высшей обезьяны с признаками, напоминающими человека. Про-



 гаащм у него выражен менее, чем у шимпанзе и гориллы. Хотя подбородочный выступ у парантропа полностью отсутствует, однако профильная линия нижней челюсти скошена назад не столь резко, как у современных антропоморфных. С другой стороны, в общем лицевой скелет парантропа отличается исключительной мощностью и толщиной.
гаащм у него выражен менее, чем у шимпанзе и гориллы. Хотя подбородочный выступ у парантропа полностью отсутствует, однако профильная линия нижней челюсти скошена назад не столь резко, как у современных антропоморфных. С другой стороны, в общем лицевой скелет парантропа отличается исключительной мощностью и толщиной.
Очень большое значение имеет находка, сделанная 17 июля 1959 г. в Танзании (Восточная Африка), где супруги Лики обнаружили на глубине 6,71 м от верхней границы олдовайского слоя почти полный череп существа, сходного с австралопитековыми, принадлежащий, вероятно, молодому самцу. Вместе с ним были найдены разбитые на части кости животных, остатки пищи, представлявшие своим раздробленным состоянием яркий контраст совершенно нетронутому черепу. Кости принадлежали птицам, земноводным, пресмыкающимся, грызунам, поросятам, антилопам. Особый интерес представляет то обстоятельство, что в соседстве с черепом находились чрезвычайно примитивные каменные орудия так называемого олдовайского типа, сделанные из гальки.
Череп по некоторым признакам может быть включен в род Paranthropus (сагиттальный гребень, малые размеры клыков и резцов, относительно прямая линия фронтальной части ряда этих зубов, уплощенность лба). Однако по другим признакам он больше похож на род Australopithecus (высокий свод черепа, глубокое нёбо, меньшие размеры М3 по сравнению с Ъ/L). Но у нового черепа имеется много особенностей, которые резко отличают его от всех других австралопитековых, что дало основание Лики выделить его в особый род — зинджантроп.
Наиболее своеобразная особенность черепа зинджан-тропа— необычайное развитие скулового отдела височной кости. Над сильно пневматизированным сосцевидным отростком (сходным по размеру с человеческим) имеется огромный валик. Назион выступает кпереди от глабеллы, носовые косточки суживаются книзу, весьмазначительна
по размеру площадь прикрепления мьшщ на нижнем крае скуловой дуги, высокий черепной свод в заднем отделе. В том же древнейшем слое Олдовайского разреза начиная с 1959 г. были сделаны некоторые ископаемые находки, которые явно отличались от зинджантропа и были значительно более сходны с человеком. В1964 г. Л. Лики, Ф. Тобайас и И. Нейпир опубликовали статью, в которой обозначили эти костные остатки, как принадлежащие виду Homo habilis, что значит, «человек умелый». Именно «пре-зинджантропу» названные выше авторы приписали галечную, или олдувайскую культуру (рис. 38).

– Конец работы –
Эта тема принадлежит разделу:
АНТРОПОЛОГИЯ
А Е Хомутов... АНТРОПОЛОГИЯ... Учебное пособие...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Ископаемые человекообразные обезьяны
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.033 сек.
Новости и инфо для студентов