рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Философия
- /
- Структурно-функциональная характеристика спинного мозга
Реферат Курсовая Конспект
Структурно-функциональная характеристика спинного мозга
Структурно-функциональная характеристика спинного мозга - раздел Философия, Тема 6. Физиология спинного мозга Спинной Мозг Находится В Полости Позвоночника. Это Наиболее Примитивная И Наи...
Спинной мозг находится в полости позвоночника. Это наиболее примитивная и наиболее древняя часть ЦНС. Она мало дифференцирована. У взрослого человека длина спинного мозга достигает 45 см, а вес – 38 г и составляет 2% от веса всей ЦНС.
Таким образом, в спинном мозге насчитывается 31 сегмент. Каждый соответствует одному из позвонков. Корешки сегмента выходят из позвоночного канала через межпозвоночные отверстия. До входа в отверстие задние и передние корешки идут раздельно. Около отверстия на заднем корешке имеется вздутие – спинномозговой ганглий (ganglion spinale), в котором находятся тела псевдоуниполярных афферентных нервных клеток. Их аксоны и образуют задние («чувствительные») корешки спинного мозга. Передние двигательные корешки состоят из аксонов крупных клеток спинного мозга – мотонейронов. После выхода из межпозвоночных отверстий передние и задние корешки объединяются в смешанные нервы периферической нервной системы. Так как рост спинного мозга отстает во время, онтогенетического развития организма от роста позвоночника, имеется несоответствие между сегментами спинного мозга и одноименными позвонками. Поскольку нервы выходят из позвоночника через определенные отверстия, корешки удлиняются. Ниже второго поясничного позвонка полость позвоночника заполнена пучком корешков, спускающихся вниз и создающих так называемый конский хвост (cauda equina).
На поперечных срезах спинного мозга видно, что его периферию занимает белое вещество (substantia alba), а центральную часть – серое вещество (substantia grisea). Белое вещество состоит из покрытых миелином аксонов нервных клеток, объединенных в пучки восходящих и нисходящих проводящих путей спинного мозга, образующих задние, передние и боковые канатики (funiculis posterior, lateralis и ventralis). В задних канатиках промежуточная борозда разделяет каждую половину спинного мозга на два пучка: тонкий, или нежный (fasciculus gracilis), располагающийся медиальнее, и клиновидный (fasciculus cunea-tus), находящийся латеральнее.
Серое вещество формирует передние и задние серые столбы (columnae grisea ventralis et dorsalis). Правая и левая половины соединяются между собой спайкой (commissura). В грудном и верхних сегментах поясничного отдела есть еще и боковые столбы серого вещества (columnae lateralis). На поперечных срезах столбы серого вещества выглядят как выступы и называются рогами. Передние рога (cornua ventralis) массивнее задних (cornua dorsales). На верхушке дорсального рога находится желатинозная субстанция Роланда. Она состоит из мелких клеток, создающих своими ветвящимися отростками густую сеть (нейропиль). Между рогами располагается центральная часть серого вещества (промежуточная зона). Нервные клетки серого вещества группируются в ядра, которые вытягиваются вдоль спинного мозга и имеют вид веретена (иногда ядра спинного мозга называют столбами). Наибольшее количество ядер сосредоточено в передних рогах и в промежуточной зоне серого вещества. Так, например, в промежуточной зоне выделяют промежуточное ядро Кахаля, столб Кларка и т. д. В белом веществе наблюдают островки серого вещества. Их совокупность определяют как ретикулярную формацию (formatio reticularis).
В спинном мозге человека насчитывается 13,5 млн. нервных клеток различных по форме, размерам и длине отростков. Наиболее крупные клетки с длинными аксонами, покидающими ЦНС в составе передних корешков, располагаются в передних столбах серого вещества. Это – мотонейроны (их аксоны направляются к скелетным мышцам). Мотонейроны составляют лишь 3% общего числа нейронов спинного мозга. Основная масса нейронов называется проприонейронами (их отростки не выходят за пределы центральной нервной системы). Проприонейроны в свою очередь подразделяются на интернейроны и канатиковые нейроны. Интернейроны (вставочные клетки) – мелкие клетки с короткими отростками, не покидающими серого вещества. У канатиковых («пучковых») нейронов размеры сомы больше, отростки, выходящие из серого вещества, и некоторые проводящие пути белого вещества длиннее. Белое вещество представлено путями восходящими (к вышележащим сегментам спинного мозга и к головному мозгу), а также нисходящими (от головного мозга и верхних сегментов). Наличие двусторонних связей обеспечивает системную организацию мозговых структур.
Тела чувствительных клеток вынесены за пределы спинного мозга (рис.17). Часть из них располагается в спинальных ганглиях. Это тела соматических афферентов, иннервирующих в основном скелетные мышцы. Другие находятся в экстра- и интрамуральных ганглиях автономной нервной системы и обеспечивают чувствительность только внутренних органов.
Чувствительные клетки имеют один отросток, который вскоре после отхождения от тела клетки делится на две ветви.
Одна из них проводит возбуждение от рецепторов к телу клетки, другая - от тела нервной клетки к нейронам спинного или головного мозга. Распространение возбуждения из одной ветви в другую может происходить без участия тела клетки.
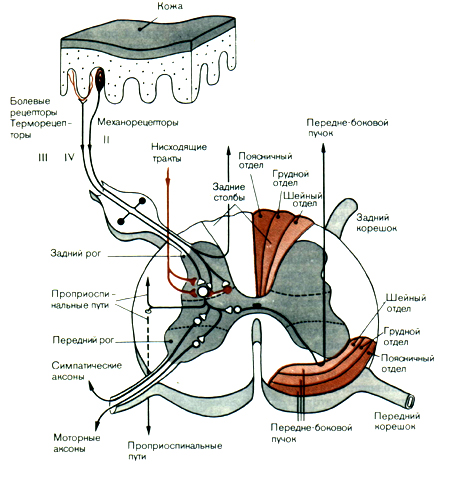
Рис.17. Поперечное сечение спинного мозга
и связи кожных афферентов в спинном мозгу.
Нервные волокна чувствительных клеток по скорости проведения возбуждения и диаметру классифицируют на А-, В- и С-группы. Толстые миелинизированные А-волокна с диаметром от 3 до 22 мкм и скоростью проведения возбуждения от 12 до 120 м/с разбиваются еще на подгруппы: альфа- волокна от мышечных рецепторов, бета - от тактильных рецепторов и барорецепторов, дельта - от терморецепторов, механорецепторов, болевых рецепторов. К волокнам группы В относят миелиновые отростки средней толщины со скоростью проведения возбуждения 3-14 м/с. По ним в основном передается ощущение боли. К афферентным волокнам типа С относится большинство безмиелиновых волокон толщиной не более 2 мкм и скоростью проведения до 2 м/с. Это волокна от болевых, хемо- и некоторых механорецепторов.
Сам спинной мозг в целом содержит, например, у человека примерно 13 млн. нейронов. Из их общего числа только около 3% составляют эфферентные, двигательные или моторные нейроны, а остальные 97 % приходятся на долю вставочных, или интернейронов. Двигательные нейроны являются выходными клетками спинного мозга. Среди них различают альфа- и гамма-мотонейроны, а также преганглионарные нейроны автономной нервной системы.
Альфа-мотонейроны осуществляют передачу скелетным мышечным волокнам сигналов, выработанных в спинном мозгу. Аксоны каждого мотонейрона многократно делятся, и, таким образом, каждый из них охватывает своими терминалями до сотни мышечных волокон, образуя в совокупности с ними двигательную единицу. В свою очередь, несколько мотонейронов, иннервирующих одну и ту же мышцу, образуют мотонейронный пул, В его состав могут входить мотонейроны нескольких соседних сегментов. В связи с тем что возбудимость мотонейронов пула неодинакова, при слабых раздражениях возбуждается только часть из них. Это влечет за собой сокращение лишь части мышечных волокон. Другие моторные единицы, для которых это раздражение является подпороговым, тоже реагируют, хотя их реакция выражается лишь в деполяризации мембраны и повышении возбудимости. С усилением раздражения они еще больше вовлекаются в реакцию, и, таким образом, все двигательные единицы пула участвуют в рефлекторном ответе.
Максимальная частота воспроизведения ПД в альфа-мотонейроне не превышает 200-300 имп/с. Вслед за ПД, амплитуда которого составляет 80-100 мВ, возникает следовая гиперполяризация длительностью от 50 до 150 мс. По частоте импульсации и выраженности следовой гиперполяризации мотонейроны разделяют на две группы: фазические и тонические. Особенности их возбуждения коррелируют с функциональными свойствами иннервируемых мышц. Фазически-ми мотонейронами иннервируются более быстрые, "белые" мышцы, тоническими - более медленные, "красные".
В организации функции альфа-мотонейронов важным звеном является наличие системы отрицательной обратной связи, образованной аксонными коллатералями и специальными тормозными вставочными нейронами - клетками Реншоу. Своими возвратными тормозными влияниями они могут охватывать большие группы мотонейронов, обеспечивая, таким образом, интеграцию процессов возбуждения и торможения.
Гамма-мотонейроны иннервируют интрафузальные (внутриверетенные) мышечные волокна. Они разряжаются с более низкой частотой, и следовая гиперполяризация у них выражена слабее, чем у альфа-мотонейронов. Их функциональное значение сводится к сокращению интрафузальных мышечных волокон, не приводящему, однако, к появлению двигательного ответа. Возбуждение этих волокон сопровождается изменением чувствительности их рецепторов к сокращению или расслаблению экстрафузальных мышечных волокон.
Нейроны автономной нервной системы составляют особую группу клеток. Тела симпатических нейронов, аксоны которых являются преганглионарными волокнами, располагаются в интермедиолатеральном ядре спинного мозга. По своим свойствам они относятся к группе B-волокон. Характерной особенностью их функционирования является низкая частота свойственной им постоянной тонической импульсной активности. Одни из этих волокон участвуют в поддержании сосудистого тонуса, другие - обеспечивают регуляцию висцеральных эффекторных структур (гладкой мускулатуры пищеварительной системы, железистых клеток).
Тела парасимпатических нейронов образуют крестцовые парасимпатические ядра. Они располагаются в сером веществе крестцовых сегментов спинного мозга. Для многих из них характерна фоновая импульсная активность, частота которой возрастает по мере повышения давления в мочевом пузыре. При раздражении висцеральных тазовых афферентных волокон в этих эфферентных клетках регистрируется вызванный разряд, характеризующийся чрезвычайно большим латентным периодом.
К вставочным, или интернейронам, спинного мозга относятся нервные клетки, аксоны которых не выходят за его пределы. В зависимости от хода отростков различают собственно спинальные и проекционные. Спинальные интернейроны ветвятся в пределах нескольких смежных сегментов, образуя внутрисегментарные и межсегментарные связи. Наряду с ними существуют интернейроны, аксоны которых проходят через несколько сегментов или даже из одного отдела спинного мозга в другой. Их аксоны образуют собственные пучки спинного мозга.
К проекционным интернейронам относятся клетки, длинные аксоны которых формируют восходящие пути спинного мозга. На каждом интернейроне расположено в среднем около 500 синапсов. Синаптические влияния в них опосредуются через ВПСП и ТПСП, суммация которых и достижение критического уровня приводят к возникновению распространяющегося ПД.
УчебныЙ вопрос № 2
– Конец работы –
Эта тема принадлежит разделу:
Тема 6. Физиология спинного мозга
САНКТ ПЕТЕРБУРГСКИЙ УНИВЕРСИТЕТ... ГОСУДАРСТВЕННОЙ ПРОТИВОПОЖАРНОЙ СЛУЖБЫ...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Структурно-функциональная характеристика спинного мозга
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.453 сек.
Новости и инфо для студентов