Кінетичні моделі популяції клітин
В основі процесів обміну клітини з середовищем і внутрішнього метаболізму лежить складна мережа організованих певним чином в часі і просторі різних реакцій. В результаті цих процесів змінюються концентрації різних речовин, чисельність окремих кліток, біомаса організмів, можуть змінюватися і інші величини, наприклад величина трансмембранного потенціалу в клітці. Зміни всіх цих величин в часі і складають кінетику біологічних процесів. Основні початкові передумови при описі кінетики в біологічних системах загалом такі ж, як і в хімічній кінетиці.
Розглянемо простий приклад замкнутої популяції кліток, в якій одночасно відбуваються процеси розмноження і загибелі і в надлишку є поживні речовини. Виникає питання, як міняється чисельність кліток в такій системі з часом і чи може в ній врешті-решт встановитися стаціонарний стан, коли число кліток мінятися не буде. Це типова кінетична задача, яка розв'язується за допомогою звичайних диференціальних рівнянь. Нехай в деякий момент часу t концентрація кліток в середовищі складе N. Швидкість dN/dt зміни концентрації кліток у середовищі складається з швидкості їх розмноження Vрозмн і швидкості загибелі Vзагиб:
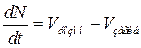 . (6)
. (6)
У простому випадку швидкість розмноження, тобто збільшення концентрації кліток в одиницю часу, пропорційна їх чисельності в кожен момент:
 , (7)
, (7)
де k1 – константа пропорційності, що залежить від умов середовища (температура, наявність поживних речовин та ін.). Аналогічно
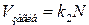 , (8)
, (8)
де k2 – константа пропорційності, що визначає інтенсивність процесів загибелі кліток. Звідси витікає, що
 , (9)
, (9)
де 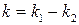 .
.
Розв’язавши це рівняння, ми знайдемо, як змінюється концентрація клітин в середовищі з часом N = N(t):
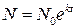 , (10)
, (10)
де N0 – концентрація кліток в початковий момент часу t = 0 спостереження за системою.
Можна побачити, що в залежності від величин констант швидкостей процесів загибелі k2 і розмноження k1 доля цієї популяції буде різною. Якщо  , то
, то  і в системі відбуватиметься необмежене зростання числа кліток з часом:
і в системі відбуватиметься необмежене зростання числа кліток з часом: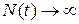 , а якщо
, а якщо 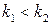 , то
, то  , то з часом популяція вимиратиме:
, то з часом популяція вимиратиме:  . І лише в окремому випадку при
. І лише в окремому випадку при 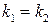 число кліток залишатиметься постійним:
число кліток залишатиметься постійним: 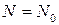 .
.
Іншим прикладом моделі зростання популяції в середовищі з обмеженою кількістю поживних речовин служить рівняння логістичної кривої. Логістичне рівняння Ферхлюста має вид:
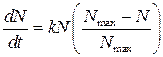 . (11)
. (11)
 Тут Nmax – максимальна чисельність популяції, що можлива в даних умовах. Крива N =N(t), що описується цим рівнянням, має дві характерні ділянки (див. рис.2). У початковий період зростання, коли N < Nmax, крива носить експоненційний характер. Потім, після точки перегину, нахил поступово зменшується і крива наближається до верхньої асимптоти N = Nmax, тобто до максимально досяжного рівня популяції в даних умовах.
Тут Nmax – максимальна чисельність популяції, що можлива в даних умовах. Крива N =N(t), що описується цим рівнянням, має дві характерні ділянки (див. рис.2). У початковий період зростання, коли N < Nmax, крива носить експоненційний характер. Потім, після точки перегину, нахил поступово зменшується і крива наближається до верхньої асимптоти N = Nmax, тобто до максимально досяжного рівня популяції в даних умовах.
Як видно, динаміку біологічних процесів можна описувати рівняннями, що аналогічні рівнянням хімічної кінетики. Проте, в порівнянні зі звичайною хімічною кінетикою, біологічна кінетика характеризується наступними особливостями:
1. Як змінні виступають не тільки концентрації речовин, але і інші величини.
2. Змінні змінюються не тільки в часі, але і в просторі.
3. Біологічна система просторово гетерогенна, і умови взаємодії реагентів можуть бути різні в різних точках системи.
4. Існують спеціальні механізми саморегуляції, що діють за принципом зворотнього зв'язку.
Основна задача в біофізиці складних систем полягає в тому, щоб одержати характеристики різних динамічних режимів і з'ясувати умови і значення параметрів, при яких вони реалізуються в живій клітині.