рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Философия
- /
- Лекция 8 Принципы метаболизма и биоэнергетики
Реферат Курсовая Конспект
Лекция 8 Принципы метаболизма и биоэнергетики
Лекция 8 Принципы метаболизма и биоэнергетики - раздел Философия, Лекция 8 ...
Лекция 8
Принципы метаболизма и биоэнергетики
- в форме органических питательных веществ - хемотрофы; - в форме энергии солнечного света – фототрофы. Метаболизм или обмен веществ - совокупность ферментативных химических реакций в организме, которые обеспечивают его…Высокоэнергетические биомолекулы.
Структура АТФ и её роль в энергетическом обмене.
Возникает вопрос, почему именно молекула АТФ является основным поставщиком химической энергии для биохимических процессов, протекающих с потреблением энергии?
Для ответа на этот вопрос рассмотрим структуру АТФ. Молекула АТФ при физиологических значениях pH существует в идее тетрааниона (рис 8.1):
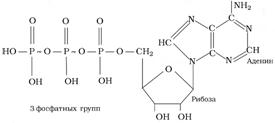
Рис. 8.1. Структура молекулы АТФ
В результате этого возникают два фактора, дестабилизирующих структуру молекулы АТФ:
Во-первых, возникает кластер одноименных отрицательных зарядов, которые отталкиваются друг от друга. Всё это приводит к напряжению молекулы, сильнее всего в области связей -P-O-P-.
Во-вторых, в связи с наличием неподелённых электронных пар у атомов кислорода, расположенного между атомами фосфора, возникает сдвиг электронной плотности и образование частичного положительного заряда на каждом атоме фосфора.
Атомы фосфора конкурируют за неподелённые подвижные электронные пары у атомов кислорода по механизму донорно-акцепторной связи. Это приводит к дополнительному напряжению в структуре молекулы АТФ.
!!! Молекула АТФ должна содержать достаточное количество химической энергии, чтобы компенсировать структурные особенности, выступающие в роли дестабилизирующих факторов.
Молекула АТФ содержит две макроэргические фосфатные связи, при гидролизе которых высвобождается значительное количество свободной энергии:
АТФ + H2O →АДФ + H3PO4 ∆G0' = –31,8 кДж/моль;
АДФ + H2O →АМФ + H3PO4 ∆G0' = –31,8 кДж/моль
Связи, при гидролизе которых изменения энергии составляют более
30 кДж/моль, в биохимии:
- называют макроэргическими связями;
- обозначают знаком ~ (тильда);
- соединения, обладающие такими связями, называют макроэргами.
К соединениям, обладающим макроэргичискими связями, кроме АТФ, относят также УТФ, ГТФ, ЦТФ, ТТФ, креатинфосфат, некоторые тиоэфиры (ацил-КоА) и некоторые другие соединения.
Роль кислорода в метаболизме. Дыхательная цепь. Окислсительное фосфорилирование.
Одной из центральных проблем биоэнергетики является биосинтез АТФ, который в живой природе происходит путём фосфорилирования АДФ:
АДФ + Pi = АТФ
!!!! В связи с этим основной задачей биоэнергетики живых организмов является регенерация АТР из АДФ.
Фосфорилирование АДФ является эндергоническим процессом и требует источника энергии.
В природе существует два таких источника энергии:
- солнечная энергия;
- химическая энергия восстановленных органических соединений.
Способы синтеза АТФ
1) Фотосинтетическое фосфорилирование. Зелёные растения способны трансформировать энергию поглощенного кванта света в энергию химических связей, которая в дальнейшем расходуется на фосфорилирование АДФ.
2) Окислительного фосфорилирование – это трансформация энергии окисления органических соединений в макроэргические связи АТФ в аэробных условиях.
3) Субстратное фосфорилирование. При субстратном фосфорилировании донором активированной фосфорильной группы (~PO3H2), необходимой для синтеза АТФ, являются промежуточные продукты процессов гликолиза и цикла трикарбоновых кислот.
Во всех этих случаях окислительные процессы приводят к образованию макроэргических соединений: 1,3-дифосфоглицерата, и сукцинил-КоА, которые при участии соответствующих ферментов способны фосфорилировать АДФ и образовывать АТФ.
Окислительное фосфорилирование.
Процесс расщепления биополимеров не связан с образованием свободной, т. е. доступной клетке энергии.
!!!! Основной источник энергии в клетке - окисление субстратов кислородом воздуха.
1) В клетках окисление протекает в форме последовательного переноса водорода и электронов от субстрата к кислороду.
2) Кислород играет в этом случае роль восстанавливающегося соединения (окислителя).
3) Окислительные реакции протекают с высвобождением энергии.
Окислительное фосфорилирование или биологическое окисление - процесс синтеза АТФ, при котором электроны и протоны с окисляемого субстрата переносятся с помощью системы окислительно-восстановительных ферментов, локализованных во внутренней мембране митохондрий к кислороду.
Дыхательная цеп или цепь переноса электронов
Любая окислительно-восстановительная реакция включает перенос электронов. При этом: - восстановленный компонент (восстановитель) отдаёт электрон и сам при этом окисляется;Стандартные окислительно-восстановительные потенциалы
1) !!! Все окислительно-восстановительные компоненты реакции отличаются по своему сродству к электронам, т. е. способности принимать электроны. 2) Относительную способность веществ (участников электрон-транспортной цепи)… 3) Значение каждого окислительно-восстановительного потенциала измеряется по отношению к стандартному…Организация дыхательной цепи транспорта электронов клеток эукариот
В клетках эукариот дыхательная цепь локализована во внутренней мембране митохондрий.
Структура митохондрий
Митохондрии - это органеллы размером с бактерию (около 1 х 2 мкм). Обычно в клетке содержится около 2000 митохондрий, общий объем которых составляет до 25% от общего объема клетки (рис. 8.2):
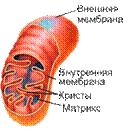
Рис. 8.2. Схема строения митохондрии в разрезе
Митохондрия ограничена двумя мембранами - гладкой внешней искладчатой внутренней, имеющей очень большую поверхность. Складки внутренней мембраны глубоко входят в матрикс митохондрий, образуя поперечный перегородки - кристы. Пространство между внешней и внутренней мембраной носит название межмембранного пространства.
Внутренняя мембрана митохондрий характеризуется необычно высоким содержанием белков (75%). В их число входят транспортные белки-переносчики, ферменты, компоненты дыхательной цепи иАТФ-синтаза.
Компоненты электрон-транспортной цепи
Компонентами электрон-транспортной цепи являются сложные ферменты и простые белки переносчики.
В переносе электронов от субстратов к молекулярному кислороду принимают участие:
1. НАД - зависимые дегидрогеназы;
2. ФАД- зависимые дегидрогеназы;
3. Убихинон (КоQ);
4. Цитохромы b, c, a+a3 ;
Белки, содержащие негемовое железо
НАД-зависимые дегидрогеназы. В качестве кофермента содержат НАД и НАДФ. Пиридиновое кольцо никотинамида способно присоединять электроны и протоны водорода.
ФАД и ФМН-зависимые дегидрогеназы содержат в качестве кофермента фосфорный эфир витамина В2 (ФАД).
Убихинон или коэнзим Q (КоQ) играет роль промежуточного переносчика водородных атомов, т. е. электронов и протонов в митохондриальной мембране, окисляя восстановленную форму флавиновых ферментов и превращается при этом в гидрохинон КоQ∙H2 (рис. 8.3):
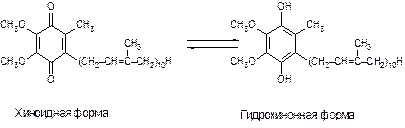
Рис. 8.3. Строение убихинона коэнзима Q (КоQ)
Цитохромы - белки хромопротеиды, способные присоединять электроны, благодаря наличию в своем составе в качестве простетических групп железопорфиринов.
Они принимают электрон от вещества, являющегося немного боле сильным восстановителем, и передают его более сильному окислителю.
Цитохромы разделяют на подгруппы а, b и с. К двум последним группам относят цитохромы, содержащие обычный гем (рис 8.4)
Цитохромы располагаются в митохондриальной цепи между убихиноном и кислородом. При этом цитохромы включаются в дыхательную цепь в определенной последовательности:
Цит. b → Цит. c1 → Цит. c → Цит. aa3
Цитохромы b, c и c1 выполняют функцию промежуточных переносчиков электроновю
Комплекс цитохромов a и a1, называемый цитохромоксидазой, является терминальным дыхательным ферментом, непосредственно взаимодействующим с кислородом.

Рис. 8.4 Строение гема, входящего в состав цитохромов b и с
!!! В отличии от гема гемолгобина атом железа в цитохромах может обратимо переходить из двух в трехвалентное состояние это обеспечивает транспорт электронов.
Белки, содержащие негемовое железо. В этих белках атомы железа связаны с атомами серы цистеиновых остатков или прямо с атомами серы S2-, поэтому их часто называют железосерными белками и обозначают как Fe-S (рис. 8.5):
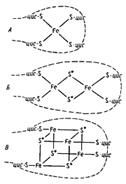
Рис. 8.5. Структура белков, содержащих негемовое железо (Fe-S-белки)
Комплексы окислительного фосфорилирования
Систему окислительного фосфорилирования удается разделить на несколько комплексов, каждый из которых осуществляет одну из главных стадий цепи переноса электронов.
Обычно эти комплексы обозначают как комплексы I, II, III и IV (рис. 8.6):

Рис 8.6. Комплексы дыхательной цепи (электрон-транспортной цепи)
Комплекс I (НАДН: КоQ-оксидоредуктаза) катализирует перенос электронов от НАДН к КоQ:
Два атома водорода эквивалентны двум протонам (Н+) и двум электронам e. НАД+ связывает лишь один протон, а второй остаётся в среде:
SH2 + НАД+ = S + НАДН + H+
НАДН + H+ + КоQ = НАД+ + КоQ∙H2;
Комплекс II (сукцинат: КоQ-оксидоредуктаза) катализирует перенос электронов от сукцината к КоQ. ФАД и ФМН ковалентно связаны с дегидрогеназами:
S´H2 + ФАД = S´ + ФАД∙H2
ФАД∙H2 + КоQ = ФАД + КоQ∙H2;
Коплекс III (КоQН2: цитохром с-оксидоредуктаза) катализирует перенос электронов от КоQН2 к цитохрому с
КоQ∙H2 +2Fe3+ = КоQ +2Fe2+
Комплекс IV (цитохромоксидаза) катализирует перенос электронов от цитохрома с к кислороду:
2Fe2+ + 1/2O2 = 2Fe3+ +H2O
Изменение стандартные окислительные потенциалы основных компонентов дыхательной цепи ∆E0´:
НАД+/НАДН + H+ -0,32 В
ФАД/ФАД∙H2 -0,12
КоQ/КоQ∙H2 -0,05
Цит a3 (Fe2+/Fe3+) +0,55
1/2O2/H2O +0,82
!!! Если мы рассмотрим окислительно-восстановительные потенциалы каждой пары, то увидим, что они становятся всё более положительными, т.е. переносчики электронов располагаются в соответствии с их возрастающей способностью к восстановлению.
Если мы рассмотрим изменение стандартной свободной энергии ∆G0´при переносе электронов с:
НАДН → КоQ ∆G0´ = -51,4 кДж/моль
Цит b → Цит с ∆G0´ = -41,4 кДж/моль
Цит а → O2 ∆G0´ = -99,6 кДж/моль,
то можем отметить, что:
Во-первых, перенос электронов от одного компонента дыхательной цепи к другому сопровождается выделением свободной энергии, т.е. протекает самопроизвольно.
Во-вторых, благодаря участию промежуточных переносчиков энергия выделяется порциями.
В-третьих, если учесть, что на синтез одной молекулы АТФ требуется не менее 31 кДж/моль, то в дыхательной цепи есть три участка, высвобождающейся энергии достаточно для синтеза АТФ.
Таким образом, перенос пары электронов от НАД-зависимых дегидрогеназ даёт в итоге образование трёх молекул АТФ.
Окисление же ФАД-зависимых дегидрогеназ – только двух молекул АТФ, так как пара электронов в дыхательную цепь на уровне коэнзима Q, минуя первый участок сопряжения.
Механизм работы электронтранспортной цепи
Все вышеперечисленные компоненты дыхательной цепи встроены во внутреннюю мембрану митохондрий (рис. 8.6):

Рис. 8.7. Дыхательная или электрон-транспортная цепь клеток эукариот
1) НАД-зависимая дегидрогеназа расположена на матриксной поверхности внутренней мембраны митохондрий отдает пару электронов водорода на ФМН-зависимую дегидрогеназу.
При этом из матрикса пара протонов переходит также на ФМН и в результате образуется ФМН Н2. В это время пара протонов, принадлежащих НАД выталкивается в межмембранное пространство.
2) ФАД-зависимая дегидрогеназа отдает пару электронов на КоQ а пару протонов выталкивает в межмембранное пространство. Получив электроны КоQ принимает из матрикса пару протонов и превращается в КоQ Н2.
3) КоQ Н2 выталкивает пару протонов в межмембранное пространство, а пара электронов передается на цитохромы и далее на кислород с образованием молекулы воды.
!!! В итоге при переносе пары электронов по цепи из матрикса в межмембранное пространство перекачивается 6 протонов (3 пары), что ведет к созданию разницы потенциалов и разницы рН между поверхностями внутренней мембраны.
4) Разница потенциалов и разница рН обеспечивают движение протонов через протонный канал обратно в матрикс.
5) Такое обратное движение протонов ведет к активации АТФ-синтазы и синтезу АТФ из АДФ и фосфорной кислоты.
!!!! В результате функционирования ферментов цепи переноса электронов, локализованной во внутренней мембране митохондрий, создается протонный градиент, энергия которого используется для синтеза АТФ с помощью ферментной системы, называемой H+-зависимой-АТФ-синитазой (рис. 8.7):
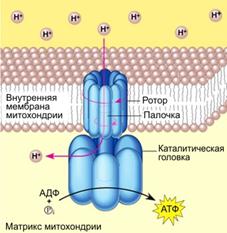
Рис 8.7. Модель молекулярной организации H+-зависимой-АТФ-синитазы
АТФ-синтаза состоит из двух частей:
- встроенного в мембрану протонного канала и
- каталитической субъединицы, которая выступает из мембраны в виде «головки».
Каталитический цикл, в результате которого происходит синтез АТФ, подразделяется на три фазы, каждая из которых проходит поочередно в трех активных центрах:
- в начале идет связывание АДФ и фосфата;
- затем образуется фосфоангидридная связь;
наконец, освобождается конечный продукт реакции - АТФ.
В клетках свободная энергия сохраняется благодаря сопряженному синтезу аденозинтрифосфата (АТР).
Образовавшаяся АТФ из матрикса в цитоплазму переносится ферментами транслоказами, при этом, в обратном направлении, в матрикс переносится одна молекула АДФ и одна молекула фосфорной кислоты.
– Конец работы –
Используемые теги: Лекция, нципы, метаболизма, биоэнергетики0.071
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Лекция 8 Принципы метаболизма и биоэнергетики
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.15 сек.
Новости и инфо для студентов