Часть I. Палеонтология
Предисловие
Одним из важнейших методов усвоения материала считается самостоятельная работа студента с учебником. В свое время получил распространение учебник «Историческая геология с основами палеонтологии», подготовленный на кафедре исторической геологии и палеонтологии С.-Петербургского Горного института Е.В. Владимирской, А.Х. Кагармановым, Н.Я. Спасским и др. Этот учебник, изданный около 30 лет назад, в значительной мере устарел. Более новым является учебное пособие В.М. Подобиной и С.А. Родыгина «Историческая геология» (2000), изданное кафедрой общей и исторической геологии ТГУ, однако раздел «Палеонтология» в нем отсутствует. В настоящее время Министерством образования и науки РФ утвержден учебник «Историческая геология» (издание 4-е, 2008) Н.В. Короновского, В.Е. Хаина, Н.А. Ясаманова объемом 29 усл. печ. листов. Учебник написан с позиций признания глобальной тектоники плит, хотя часть российских ученых, до сих пор, считает, что эта теория не объясняет всего комплекса геологических процессов планеты Земля.
Палеонтология хорошо освещена в двухтомном учебнике «Палеонтология» сотрудников кафедры палеонтологии МГУ И.А. Михайловой и О.Б. Бондаренко (1997) (62 усл. печ. листа). Значительный объем имеет учебное пособие М.Д. Парфёновой «Историческая геология с основами палеонтологии» (1998), подготовленное на кафедре общей и исторической геологии ТПУ. К сожалению, в настоящее время оно также устарело. Перечисленные учебники и учебные пособия, рассчитанные на большой объем курсов «Историческая геология» и «Палеонтология», мало пригодны при значительном сокращении этих курсов. С целью восполнения этого пробела было опубликовано учебное пособие «Историческая геология» Н.В. Гумеровой (2010) объемом около 6 усл. печ. листов. Недостатком пособия является слабое освещение вопросов геодинамики и отсутствие предусмотренного программой раздела «Палеонтология».
В публикуемом пособии «Историческая геология с основами палеонтологии» эти недочеты устранены. Учитывая, что значительная часть выпускников института природных ресурсов будут работать в Кузбассе, дана история геологического развития этого крупнейшего угольного бассейна России.
Авторы благодарны научному редактору профессору А.А. Поцелуеву, а также преподавателям кафедры географии, геологии и методики преподавания географии Куз. ГПА и кафедры геологии и геодезии СГИУ за ценные замечания и уточнения.
Введение
Среди наук геологического цикла палеонтология и историческая геология характеризуются определенным сходством стоящих перед ними задач и методов их решения. Объектом исследования этих отраслей знания являются горные породы с заключенными в них остатками организмов геологического прошлого Земли. Историческая геология ставит своей задачей изучение эволюционного развития планеты Земля, которое по современным представлениям носит циклично-поступательный характер, отражающий характер развития космического пространства. Воздействием космического фактора, вероятно, обусловлена периодическая активизация эндогенных процессов, установленная профессором С.Г. Неручевым.
Палеонтология изучает историю развития живого вещества планеты Земля. В настоящее время она неразрывно связана с учением о биосфере, сформулированным В.И. Вернадским в начале ХХ века.
Другим важным направлением палеонтологии является учение о механизме возникновения жизни. Подробно гипотезы возникновения жизни на Земле будут рассмотрены в разделе «Архей и ранний протерозой».
Важным элементом палеонтологии было учение о неравномерном развитии органического мира Земли, впервые сформулированное австрийским ученым Э. Зюссом в начале ХХ века. В середине XIX века «теория божественных катастроф», когда бог уничтожает весь ранее созданный органический мир и создает новый, рассматривалась как альтернатива эволюционистской концепции, предложенной Ч. Дарвиным. Она была отвергнута после окончательной победы идей эволюционизма. Время показало, что в истории Земли длительные этапы сравнительно медленного развития органического мира чередовались с кратковременными этапами кардинальных перестроек (в XIX веке эти моменты назывались «катастрофами»). Таким образом, в развитии органического мира присутствуют и катастрофы и длительные периоды эволюции.
В настоящее время причины экологических кризисов и катастроф также как и возрастные границы этапов, разными исследователями трактуются по-разному, но принцип этапности (периодически и закономерно повторяющихся этапов эволюционного процесса) признается практически всеми.
Часть I. Палеонтология
Палеонтология – наука, изучающая органический мир прошлых геологических эпох и… По характеру объектов исследования палеонтология подразделяется на палеозоологию и палеоботанику, изучающие,…Семейство
род,
Вид.
С учетом приставок над- и под- их число возрастает в 2-3 раза. В данной работе таксономический ранг ниже отряда обычно не рассматриваются. В отдельных случаях тип подразделяется на инфратипы. Реальным подразделением является вид, существующий в природе в виде одной или нескольких популяций. Остальные подразделения выделяются более или менее условно, причем разными исследователями – по-разному.
Система соподчиненных таксонов животных и растений – номенклатура, предложенная К. Линнеем более 200 лет назад, сохранилась до настоящего времени в качестве обязательных «Международных правил номенклатуры». Согласно этим «Правилам» при наименовании какого-либо организма используется тройное название: указывается род, вид и фамилия автора, впервые описавшего этот вид. В случае приближенных определений используется так называемая «открытая номенклатура». При невозможности определения видовой принадлежности употребляется понятие «spesies» (sp.), при сомнительных определениях – «conformis» (cf.), «affinis» (aff.), «ex grege» (ex gr.)
2. Современная классификация органическоо мира
Согласно представлениям В.И. Вернадского, Земной шар состоит из живого и неживого вещества. Живое вещество Земли, по систематике сотрудников МГУ, выделяется в империю Жизнь, в состав которой входят надцарства Доядерные (Прокариоты) и Ядерные (Эукариоты) организмы (Приложение 1, рис. 1).
2.1. Надцарство Доядерные организмы. Прокариоты (Procaryota)

К надцарству принадлежат наиболее примитивные одноклеточные и многоклеточные организмы, не имеющие обособленного ядра. Генетическая информация заключена в единственной хромосоме (рис.1 а). Размеры прокариот от 0,015 мкм до 20 см. Они появились в интервале 3,7-3,1 млрд. лет. Питание прокариот осуществляется за счет хемосинтеза и фотосинтеза. Надцарство подразделяется на царства Бактерии и Цианобионты (Приложение 1).
2.1.1. Царство Бактерии (Bacteria)
Бактерии представляют собой микроскопические организмы размером 1-5 мкм, но иногда достигают 10000 мкм (1 см). Среди бактерий встречаются автотрофные и гетеротрофные группы. Роль бактерий велика в образовании месторождений полезных ископаемых. Достоверные находки бактерий известны в породах, имеющих возраст около 3,5 млрд. лет. В настоящее время бактерии населяют все среды обитания. Некоторые исследователи объединяют бактерии с вирусами, другие выделяют вирусы в качестве самостоятельного царства, рассматривая их как доклеточную форму жизни.
Царство Цианобионты (Cyanobionta)
А
Рис. 2. Строматолиты:
а – столбчатый строматолит (в разрезе)
б – колония современных цианобионтов в заливе
Шарк (Австраия)
Б
В настоящее время изучается возможность разработки на основе строматолитов относительной геохронологии докембрийских толщ.
Появившись около 3,5 млрд. лет назад, цианобионты, благодаря наличию хлорофилла, явились первыми на Земле продуцентами биогенного кислорода. Современные цианобионты населяют зону мелководья преимущественно до глубины 20 м. Они хорошо переносят колебания солености и температуры водоема. Отсутствие ядра сближает цианобионтов с бактериями, а способность к фотосинтезу – с водорослями. По уровню организации цианобионты превосходят бактерии, но уступают водорослям.
2.2. Надцарство Ядерные организмы или Эукариоты (Eucaryota)
Эукариоты, имеющие в отличие от прокариот обособленное ядро, появились в раннем протерозое (1,7-1,5 млрд. лет назад) и обычно рассматриваются как потомки одной из групп прокариот. ДНК эукариот была сосредоточена в хромосомах, находящихся внутри клеточного ядра, окруженного ядерной оболочкой. Ядро эукариот управляло, таким образом, всей жизнедеятельностью клетки (рис. 1 б).
Размеры эукариот варьируют от 10 мкм (Одноклеточные) до 33 м (длина Китообразных) и 100-110 м (высота метасеквой). Эукариоты подразделяются на царства Растения, Грибы и Животные (Приложение 1), однако на ранних стадиях эволюции этих царств их различия проявляются слабо. В частности, среди Жгутиковых, отнесенных к царству животных, содержатся группы, имеющие признаки растений и животных. Некоторые исследователи объединяют прокариот и наиболее примитивных эукариот в особое царство Протисты.
Царство Растения (Phyta)
Растения – автотрофные одноклеточные и многоклеточные организмы – подразделяются на подцарства Низшие растения (водоросли) и Высшие растения, характеризующиеся, в основном, наземным образом жизни (Приложение 1, 2).
Подцарство Низшие растения (Thallophyta)
Отдел Красные(Багряные) водоросли (Rhodophyta), по многим признакам близкий к цианобионтам, включает многоклеточные, редко одноклеточные формы,… Отдел Диатомовые водоросли (Diatomeae) (рис.4) представлен одноклеточными… Отдел Золотистые водоросли (Chrysophyta) в настоящее время представлен колониальными, преимущественно одноклеточными…А б а в
Рис. 3. Золотистые водоросли Рис. 4. Диатомовые водоросли
а – кокколитовые (Q), а – род Arachnodiscus ( P ),
б - кремнистые жгутиковые (Q) б – колония рода Chaetoceras (Q),
в – род Melosira (N)


а б
|
Отдел Харовые водоросли (Charopyta) включает многоклеточные организмы, близкие к зеленым водорослям и нередко объединяемые с ними в одну группу. По многим признакам напоминают хвощи. В ископаемом состоянии известны с силурийского периода.
Подцарство Высшие растения (Telomophyta)
Надотдел Споровые растения (Sporophyta) Споровые растения появились в середине силурийского периода и сохранились в настоящее время, уступая, однако, по…А б
Рис. 8. Современный хвощ: а – спорангии на вершине стебля,
б – стебель с листьями
нередко разрушенная, и кора. Листья имели различную форму: от игольчатых до ланцетовидных (рис. 8 а, б). Спорангии располагались на
концах побегов (рис. 8 а). В ископаемом состоянии сохраняются слепки внутренней полости стволов, отпечатки листьев, остатки спорангиев. Особенности их строения положены в основу разделения отдела на несколько порядков. Наибольшее значение для стратиграфии имеют порядки: Клинолистники, Каламиты и Хвощевые, являвшиеся наряду с лепидодендровыми основными углеобразователями. Порядок Клинолистники (D3 – P, возможно T) объединяет травянистые лианоподобные растения с членистым продольноребристым ветвящимся стеблем. Листья клиновидной формы, иногда рассеченные. Порядок Каламиты (D3 – P) представлен вымершими деревьями высотой до 20 м, напоминавшими по внешнему облику гигантские хвощи (рис. 11, 4). Листья узкие ланцетовидные. Спорангии имели колосковидную форму. В ископаемом состоянии встречаются ядра сердцевины, отпечатки стволов и листьев. Порядок Хвощевые (C – ныне, расцвет C – P1) объединяет небольшие травянистые растения, типичным представителем которых является современный хвощ.

 Класс Папоротниковидные(Pteropsida) (D2 – ныне, расцвет C – P – T1) включает травянистые, лианоподобные и древесные формы. В современной флоре преобладают травянистые растения. Древовидные и лианоподобные папоротники встречаются как реликты лишь в субтропиках. Размеры растений варьируют от нескольких миллиметров до 20-30 м. Нижняя часть стебля разветвлена, образуя корневище. Лист папоротника (вайя), имеет сложно рассеченную листовую пластину, состоящую из перьев, отходящих от главной оси (рахиса) (рис. 9). Листья весьма крупные, в палеозое длина листа некоторых папоротников достигала 5м. Спорангии располагаются на нижней стороне листа (рис.10). Папоротниковидные, произошли, по-видимому,
Класс Папоротниковидные(Pteropsida) (D2 – ныне, расцвет C – P – T1) включает травянистые, лианоподобные и древесные формы. В современной флоре преобладают травянистые растения. Древовидные и лианоподобные папоротники встречаются как реликты лишь в субтропиках. Размеры растений варьируют от нескольких миллиметров до 20-30 м. Нижняя часть стебля разветвлена, образуя корневище. Лист папоротника (вайя), имеет сложно рассеченную листовую пластину, состоящую из перьев, отходящих от главной оси (рахиса) (рис. 9). Листья весьма крупные, в палеозое длина листа некоторых папоротников достигала 5м. Спорангии располагаются на нижней стороне листа (рис.10). Папоротниковидные, произошли, по-видимому,
|

от риниофит. Это были не высокие деревья с мощными стволами диаметром до 1 м (рис.11, 3). В карбоне папоротниковидные дали начало голосеменным.

Рис. 11 . Реконструкция леса карбона и ранней перми.
Плауновидные: 1 – сигиллярия, 2 – лепидодендрон; папоротниковидные: 3 - древовидный папоротник; членистостебельные: 4 – каламитон
Надотдел Семенные растения
Отдел Голосеменные (Пинофиты) (Pinophyta)включает деревья, кустарники, реже травянистые растения. Они имели уже настоящую всасывающую корневую… КлассЦикадоподобные (Cycadopsida)включает порядки Семенные папоротники,… Наибольшее сходство с папоротниковидными имел порядок Семенные папоротники (Птеридоспермы). При находках листьев без…Рис. 16. Ископаемая шишка хвойных Рис. 17. Отпечатки листьев хвойных

Рис.18. Ветка современной сосны
1 – женские шишки, 2 –мужские шишки,
3 – семя, переносимое по ветру
|

Рис. 19. Реконструкция леса середины мезозоя.
Цикадоподобные: 1 - семенные папоротники, 2 - беннеттитовые, 3 – цикадовые;
4 – Пинопсиды; 5 – Гинкгоподобные
ОтделПокрытосеменные (Магнолиофиты) (Magnoliophyta)занимают господствующее положение в современной флоре, обитая на всех широтах Земного шара. В состав отдела входят листопадные и вечнозеленые, травянистые и древесные растения, кустарники и лианы. Весьма разнообразны форма листьев, характер их жилкования, расположение на стеблях. От других отделов высших растений магнолиофиты отличаются наличием цветка, двойным оплодотворением, наличием сосудистой системы. Цветок покрытосеменных растений состоит из тычинок с пыльцой, пестика с завязью, лепестков и чашелистиков.Тычинки – мужские половые органы, продуцируют пыльцу, для оплодотворения пестика – женского органа.Лепестки и чашелистики – видоизмененные листья, защищающие половые органы (рис.20).
Эволюция цветковых связана с насекомыми, участвующими в их опылении (рис. 21). Покрытосеменные произошли от беннеттитовых в меловом периоде. В настоящее время представлены классами Двудольных и Однодольных, объединяющими около 235000 видов. Классы делятся на подразделения низших рангов. Цветковые участвуют в образовании торфа и бурых углей.
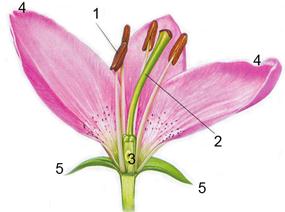
|

Рис. 20. Цветок:
1 – тычинки с пыльниками,
2 – пестик, 3 – завязь,
4 – лепестки, 5 – чашелистики
Класс Двудольные(Dicotyledones) включает более 70% представителей магнолиофитов: трав, кустарников, лиан и деревьев. Они имеют две семядоли, четырех- или пятичленный цветок, листья обычно с сетчатым жилкованием. Особенности жилкования листьев являются важным систематическим признаком вымерших видов.
Класс Однодольные(Monocotyledones) в большинстве случаев представлен травянистыми растениями, редко кустарниками и деревьями. Общими признаками являются наличие у зародыша одной семядоли и трехчленная форма цветка. Форма листьев удлиненная лентовидная, стреловидная, иногда веерная и эллиптическая. Жилкование преимущественно параллельное, дуговидное, веерное, реже сетчатое.
Царство Грибы (Fungi)
Вопросы для самоконтроля: 1. Изложите современные представления об этапном характере эволюции органического мира Земли.Царство Животные (Zoa)
Характерными признаками царства Животные являются гетеротрофный тип питания и подвижность, хотя многие представители ведут, в основном, прикрепленный образ жизни. При классификации животных используются разные признаки, в силу чего количество одноранговых подразделений велико (выделяется от 16 до 25 типов). Появление животных предположительно датируется началом позднего протерозоя.
Подцарство Одноклеточные (Простейшие) (Protozoa)
Тип Саркодовые (Sarcodina) характеризуется наличием ложноножек (псевдоподий),… К классу Фораминиферы (Foraminifera)относится большинство саркодовых, имеющих раковину с тонкими отверстиями, через…Рис. 22. Агглютинированные б
Раковины фораминифер
ввв вв а Рис. 23 . Известковые раковины фораминифер:А б
Рис. 24. Радиолярии:
(а, в) – скелеты радиолярий
б – поперечное сечение скелета радиолярий
|
Кремнистый скелет сохраняется в ископаемом состоянии, целестиновый – растворяется после смерти организма. Скелет имеет сферическую, звездчатую, шлемообразную или колоколообразную форму (рис. 24 а,в). Он состоит из вложенных один в другой решетчатых элементов и длинных радиальных игл (спикул), отходящих от центра (рис. 24 б). По особенностям строения скелета разные авторы выделяют от 2 до 5 отрядов радиолярий. Скелет частично выполняет защитные функции, но главное – увеличивает поверхность тела, обеспечивая парение в толще воды.
Появление радиолярий датируется ранним палеозоем, но есть данные о находках этих организмов в позднем докембрии. В современных морях радиолярии распространены в низких широтах, где накапливаются их скелетные остатки, образуя радиоляриевые илы. Стратиграфическую значимость радиолярии получают, начиная с мезозоя. В настоящее время насчитывается не менее 7000 видов этого класса, в то время как в ископаемом состоянии известно около 1000.
2.2.3.2. Подцарство Многоклеточные(Metazoa)
В отличие от простейших тело многоклеточных состоит из значительного количества клеток и межклеточного вещества, образующих ткани и органы. Выделяется 4 типа тканей, различающихся строением и функциями: эпителиальная, соединительная, мышечная и нервная. Органы многоклеточных, состоящие из комплекса тканей, регулируются нервной и кровеносной системами. Для многоклеточных характерно половое размножение, но у примитивных (низших) многоклеточных может наблюдаться чередование полового и бесполого.
Надраздел Низшие многоклеточные (Parazoa)
К примитивным многоклеточным относятся губковые, археоциаты и некоторые группы организмов неясного систематического положения (Приложение 3).
В состав типа Губковые (Spongia)входят морские и пресноводные прикрепленные бентосные организмы, тело которых состоит из 2 слоёв. Между слоями находится желеобразное вещество мезоглея, в которой формируется скелет, состоящий из иголочек (спикул). По способу питания губковые фильтраторы. Тело пронизано системой каналов, по которым поступает вода, обеспечивающая поступление пищи. Устья каналов (поры) обусловили другое название типа – Porifera (Пороносцы). До середины ХХ века в состав порифер включались археоциаты, позднее выделенные в особый тип. В последние годы появляются данные о целесообразности объединения губок и археоциат в тип Porifera. Согласно современной классификации Губковые подразделяются на три класса, из которых наибольшее распространение (около 10000 видов) имеет класс Губки. Остальные два класса (Коралловые губки и Сфинктозоа) встречаются сравнительно редко и в данной работе не рассматриваются.
По химическому составу скелета класс Губки расчленяется на подклассы Кремневые (Опаловые) и Известковые губки. Более дробная классификация основывается на структуре спикул. В ископаемом состоянии кремневые губки известны с кембрия, а известковые – с девона. Имеются указания о находках спикул в отложениях венда. Известковые губки участвуют в формировании рифов, кремневые губки современных осадков в процессе диагенеза могут превращаться в яшмы,  трепелы, опоки.
трепелы, опоки.


А б в
Рис. 25. Тип Губковые: а – вымершая одиночная губка (K),
б – колония вымерших губок,
в – современная морская губка
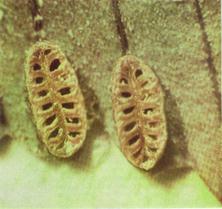
Тип Археоциаты (Archaeocyathi) (С) – вымершие бентосные (обычно прикрепленный бентос) одиночные и колониальные организмы губкоподобного облика. Наличие пор, в которые поступала вода, и центрального отверстия, через которое она выливалась, сближает археоциат с губками. Однако, в отличие от губок, скелет археоциат был не спикульный, а зернистый и только известковый. Одиночные археоциаты имели кубкообразную, реже цилиндрическую или блюдцеобразную форму. Диаметр кубка составлял 1-5 см, высота – 6-10 см. Колониальные археоциаты имели ветвистый, иногда массивный облик. Скелет археоциат обычно состоял из внутренней и наружной стенок, в промежутке между которыми располагалось тело организма, заполненное вертикальными перегородками (класс Правильные) илистерженьками (класс Неправильные) (рис. 26 1,2).
При благоприятных условиях скопления археоциат превращались в древнейшие рифы (рис. 27). Археоциаты являлись одной из быстро эволюционирующих групп морской фауны. Они появились в начале раннекембрийской эпохи, достигли расцвета и вымерли до её окончания. Не исключено, что вымирание археоциат на ранне-среднекембрийской границе было, в некоторой мере, кажущимся.
Возможно известковые губки, достоверно известные с девонского периода, были генетически как-то связаны с археоциатами. В настоящее время найдена губка Vacletia, морфологически сходная с археоциатами. На современном уровне знаний очевидна стратиграфическая значимость археоциат для раннекембрийской эпохи.
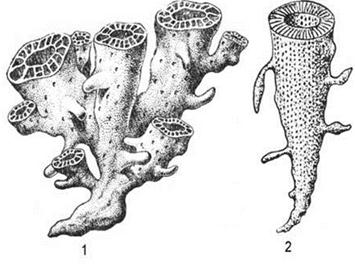
Рис. 26. ТипАрхеоциаты:
1 – колония археоциат, 2 – одиночная форма археоциат

Рис. 27. Рифовый известняк, образованный скелетами археоциат
Надраздел Настоящие многоклеточные (Высшие) (Eumetazoa)
Настоящие многоклеточные объединяют организмы, характеризующиеся дифференцированными тканями и органами. Они развиваются из двух или трех зародышевых листков (экто-, эндо- и мезодермы), соответственно, подразделяясь на радиально- и двусторонне-симметричных (Приложение 4).
Раздел Радиально-симметричные (Radiata)
Радиально-симметричные организмы (радиалии) обычно принадлежат к прикрепленному бентосу или планктону. Из ископаемых представителей радиалий наибольшее распространение имеет тип Стрекающие.
Стрекающие (Cnidaria)обитают исключительно в водной среде, хотя некоторые виды могут находиться на суше во время отлива. Лишь небольшая часть животных приспособилась к жизни в пресных водоемах (гидры и гидромедузы). Уникальная черта стрекающих — наличие стрекательных клеток, которые они используют для охоты и защиты от хищников, что и послужило поводом для названия этого типа. Каждая стрекательная клетка снабжена чувствительным волоском, который отвечает за выброс стрекательной нити, через которую в организм жертвы и попадает яд (рис. 28). Описано около 11 тысяч видов современных и вымерших книдарий.
Жизненный цикл стрекающих имеет две стадии: свободно плавающую форму медузы и прикрепленную полипов (рис. 29). Многие виды чередуют бесполую стадию полипа и половую медузы, у некоторых цикл лишён одной из стадий. Тело книдарий обладает радиальной симметрией и представляет собой открытый на одном конце двухслойный мешок (кишечную полость), другой конец замкнут. Единственное отверстие, ведущее в полость тела (рот), используется для дыхания, пищеварения и выделения. Вокруг рта располагается венчик щупалец со стрекательными клетками. Все книдарии хищники, имеют примитивную нервную систему, состоящую из отдельных нервных клеток.
Многие виды книдарий образуют колонии (обычно путём почкования), являющие собой единый организм, состоящий либо из медузоподобных, либо полипоподобных особей. В колонии все особи находятся в органической связи друг с другом, осуществляя общий обмен веществ, направленный на поддержание жизнедеятельности колонии. У колониальных полипов, кроме того, развивается общий скелет. Во время полового размножения в колонии образуются мужские и женские половые клетки, которые выпускаются в воду, где и происходит оплодотворение. Образуется личинка – планула, которая является расселительной формой. Она некоторое время плавает, затем прикрепляется к субстрату, давая начало полипу (рис. 29).
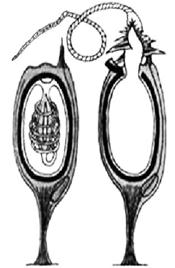
Рис. 28. Стрекательные клетки книдарий:
а – стрекательная клетка в покое,
б – стрекательная клетка в действии
А б
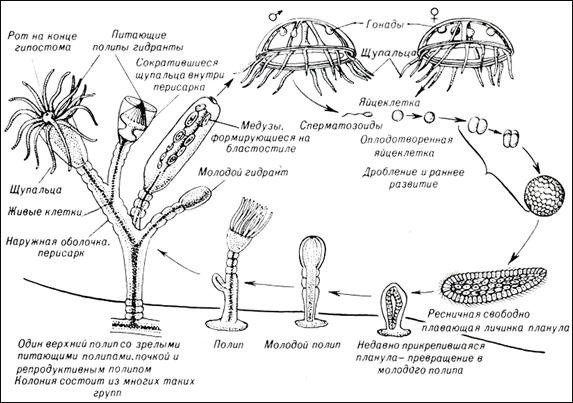 Рис. 29. Схема полового размножения полипов.
Рис. 29. Схема полового размножения полипов.
В настоящее время тип насчитывает 7 классов, из которых для палеонтологии важны два: Гидроидные (Hydrozoa) и Коралловые полипы (Anthozoa).
К классу Гидроидные (Hydrozoa)относятся наиболее примитивные стрекающие, чаще колониальные, реже одиночные с радиальной или четырехлучевой симметрией тела. Вокруг рта располагается 1-3 венчика щупалец, которые используются, в основном, для захвата пищи. Брюшная полость не разделена на перегородки. Скелет хитиноидный или карбонатный. Размножение бесполым путем (почкование) или половым (чередование полипоидной и медузоидной стадий). В отличие от других классов эти стадии носят названия гидрополипов и гидромедуз. В ископаемом состоянии гидроидные известны с кембрия. У некоторых представителей этого класса, например, у гидры пресноводной, развита только полипоидная стадия. Морские гидроидные основной жизненной формой имеют медузу. Для стратиграфии важна группа пока до конца неясного систематического происхождения — строматопораты.
Подкласс Строматопораты (Stromatoporata) традиционно относят к Hydrozoa. Однако в современных классификациях их часто сближают с губками и археоциатами. Это вымершие колониальные организмы; от них сохранились известковые слоистые скелеты разнообразной формы: от пластинчатой, корковидной до ветвистой (рис. 30). Слоистые скелеты представляют собой наслаивающиеся параллельно друг на друга пластины — ламины. В шлифах при увеличении видно, что ламины имеют разнообразное строение: пузырчатое, губчатое, петельчатое и т.д. Скелет строматопорат пронизан разнообразными пустотами. Самыми примечательными из них являются астроризы — каналы, не имеющие самостоятельных стенок, зато имеющие на верхней поверхности ламин вид звездчатых пустот, обычно с длинными горизонтальными разветвлениями (рис. 31). Они сходны со звездчатыми каналами некоторых губок, но также напоминают циклосистемы гидрокораллов. Отсюда дискуссионность их систематического положения: тип губки (Spongiata) или тип стрекающие (Cnidaria). Перпендикулярно ламинам располагаются зубчики, пересекающие ламины, шипики и стерженьки — столбики (рис. 32). Зубчики, шипики и стерженьки располагаются изолированно или создают различные комплексы в виде щеток, сеток, петель, трубок, колонн и пр. Строматопораты — донные колониальные животные, прикрепленные или свободнолежащие, нередко входившие в состав банок и рифов, образующие в ископаемом состоянии строматопоровые известняки. В палеозое (поздний ордовик, силур и девон) и мезозое (триас и юра) строматопораты активно участвовали в рифостроении. В отложениях каменноугольного и пермского периодов строматопораты практически не встречаются. Они существовали в нормально-морских условиях, но могли выдерживать и колебания солености, так как их находки известны вдоломитах. Форма палеозойских колоний разнообразная: пластинчатая, корковидная, полусферическая,


Рис. 30. Ископаемые колонии строматопорат


Рис. 31. Астроризы (звездчатые каналы) Рис. 32. Ламины с вертикальными
шипиками и зубчиками
дендроидная. Для мезо-кайнозойских строматопорат характерны сферические колонии.
Класс Коралловые полипы (Anthozoa) по форме тела и яркой окраске напоминают цветы, anthos — цветок. Это исключительно морские книдарии, требовательные к нормально морским условиям среды. Малейшее изменение температуры солености и газового режима грозит им гибелью. Коралловые полипы могут быть колониальные (рис. 33,а), или одиночные (рис. 33,б), и те и другие имеют только полипоидную стадию существования. В отличие от гидроидных коралловые полипы имеют более высоко организованное строение гастральной полости. Она разделена перегородками (мезентериями) на центральную пищеварительную полость и периферические камеры. Вокруг рта расположены щупальца (от 8 до нескольких десятков). Коралловые полипы (Anthozoa) могут питаться двумя способами. Первый основан на симбиозе с одноклеточными водорослями – зооксантеллами, обитающими в тканях животных. Водоросли получают от хозяина азотистые соединения, фосфаты и углекислоту, отдавая взамен полученные в результате фотосинтеза аминокислоты, углеводы и главный энергетический материал кораллов – липидные соединения. Второй (хищный) способ предусматривает захват из окружающей среды различных организмов.
Скелет кораллов – кораллит может быть роговой или карбонатный, (наружный или внутренний), некоторые виды вообще не имеют скелета. Кораллы обитатели морей всех глубин от литорали до ультраабиссали. Первые коралловые полипы известны из кембрия.


Рис. 33.а Колония коралловых полипов Рис. 33.б Одиночные полипы
В ископаемом состоянии скелеты кораллов образовывали органогенные постройки – биостромы и биогермы. Биостром – плоская органогенная постройка, из нарастающих друг на друга рифообразователей, не достигающая уровня воды. В ископаемом состоянии она выглядит как линза массивного известняка среди слоистых известняков. Разновидностью биострома является банка. Биогерм – изометричная органогенная постройка, резко возвышающаяся над морским дном, достигающая поверхности воды. В ископаемом состоянии это массивный известняк, «протыкающий» слоистые образования вокруг него. Риф (рис. 34) является разновидностью биогерма. По форме и расположению относительно берега рифы делятся на примыкающие (являются продолжением берега), барьерные (отделяют прибрежную лагуну от открытого моря) и атоллы (подводные горы, увенчанные рифовой постройкой) (рис. 34 а, б, с). Вокруг рифа (зона риффлет, рис. 34, В), накапливаются продукты его разрушения – глыбы, дресва, щебень, которые в процессе диагенеза превращаются в известковую брекчию. Высота рифов колеблется от нескольких метров до сотен метров. Древние коралловые рифы являются «ловушками» для нефти. Многие разрабатываемые нефтяные месторождения связаны с рифами.

Рис. 34. Типы и зональность рифов:
a - примыкающий, b - барьерный, c - атолл,
Различные зоны: A - лагуна, B - риффлет, C - склон рифа
В состав банок и рифов, как правило, входят скелеты не только кораллов, но и других организмов: моллюсков, брахиопод, губок. В строительстве рифов у кораллов есть помощники — определенные виды морских водорослей, скрепляющих коралловую массу.
Современный коралловый риф – это сложная и хрупкая экосистема, изобилие разных, притом чрезвычайно тесно соседствующих существ. Дополняют и усиливают эту экстраординарную интенсивность жизни сказочная красота форм и разнообразие красок. На коралловом рифе в одном месте и в одно время можно увидеть представителей практически всех структурных уровней развития организмов. Здесь представлен полный их спектр: от сине-зеленых морских водорослей и бактерий, многочисленных типов беспозвоночных животных до рыб, рептилий, птиц и млекопитающих.
Современные рифы заметно страдают от прожорливых врагов — морских звезд. Морская звезда «терновый венец» способна за день уничтожить тысячи полипов, оставляя хрупкие белые скелеты. Эти хищники нанесли немалый урон коралловым рифам у Японских берегов, а также Большому Барьерному рифу Австралии. В настоящее время выяснилось, что у морских звезд есть враг — гигантский моллюск тритон, который активно их уничтожает. Оказалось весьма эффективным в качестве охранных мероприятий выращивать тритонов в специальных питомниках и выпускать их на рифы.
Подкласс Табулятоморфы (Tabulatomorpha) объединяетвымерших палеозойских исключительно колониальных коралловых полипов, которые жили с позднего кембрия и вымерли в карбоне. Скелет их состоял из небольших (0,5-4 мм) трубчатых кораллитов, округлых или многоугольных в сечении (рис. 35; а, б).
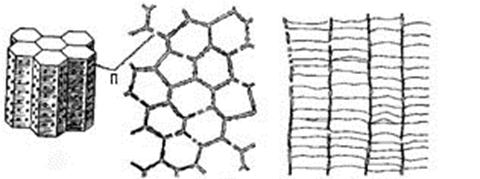
А б в
Рис. 35. Строение кораллитов табулятоморф:
а – общий вид, б – поперечное сечение,
в – продольное сечение с днищами, п – поры
Внутренние элементы кораллита – горизонтальные или воронковидные днища и радиальные септальные шипики (рис. 35, в; 37, 3а). Кораллиты были соединены друг с другом порами или соединительными трубочками; у некоторых видов связь между кораллитами отсутствовала. Колонии массивные, цепочечные (рис. 37, 1,2,3) кустистые, или стелющиеся.
Скелет по составу карбонатный. Подкласс подразделяется на отряды: аулопориды, хализитиды, фавозитиды, михелениды, сирингопориды и другие. От аулопорид возникли ругозы. Табулятоморфы обитали в мелководной зоне морей и входили в состав рифов. Имеют значение для стратиграфии палеозойских отложений.
Подкласс Ругозы (Rugosa)или четырехлучевые кораллы – одиночные (рис. 38, 1) или колониальные бентосные прикрепленные организмы. Колонии массивные, ветвистые (рис. 38; 3, 4). Произошли от табулят в ордовике, расцвет подкласса приходится на силур-девон, окончательно вымерли в пермском периоде. Имеют карбонатный скелет. На начальной стадии развития в кораллите выделяются 4 вертикальные перегородки – септы, разделяющие полость коралла на 4 части. По мере роста появляются другие большие септы, а между ними развиваются малые – септы второго порядка (рис. 38; 1, 2а). В карбоне у некоторых ругоз в центре кораллита развился столбик (рис. 38, 1). Кроме вертикальных перегородок в кораллитах ругоз имеются еще и горизонтальные элементы – днища и пузыреобразные пластинки диссепименты (рис. 38, 2б). Ругозы имеют значение для стратиграфии большей части палеозойских отложений, и являются основными рифообразователями палеозоя. В карбоне они дали начало шестилучевым кораллам.
 Подкласс шестилучевые кораллы (Hexacorallia) – ископаемые и ныне живущие, морские колониальные и одиночные организмы (рис. 33, 34), ведущие бентосный прикрепленный образ жизни. Скелет карбонатный, разделен септами на 6 частей. Имеют шестилучевую симметрию тела, количество щупалец кратное шести и септы трех порядков (рис. 36). Гексакораллы произошли от ругоз в каменноугольном периоде. В настоящее время они составляют основу современных рифов.
Подкласс шестилучевые кораллы (Hexacorallia) – ископаемые и ныне живущие, морские колониальные и одиночные организмы (рис. 33, 34), ведущие бентосный прикрепленный образ жизни. Скелет карбонатный, разделен септами на 6 частей. Имеют шестилучевую симметрию тела, количество щупалец кратное шести и септы трех порядков (рис. 36). Гексакораллы произошли от ругоз в каменноугольном периоде. В настоящее время они составляют основу современных рифов.
Рис. 36. Ископаемый гексакоралл
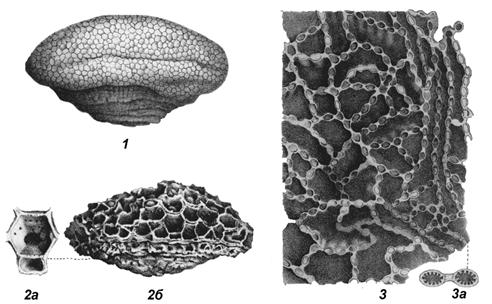
Рис. 37. Подкласс Табулятоморфы:
1 – общий вид массивной колонии фавозитид,2 а – ячейка отдельного кораллита (видны поры), 2 б – общий вид массивной колонии михеленид, 3 – цепочечная колония хализитид (вид сверху), 3 а – увеличенные ячейки хализитид (поперечное сечение) с септальными шипиками

 4
4
Рис. 38. Подкласс Ругозы:
1 – общий вид скелета одиночного коралла, 2 а - поперечное сечение одиночного коралла (видны септы), 2 б – продольное сечение одиночного коралла (видны днища и пузырчатая ткань), 3 – общий вид массивной колонии, 4 – общий вид ветвистой колонии
 Подкласс Восьмилучевые кораллы (Octocorallia) или альционарии — подкласс морских преимущественно колониальных организмов, с восемью щупальцами, насчитывающих около 2800 видов, которые разделяются на три отряда: альционарии (мягкие кораллы), роговые кораллы (в том числе благородный коралл, используемый как поделочный материал) и морские перья (рис. 39). Восьмилучевые кораллы распространены в основном в тропических морях, однако некоторые виды морских перьев населяют арктические воды.
Подкласс Восьмилучевые кораллы (Octocorallia) или альционарии — подкласс морских преимущественно колониальных организмов, с восемью щупальцами, насчитывающих около 2800 видов, которые разделяются на три отряда: альционарии (мягкие кораллы), роговые кораллы (в том числе благородный коралл, используемый как поделочный материал) и морские перья (рис. 39). Восьмилучевые кораллы распространены в основном в тропических морях, однако некоторые виды морских перьев населяют арктические воды.
Рис. 39. Современный восьмилучевой коралл
Отдельные виды октокораллов могут существовать на больших глубинах. Обитающие на рифах представители Octocorallia обычно более терпимы к экстремальным значениям факторов среды, чем Hexacorallia, и заселяют участки недоступные для них. Скелет их карбонатный, развивается в мезоглее. Многие восьмилучевые кораллы используют химические средства защиты, отравляя или отпугивая с их помощью потенциальных хищников. Это отличает их от шестилучевых, которые применяют для этой цели стрекательные клетки.
Раздел Двусторонне-симметричные (Bilateria)
Двусторонне-симметричные организмы характеризуются способностью к активному перемещению. Они подразделяются на первично- и вторичноротых. Первичноротые получили название по основному признаку: ротовое отверстие взрослой особи находится там же, где и у личинки.
Подраздел Первичноротые (Protostomia)
К первичноротым относятся приапулиды (низшие черви), высшие (кольчатые) черви, членистоногие, моллюски, а также, возможно, мшанки и брахиоподы (Приложения 5, 6). Для палеонтологии важны и будут здесь рассмотрены высшие (кольчатые) черви (аннелиды).
Тип Кольчатые черви (Annelida),обитающие в водной и наземной среде, имеют пищеварительную, кровеносную, половую, нервную, мышечную и иногда дыхательную системы. Водные аннелиды обычно ведут неподвижный образ жизни, строя трубки из хитинового и известкового материала. Трубка может быть цилиндрической, прямой, изогнутой (рис. 40, 2, 3). Наружное отверстие трубки может закрываться крышечкой (рис. 40, 4). В результате полового размножения возникает плавающая личинка, которая потом садится на дно и дает начало кольчатым червям (рис. 40, 1). При массовом скоплении рода Serpula и близких ему родов образуются карбонатные породы – серпулиты. Наиболее древние аннелиды, вероятно, появились в венде. В вендских отложениях часто встречаются следы ползания, проедания грунта и зарывания в него, образованные различными организмами, в том числе, аннелидами. Достоверно доказанные аннелиды известны с кембрия. В современных морях аннелиды живут на различных глубинах, вплоть до ультраабиссали. За счет эволюции аннелид произошли членистоногие и моллюски.

Рис. 40. Кольчатые черви (Annelida): 1 – личинка современных аннелид,
2 – отпечаток кольчатого червя на сланце, 3 - общий вид колонии современных аннелид, 4 - крышечки раковин вымерших палеогеновых аннелид
Представители типа Членистоногие (Arthropoda), характеризуются сегментарным строением тела, и в настоящее время населяют все среды обитания органического мира Земли. В современной фауне они представлены 3 млн. видов, из которых около 1 млн. составляют насекомые. Тело артропод защищено хитиновым скелетом (панцирем), иногда пропитанным углекислым кальцием или фосфатом кальция. Рост особи осуществлялся в промежутках между обновлениями панциря (линьками). По строению головного отдела и органов дыхания членистоногие делятся на 4 подтипа: Трилобитообразные, Ракообразные (Жабродышащие), Хелицеровые и Трахейные. Древнейшие достоверно доказанные артроподы известны с венда.
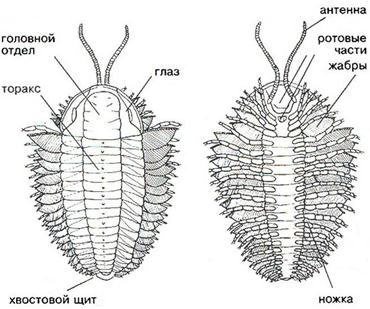
В подтипе Трилобитообразных (Trilobitomorpha) наибольшее значение для стратиграфии имеет класс Трилобиты(Trilobita) – вымершие морские животные (подвижный бентос), широко распространенные в отложениях нижнего и среднего палеозоя (более 10000 видов, принадлежащих к 2000 родов). Фактически трилобиты являются прародителями всех современных членистоногих. Они являлись первым подвижным бентосом на Земле: ползали по дну, зарывались в ил; некоторые плавали в придонных частях бассейнов. В ископаемом состоянии обычно встречаются фрагменты панциря и сравнительно редко – остатки конечностей, глаз, пищеварительного тракта. Скелет, в виде панциря, защищал лишь спинную часть тела трилобитов, а брюшная прикрывалась только кожистым покровом (рис.41, а,б). Скелет разделялся на сегменты: продольными бороздками обособлялась осевая часть тела и две боковые, а поперечными – головной и хвостовой щиты, туловищный отдел. В процессе эволюции у трилобитов выработалась способность сворачиваться, прижимая головной щит к хвостовому, в результате чего уязвимое брюшко оказывалось защищенным от хищников.
Головной щит, образованный 5-7 слившимися сегментами, подразделяется на осевую часть (глабель) и боковые (щёки), краевые части которых были подвижными. На щите располагались простые или фасеточные глаза. Туловище состояло из сегментов, число которых сильно варьировало (от 2 до 44). Каждый сегмент был снабжен парой двуветвистых конечностей, служивших для плавания или ползания по дну; их наружные ветви выполняли функцию жабр. Хвостовой щит, также как и головной, был образован сросшимися сегментами (от 1 до 30). По количеству сегментов и размерам трилобиты подразделяются на два подкласса: Малочленистые (Миомеры) и Многочленистые (Полимеры).
К миомерам относят мелких трилобитов (до 2 см), состоящих из 2-3 сегментов. Глаза обычно отсутствовали. Время существования – €-O. Полимеры характеризовались значительными размерами (1-70 см), а число сегментов у них было более 5. Они делятся на 7 отрядов. В процессе эволюции полимеров уменьшалась сегментация головного и хвостового щитов, сблизились их размеры (первоначально головной щит был значительно больше хвостового). Полимеры были распространены в течение всего палеозоя (рис.41, в), но уже в каменноугольных отложениях встречаются редко.
Класс Ракообразные(Crustacea) являются единственным классом подтипа, имеющего то же название – Ракообразные. Это – водные животные, тело которых подразделяется на голову, грудь и брюшко. На голове располагаются две пара усиков, играющих роль антенн, челюсти и глаза. К грудному отделу принадлежат конечности, служащие для движения, дыхания и захвата пищи. Для стратиграфии наибольший интерес представляют две группы примитивных ракообразных: Остракоды и Филлоподы.
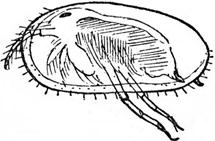
Подкласс Остракоды – ракушковые рачки, населяющие морские, лагунные и пресноводные водоемы, подразделяются на 11 палеозойских и 5 мезо-кайнозойских отрядов. Размеры остракод варьируют в пределах 0,2-7 мм, но представители некоторых групп достигают 1-3 см. Тело особи заключено в двустворчатую раковину, построенную из органического вещества, пропитанного углекислым кальцием, (рис. 42, а, б). Рост тела происходил в промежутках между линьками, поэтому знаков роста на раковинах нет. Остракоды известны с кембрийского периода; пресноводные группы впервые появились в каменноугольных отложениях.
Из подтипа Хелицеровых класс Меростомовые (Merostomata) имеет наибольшую стратиграфическую значимость. Он разделяется на подклассы Мечехвостые и Эвриптероидеи (ракоскорпионы) (рис 43). Размеры эвриптероидей обычно составляли 10-20 см, но некоторые роды достигали 2 м в длину. С ордовика до перми они населяли солоноватые и пресные водоемы, являясь хищниками. Считается, что именно эти членистоногие стали первыми обитателями суши.
Класс Насекомых(Insecta) представлен наибольшим количеством видов (около 1 млн.). Тело насекомого длиной от 0,2 мм до 33 см состоит из головы, груди и брюшка, покрытых хитиновой оболочкой. Голова, образованная слиянием 4 сегментов, снабжена глазами, парой конечностей, преобразованных в антенны и тремя парами конечностей,
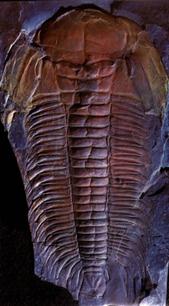
А б в
Рис. 41. Класс Трилобиты (Trilobita), подкласс Полимеры:
а – спинной отдел трилобита, защищенный панцирем, б – брюшной отдел трилобита, в - отпечаток кембрийского трилобита рода Paradoxides

А
Б
Рис. 42. Подкласс Остракоды (Ostracoda): а –схема внутреннего строения остракод (видны конечности, глаз и жабры), б – современная остракода (увелич.)

Рис. 43. Отпечаток ракоскорпиона Eurypterus remipes на песчанике
превращенных в челюсти. Грудь состоит из трех сегментов, каждый из которых снабжен парой ног. Кроме того, второй и третий сегменты груди несут крылья. Брюшко состоит из 5-11 сегментов, не имеющих конечностей. Насекомые появились в девоне, а в настоящее время переживают эпоху расцвета. Важны для стратиграфии, начиная с позднего палеозоя. Насекомые играли важную роль в эволюции растительности, особенно цветковых. Из беспозвоночных это единственная группа, освоившая полет в воздушном океане.
Тип Моллюски (Molluska)подразделяется на 10 классов, из которых наибольшее значение для стратиграфии и фациального анализа имеют Двустворчатые, Брюхоногие и Головоногие моллюски, а также Тентакулиты (Приложения 6 и 7). Тело моллюска, как правило, имеет двусторонне-симметричное строение, подразделяясь на голову, туловище и ногу. Обычно оно защищено известковой раковиной и только у некоторых групп она отсутствует. Характерной особенностью моллюсков является обитание в разных средах, что обусловило высокую изменчивость морфологических признаков. Особым разнообразием характеризуются нога и раковины. По видовому разнообразию (150 тыс. видов) моллюски стоят непосредственно за членистоногими. Моллюски известны, начиная с кембрия.
Представители класса Двустворчатые моллюски(Bivalvia) – морские или пресноводные животные, тело которых заключено в двустворчатую раковину, причем одна створка является зеркальным отражением другой (рис. 44, б). Мягкое тело не имеет обособленной головы, но есть мускульная нога для прикрепления. Прикрывающая тело мантия – особая складка кожи, выделяющая перламутр, вещество, которое является основой для жемчуга (рис. 44, г). Жемчуг – это органогенная конкреция, растущая внутри раковины вместе с моллюском.
Раковины двустворок имеют выступающую макушку и край с замочным аппаратом, скрепляющим их друг с другом. Сочленение створок осуществлялось с помощью связки и замка, состоящего из выступов (зубов) на одной створке и зубных ямок – на другой (рис. 44, а). Размеры раковин варьируют в широких пределах от долей миллиметра и почти до метра. Наиболее крупные характерны для приэкваториальных зон. Форма раковин разнообразна до причудливости: округлые, эллиптические, конические, рогообразные, шиповатые (рис. 44,) и т.д. Систематика современных и ископаемых двустворок строится по-разному: современные классифицируются по

А б


В г
Рис. 44. Двустворчатые моллюски: а – внутренняя часть створки с зубным аппаратом, б – раковина современного рода Cardium, в - наружная часть створки с шиповато-ребристой скульптурой, г - современная двустворка с жемчужиной
 строению мягкого тела, ископаемые – по строению замка. По этому принципу выделяется шесть отрядов, различающихся наличием или
строению мягкого тела, ископаемые – по строению замка. По этому принципу выделяется шесть отрядов, различающихся наличием или
| |
Из зон Мирового океана двустворки предпочитают сублитораль. Одни группы медленно передвигались по поверхности грунта, другие переходили к неподвижному образу жизни, прирастая к субстрату. Некоторые двустворки за счет ритмичного открывания и захлопывания створок освоили перемещение в придонном слое воды, другие зарывались в илистые грунты или сверлили камни или дерево. Нередко образовывали массовые скопления – банки. Некоторые двустворки (устрицы, гребешки) являются продуктами питания человека.
Двустворки появились в кембрии, но настоящего расцвета достигли только в мезозое и широко распространены до наших дней. Во второй половине мезозоя в теплых бассейнах появились массивные прикрепленные двустворки (рудисты), конвергентно сходные с ругозами. Они активно участвовали в рифостроении. Бивальвии имеют значение для фациального анализа, а также для стратиграфического расчленения мелового, палеогенового и неогенового периодов.
Класс Брюхоногие моллюски (Gastropoda) включает современные и вымершие виды (рис. 45 – 48) и является наиболее многочисленным классом типа моллюсков, освоившим разнообразные среды обитания (море, пресные водоемы, суша). Они характеризуются асимметричным телом с обособленной головой, на которой располагаются органы зрения, обоняния и осязания в виде антенн. Тело заключено в раковину, спирально закрученную в горизонтальной или вертикальной плоскости (рис. 45, 46). Большинство гастропод ведет подвижный образ жизни, передвигаясь с помощью ноги c присоской, помещенной на брюшной полости. В зависимости от строения органов дыхания выделяются подклассы Переднежаберные, Заднежаберные и Легочные.
Подкласс Переднежаберные характеризуется расположением жабер впереди сердца. Это – преимущественно морские организмы, обладающие спиральнозавитой (рис.47, а) или колпачковидной (рис. 47, б) раковиной. Имеется нога в виде подошвы или киля. Переднежаберные появились в кембрии и существуют до настоящего времени. По традиции выделяются отряды Археогастроподы, Мезогастроподы и Неогастроподы, хотя сейчас принято более дробное деление.
Подкласс Заднежаберные обладает одной жаброй, расположенной позади сердца. Раковина редуцирована и нередко отсутствует. Преимущественно морские организмы, населяющие все зоны Мирового океана. Некоторые группы освоили пресноводные водоемы. Заднежаберные известны с каменноугольного периода до настоящего времени. В качестве особого отряда Заднежаберных выделяются Крылоногие моллюски – Птероподы (P – ныне) – характеризующиеся мелкими раковинами (1-2 мм). Нога их изменена в два крыловидных плавника (рис.48). После смерти животных за счет скопления раковин образуются птероподовые илы.
Подкласс Легочные (Pulmonata), (рис. 45) существующие с каменноугольного периода, освоили субаэральную среду, в связи с чем, жабры у них заменились легкими. В составе подкласса имеются раковинные и безраковинные формы. Большинство легочных моллюсков являются растительноядными, некоторые – хищниками или паразитами. Гастроподы используются для стратиграфии кайнозойских отложений. В последние годы предпринимаются попытки использования для стратиграфии палеозойских и мезозойских представителей этого класса.


|
Раковина вертикально закрученная, с крышечкой
на задней части тела. Голова обособлена и
снабжена органами зрения, обоняния и осязания
А б
Рис. 47. Представители подкласса Переднежаберные:
а - хищные гастроподы рода Conus (палеоген-ныне),
б – различные виды современных гастропод рода Patella (мел-ныне)
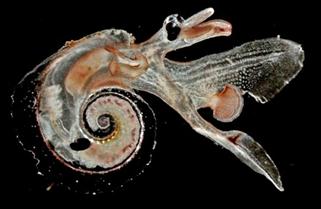
Рис. 48. Подкласс Заднежаберные: современные птероподы
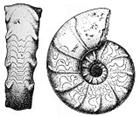
Класс Головоногие моллюски (Cephalopoda) является наиболее важным для стратиграфии фанерозоя классом типа Моллюски. У большинства цефалопод тело заключено в прямую или спирально закрученную многокамерную раковину. Строение межкамерной перегородки и её отпечаток на раковине – перегородочная линия – являются важными классификационными признаками. Эволюция цефалопод сопровождалась усложнением очертаний перегородочной линии. Тело располагается в последней камере, а остальные заполнены газом, благодаря чему увеличивается плавучесть раковины. Соединение камер обеспечивается сифонной трубкой (рис. 49, б). Частично заполняя камеры водой, особь могла регулировать свое положение в толще воды. Нога цефалопод преобразована в венчик щупалец и воронку, выбрасывающую воду, что обеспечивает реактивное движение особи. В последние годы несколько изменена классификация цефалопод. Они разделены на 7 подклассов, из которых два (наутилоидеи и колеоидеи) представлены в современной фауне, а остальные – вымерли. У шести подклассов тело заключено в раковину, а у седьмого (колеоидеи) – раковина находится внутри тела, выполняя роль внутреннего скелета.
Подкласс Наутилоидеи, известный в ископаемом состоянии с позднего кембрия, в современной фауне представлен единственным родом Nautilus (отряд Наутилиды). Тело наутилоидей было заключено в прямую или спирально закрученную раковину, межкамерная перегородка прямая (рис.49, а, б).


А б
Рис. 49. Род Nautilus: а – современный наутилус
б – ископаемый наутилус с прямой перегородочной линией
Подкласс Ортоцератоидеи (O –T) характеризовался прямой раковиной, достигавшей в длину 1-1,5 м. Межкамерная перегородка линзовидно-изогнутая, перегородочная линия прямая или почти прямая. Сифон субцентральный (рис. 50). К подклассу Эндоцератоидеи(O) принадлежали наиболее крупные цефалоподы, достигавшие в длину 3-4, а в отдельных случаях до 10 м. Поперечное сечение – круглое или овальное. На сифон, расположенный на брюшной стороне и занимающий краевое положение, могло проходиться до 1/3 поперечного сечения (рис. 50). Прямая перегородочная линия. Подкласс Бактритоидеи являлся небольшой по числу видов группой, обладавшей прямой раковиной. Узкий сифон занимал краевое положение. Подкласс рассматривается как возможный предок аммоноидей.
| |
| |

Рис. 50. Реконструкция ордовикского моря: 1 – плавающие ортоцератоидеи,
2 – охотящийся на трилобитов, представитель эндоцератоидей
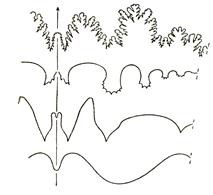
Подкласс Аммоноидеи, характеризующийся наибольшим видовым разнообразием, появился, по-видимому, в девонском периоде и вымер в конце мелового (рис. 53). Аммоноидеи со спирально закрученной раковиной существовали с девона до мела и фигурируют в качестве мономорфных; с прямой, коленообразно изогнутой или крючковатой раковиной – как гетероморфные. Они появились в позднем триасе, а вторично возникли в средней юре, но наиболее широкое распространение получили во второй половине мела. Жилая камера у разных представителей этого подкласса занимала от половины до 2 оборотов раковины. В ней отсутствуют межкамерные перегородки, а в остальных камерах они имеют сложноскладчатый характер. Перегородочная линия состоит из лопастей и сёдел. Изгибы перегородочной (лопастной) линии в зависимости от направления изгиба (к жилой камере или к заднему краю раковины) называются либо седлами, либо лопастями. У аммоноидей наблюдается 4 типа перегородочной линии: агониатитовая (рис. 52, а), гониатитовая (рис. 52, б), цератитовая (рис. 52, в) и аммонитовая (рис. 52, г). Агониатитовая линия характеризуется округлым характером нерасчлененных сёдел и лопастей. При гониатитовой линии сёдла и угловатые лопасти не расчленены. При цератитовой – расчленены все лопасти, при аммонитовой – расчленены и сёдла, и лопасти. Строение перегородочной линии, её усложнение во времени, а также положение сифона лежат в основе расчленения подкласса Аммоноидеи на восемь стратиграфически значимых отрядов.
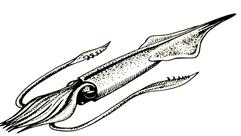
Подкласс Колеоидеи объединяет группы головоногих моллюсков, у которых раковина играла роль внутреннего скелета. Это – активно плавающие хищники (рис. 51, а). Древние колеоидеи объединены в надотряд Белемноидеи (C – K). В ископаемом состоянии обычно сохраняется основная часть внутреннего скелета – ростр – массивное, заостренное на конце цилиндрическое образование, состоящее из арагонита (рис. 51, б). В процессе онтогенеза форма ростра нередко менялась. В углублении расширенного конца ростра – альвеоле – располагались важнейшие части тела особи. Современные колеоидеи – надотряд Декабрахии (J – Q) – представлены осьминогами, каракатицами, кальмарами. К ним, в частности, принадлежат наиболее крупные представители беспозвоночных – гигантские кальмары, достигающие в длину 18 м. Представители третьего надотряда – Октобрахии (K – Q) не имеют скелета, в силу чего их стратиграфическая значимость невелика.

|
|

,
Рис.51. Надотряд белемноидеи: а –реконструкция древних форм
б – ростры ископаемых белемноидей (J-K
 г
г
В
Б
Рис. 52. Перегородочные линии
Аммоноидей
А

|
|

| |
D – P
D
|
Рис. 53. Эволюция во времени раковин цефалопод и их перегородочных линий
 Тентакулиты – группа неясного систематического положения. Это вымершие морские животные, обладавшие конической известковой раковиной размером 2-5 до 30-70 мм. Раковина состояла из жилой камеры или была разделена на несколько камер, не сообщавшихся между собой. Поверхность раковины иногда была гладкой, чаще характеризовалась наличием ребер и пережимов (рис. 54). Тентакулиты появились в силуре и широко распространились в начале девона. Некоторые приспособились к планктонному образу жизни, другие – населяли придонные слои бассейна или зарывались в ил. Вымирание тентакулитов произошло в конце девона. Большая часть исследователей рассматривает тентакулитов в качестве класса типа Моллюски.
Тентакулиты – группа неясного систематического положения. Это вымершие морские животные, обладавшие конической известковой раковиной размером 2-5 до 30-70 мм. Раковина состояла из жилой камеры или была разделена на несколько камер, не сообщавшихся между собой. Поверхность раковины иногда была гладкой, чаще характеризовалась наличием ребер и пережимов (рис. 54). Тентакулиты появились в силуре и широко распространились в начале девона. Некоторые приспособились к планктонному образу жизни, другие – населяли придонные слои бассейна или зарывались в ил. Вымирание тентакулитов произошло в конце девона. Большая часть исследователей рассматривает тентакулитов в качестве класса типа Моллюски.
Рис. 54. Раковина тентакулита
Тип Брахиоподы (Brachiopoda) – одиночные донные двусторонне-симметричные морские животные, тело которых заключено в двустворчатую раковину. Размеры раковины 0,1-30,0 см в длину, наиболее часто – 3-5 см. Раковина состоит из неравных по величине створок, поперек которых проходит плоскость симметрии (рис. 56, 1, 2). Прикрепление раковины к субстрату осуществляется с помощью ножки, расположенной в задней части тела. У многих брахиопод во взрослом состоянии ножка редуцируется. По типу обмена веществ брахиоподы являются фильтраторами. Приток воды обеспечивается кожными выростами – лофофорами, функционально аналогичными щупальцам мшанок (рис. 57). Лофофоры поддерживаются выростами раковины, образующими ручной аппарат, строение которого является важным систематическим признаком (рис. 56, 3; 57). Брахиоподы появились в кембрии, получили широкое распространение в течение палеозоя и в значительной мере вымерли в конце пермского периода. Классификационная принадлежность брахиопод, как и описанного ниже типа Мшанки, остается дискуссионной. В данной работе брахиоподы и мшанки (Приложение 4) условно рассматриваются в качестве особых типов без указаний о принадлежности последних к первично- или вторичноротым.
Класс Беззамковые брахиоподы (кембрий – ныне) подразделяется на отряды (Лингулиды, Краниды и т.д.). У первого отряда раковина хитиново-фосфатная и имеется ножка для зарывания в грунт (рис. 55), у второго – раковина известковая, а ножка отсутствует. Для стратиграфии кембрия и ордовика наибольшее значение имеет отряд Лингулида,

Рис. 55. Современные раковины лингулид
представителями которого являются роды Lingula (ордовик – ныне) и Obolus (средний кембрий – ранний ордовик).
У класса Замковых брахиопод соединение створок раковины осуществлялось с помощью замка – зубов на брюшной створке и зубных ямок – на спинной (рис. 56, 3). Брюшная створка отличалась более загнутой макушкой и обычно большими размерами. Известковая раковина имела разнообразную форму; поверхность её либо несла следы роста, либо была скульптурирована. Иногда створки раковины были изогнуты таким образом, что на брюшной створке образовывалась борозда – синус, а на спинной возвышение – седло. Наличие седла и синуса способствовало разделению входящих и выходящих потоков воды.
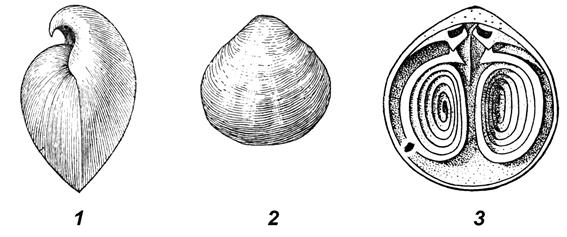
Рис. 56. Раковины брахиопод:1 – вид сбоку обеих створок, 2 – наружная часть спинной створки,3 – внутренняя часть брюшной створки с ручным аппаратом
По особенностям строения створок, характеру отверстия для ножки, а, главное, по строению ручного аппарата замковые брахиоподы делятся на подклассы Ортаты, Строфоменаты, Спирифераты, Теребратулаты.
Подклассы делятся на отряды. Для стратиграфии наибольшее значение имели отряды Продуктиды (девон – пермь), Спирифериды (ордовик – нижняя юра), Теребратулиды (девон – ныне). Продуктиды характеризовались вогнуто-выпуклой или плосковыпуклой раковиной, снабженной шипами. Отверстие для ножки и зубы обычно отсутствуют. Ручной аппарат не развит. Раковины достигают 30 см. Спирифериды имели развитый ручной и зубной аппарат, ребристую поверхность раковины, четко выраженные синус и седло, треугольное отверстие для ножки (дельтирий). Теребратулиды имели овальную двояковыпуклую раковину, хорошо выраженный ручной аппарат, зубы, округлое отверстие для ножки (форамен). Большинство замковых брахиопод прикреплялось ножкой к дну (рис. 57) или лежало на грунте, частично погружаясь в него. Глубокому погружению препятствовало наличие шипов.
|
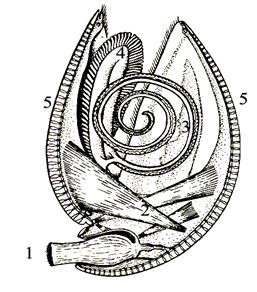
Появившись в кембрии, замковые брахиоподы получили широкое распространение в течение последующих периодов палеозоя, населяя зону мелководья. В течение мезо-кайнозоя эту зону освоили двустворчатые моллюски, постепенно вытеснившие брахиопод. До настоящего времени дожили отряды Ринхонеллид и Теребратулид. Среди ныне живущих брахиопод имеются глубоководные виды. Нередко в отложениях палеозоя наблюдались скопления замковых брахиопод – брахиоподовые известняки.
Тип Мшанки (Bryozoa)– колониальные животные, населяющие нормально-морские, солоноватые и пресные водоемы до глубины 400-500 м. Они являются прикрепленными организмами, за исключением нескольких форм ползающих по дну. Размеры колоний от 1 мм до нескольких десятков сантиметров. Форма колоний мшанок разнообразна: ветвистые, массивные, пластинчатые, сетчатые, инкрустирующие. Нередко форма колоний является индикатором среды обитания. Между элементами колонии – зооидами наблюдается определенная степень специализации. Автозооиды выполняют функции питания, а иногда и полового размножения. Вторичная полость тела – целом – делится тонкой перегородкой на туловищный и щупальцевый
отделы. Ротовое отверстие окружено венчиком щупалец, обеспечивающих приток воды, поскольку мшанки по способу питания являются фильтраторами. Гетерозооиды выполняют функции защиты, укрепления и размножения. Элементы скелета колоний морских мшанок – авто- и гетерозооеции имеют известковый состав. Морские мшанки являются рифостроящими организмами (рис. 59).
|


Рис. 58. Класс Gymnolaemata,
род Archimedes (C-P1)
Тип включает около 4000 современных и приблизительно столько же ископаемых видов (рис. 58), объединяемых в два класса: Гимнолематы (Голоротые) и Филактолематы (Покрыторотые) (Приложение 4).
К классуГимнолематы(рис. 60) относятся морские ныне живущие и все вымершие мшанки. Щупальца располагаются вокруг рта по окружности, а ротовое отверстие не прикрыто. Известны с ордовикского периода (аренигский век) по настоящее время. По строению автозооециев, наличию определенных гетерозооециев, типам почкования автозооециев и некоторым другим признакам большинством исследователей класс Гимнолематы подразделяется на два подкласса и шесть отрядов.
В состав класса Филактолематывходят пресноводные мшанки. Из-за отсутствия минерального скелета они в ископаемом состоянии не сохраняются. Некоторые исследователи относят мшанки к особой группе целомических животных.
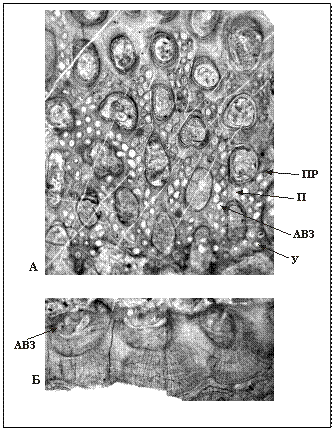
Рис. 60. Класс Gymnolaemata, отряд Fenestellida (О2 – P).
А, Б – сетчатая колония рода Septopora (D1 – P2), сильно увел.:
А – тангенциальное сечение,
Б – поперечное сечение;
ПР – прутья, П – перекладины, АВЗ – камеры автозооециев,
У – устья автозооециев.
Подраздел Вторичноротые (Deuterostomia)
Из вторичноротых животных наибольшее значение для стратиграфии имеют иглокожие, полухордовые, конодонты и хордовые (Приложение 7).
Тип Иглокожие (Echinodermata) – морские животные, приспособившиеся к бентосному или планктонному образу жизни. Их характерной особенностью является пятилучевая симметрия и наличие амбулакральной системы, состоящей из заполненных водой каналов, обеспечивающих движение и обмен веществ. Появление иглокожих датируется вендом, но они широко представлены и в современной фауне. Стратиграфическое значение имеют классы цистоидей и криноидей.
Класс Морские пузыри (Cystoidea) – наиболее примитивный класс кринозой. Пятилучевая симметрия не всегда выражена, амбулакральная система находится в зачаточном состоянии. Цистоидеи – морские бентосные животные, тело которых заключено в округлую чашечку. Они появились в среднем кембрии, получили широкое распространение в ордовике, а к началу позднего девона – вымерли. Имеют значение для стратиграфии ордовикских отложений.

Рис. 61. Тип Иглокожие.
1- морские пузыри; 2 а,б – морские лилии;
3 - морские ежи: а – общий вид морского ежа, б – вид ежа снизу (видны рот и анус)
Представители классаМорских лилий (Crinoidea) имеют тело, состоящее из чашечки, пяти рук и стебля, для прикрепления к грунту или каким-либо объектам. Чашечка округлой или полушаровидной формы состояла из нескольких поясов, сложенных пятью табличками каждый. Руки, отходящие от табличек, иногда ветвятся, что увеличивает площадь сбора пищи. Количество рук кратно пяти. Стебель морских лилий имел членистое строение (рис. 62). Членики рук и стебля обычно сохраняются в ископаемом состоянии отдельно от чашечки, образуя криноидные известняки. Форма и размеры члеников весьма разнообразны и являются предметом специальных исследований. Морские лилии появились в ордовике, как обитатели бентоса и достигли разнообразия в морях позднего палеозоя. Ко дну они прикреплялись стержнем. В мезозое часть криноидей утратила стебель и перешла к планктонному образу жизни.
Среди современных представителей этого класса господствуют бесстебельные формы. Стебельчатые группы криноидей в современных морях населяют лишь глубоководные зоны. Некоторые виды известны на глубинах до 10000 м. Длина их стебля не превышает 1 м. В ископаемом состоянии известны виды с длиной стебля до 20 м. Таким
образом, ископаемые криноидеи вместе с современными гигантскими кальмарами принадлежат к наиболее крупным беспозвоночным.
 КлассМорские ежи (Echinoidea) – подвижные морские животные с округлой или дисковидной формой тела и четко выраженной пятилучевой симметрией, иногда усложненной наличием двусторонней симметрии (O – ныне). Передвижение по дну осуществляется иглами, приводимыми в движение с помощью амбулакральной системы (рис. 63). Класс характеризуется наличием панциря и пятичелюстного ротового аппарата. Древнейшие представители этого класса, вероятно, произошли от прикрепленных животных. Изменение образа жизни обусловило появление элементов двусторонней симметрии, особенно характерной для мезозойских неправильных ежей. Современные морские ежи – подвижные хищники или илоеды, обитающие в теплых морях.
КлассМорские ежи (Echinoidea) – подвижные морские животные с округлой или дисковидной формой тела и четко выраженной пятилучевой симметрией, иногда усложненной наличием двусторонней симметрии (O – ныне). Передвижение по дну осуществляется иглами, приводимыми в движение с помощью амбулакральной системы (рис. 63). Класс характеризуется наличием панциря и пятичелюстного ротового аппарата. Древнейшие представители этого класса, вероятно, произошли от прикрепленных животных. Изменение образа жизни обусловило появление элементов двусторонней симметрии, особенно характерной для мезозойских неправильных ежей. Современные морские ежи – подвижные хищники или илоеды, обитающие в теплых морях.

| |||
| |||
К типу Полухордовых (Hemichordata) принадлежат три класса, из которых наибольшее значение имеет класс Граптолиты.
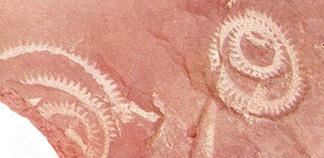
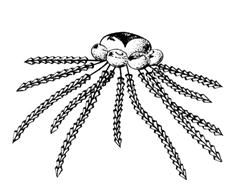
Класс Граптолиты (Graptolithina) – вымершие колониальные животные, имевшие скелет близкий по составу к хитиновому. Колонии граптолитов (рабдосомы) весьма разнообразные по форме состояли из большого количества ячеек (от одного до четырех рядов), прикрепленных к ветвям, также имевшим различную форму. Граптолиты либо прикреплялись к дну, либо к плавающим объектам (псевдоплактон). Некоторые формы, имевшие воздушный пузырь, принадлежали к типичному планктону (рис. 64, а). Остатки граптолитов обычно содержатся в глинистых сланцах, реже – в песчаниках и известняках (рис. 64,б). При изобилии граптолитов выделяется особая разновидность пород – граптолитовые сланцы.
Граптолиты подразделяются на подклассы Стереостолонаты и Граптолоидеи (Приложение 7). Из стереостолонат наибольшее значение имеет отряд Дендроидные граптолиты (C3–C), образовывавшие уплощенные или воронкообразные колонии. По образу жизни дендроидные граптолиты принадлежали к бентосу, псевдопланктону или планктону. Граптолоидеи, имевшие прямые и спиральнозавитые колонии, характеризовались планктонным или псевдопланктонным образом жизни (рис. 64, б). Граптолоидеи были широко распространены с ордовика до раннего девона.
 а б
а б
Рис. 64. а – реконструкция колонии граптолитов с плавательным пузырем,
Б – отпечаток на песчанике отдельных нитей
Древнейшие конодонты известны со среднего кембрия. Расцвет простых форм приходится на ордовик. Сложные и платформенные конодонты достигают… Вопросы для самоконтроля:Рис. 67. Живое ископаемое – кистеперая рыба латимерия
ПодклассДвоякодышащие рыбы имеют некоторые общие черты с кистеперыми, но отличаются от них отсутствием теменного глаза и строением зубов (рис. 68, 2). Они могли некоторое время находиться на суше и дышать воздухом. Длина тела до 2 м. Населяют только пресноводные водоемы, питаясь, в основном, растительностью. Раньше их считали предками земноводных, но в настоящее время это вызывает сомнение.
ПодклассЛучеперые рыбы – наиболее распространенный в настоящее время подкласс, представители которого достигают 9 м в длину. Они характеризуются развитием чешуи разных типов, наличием в плавниках лучевидных шипов, отсутствием непарного глаза. Среди ископаемых лучеперых широкое распространение получили хейролеписы (рис.68, 1) и палеониски (D3 – K1), обеспечившие широкое распространение подкласса в течение каменноугольного и пермского периодов. В кайнозое началась вторая эпоха расцвета лучеперых.

Рис. 68. Древние рыбы: 1 – лучеперая рыба хейролепис, 2 – двоякодышащая, диптер, 3 – пластинокожая рыба, 4 – антиархи
К Надклассу Четвероногих (Tetrapoda) принадлежат пять классов: Земноводные (Амфибии), Парарептилии, Пресмыкающиеся (Рептилии), Птицы и Млекопитающие (Приложение 7). По уровню организации четвероногие подразделяются на низших (амфибии) и высших, включающих остальные четыре класса. Для тетрапод характерно наличие легких, усложненное строение внутреннего скелета и зубной системы, наличие внешнего скелета (чешуя, панцири, когти, клюв, рога, волосы). Они в большинстве случаев являются наземными животными, иногда – первично- и вторичноводными.
Класс Земноводные (Amphibia) имеют общие черты, как с рыбами, так и с представителями высших тетрапод (D3 – ныне). С рыбами их сближает наличие жабр (на начальной стадии развития, реже – в течение всей жизни), размножение икрой, двояковогнутые позвонки и некоторые другие черты строения. Общими признаками с высшими тетраподами являются наличие четырех конечностей и связанных с этим плечевого и тазового поясов, положение теменного «глаза», а также наличие шейного и крестцового позвонков.
Позвонки земноводных по особенностям строения подразделяются на четыре типа. У древних и некоторых современных земноводных присутствует наружный скелет в виде щитков, пластин и покровных частей черепа. Облик земноводных различен: одни напоминают рыб, другие – лягушек, третьи имеют змеевидный облик (червяги). Длина тела у современных земноводных достигает 1,8 м, у ископаемых – 5 м. Классификация земноводных, основанная на строении черепа, осевого скелета и конечностей, остается дискуссионной. Некоторые группы (стегоцефалы) оказались сборными, имеющими разное происхождение. В частности, многие исследователи исключают из земноводных группу сеймуриаморфов, широко распространенных в пермском периоде. Стегоцефалы (панцирноголовые) (D3 – K1) населяли пресноводные внутри-континентальные водоемы (рис.69).Этовымершая группа земноводных, вышедших одними из первых на сушу в конце девона. Предками стегоцефалов являлись кистепёрые рыбы, скелет которых имеет много сходных черт с их скелетом. В настоящее время бо́льшую часть стегоцефалов включают в надотряд Labyrinthodontia (лабиринтозубых).

Рис. 69. Самое древнее земноводное – акантостега, считается предком всех четвероногих животных
Современные земноводные, подразделяются на бесхвостых
(лягушки, жабы, квакши), хвостатых (тритоны, саламандры) и безногих (червяги).
В пермском периоде появились парарептилии сочетающие в себе признаки земноводных и рептилий. К ним относят некоторых сеймуриаморфов, частично котилозавров и древних черепах. Это были морские и пресноводные животные. Плотоядные (главным образом, рыбоядные) сеймуриаморфы (P) жили в воде и дышали жабрами. Растительноядные парейазавры (P), имевшие головной панцирь, достигали 4 м в длину. У черепах (T – ныне) тело заключено в панцирь, достигавший в поперечнике 2 м. Зубной аппарат у них отсутствует.
Класс Пресмыкающиеся (Reptilia) (C – ныне) обладают способностью размножаться яйцами, что в совокупности с легочным дыханием и чешуей на коже обеспечило их независимость от водной среды. Таким образом, был расширен ареал обитания и характер пищи. Внешний вид, образ жизни и размеры рептилий резко различны. Наиболее древние группы обнаруживают сходство с земноводными и парарептилиями, более молодые – с птицами и млекопитающими вплоть до появления теплокровности, вторичного нёба, четырехкамерного сердца, перьев и шерсти. Столь же различен был их облик: одни напоминали рыб и ластоногих, другие – птиц, третьи – млекопитающих. Длина водных и сухопутных рептилий достигала 35-40 м, а размах крыльев летающих – 11-15 м. Наиболее известными ископаемыми рептилиями являются динозавры, ихтиозавры и птерозавры, а среди современных – крокодилы, черепахи, змеи, ящерицы (Приложение 9).
Намечается комплекс признаков специфичных для элементов скелета рептилий. В отличие от парарептилий череп рептилий имеет височные окна. От земноводных рептилии отличаются наличием грудной клетки, образованной сращением ребер с грудиной. От птиц и млекопитающих их отличает, в первую очередь, строение и количество позвонков. Выделяется четыре типа строения черепа. По строению черепа и конечностей, в сочетании с другими признаками, выделяется пять подклассов рептилий: ихтиозавры, синаптозавры, чешуйчатые, архозавры и синапсиды. Систематика рептилий остается дискуссионной. Большинство современных рептилий ведет наземный образ жизни. Часть ископаемых рептилий во время максимального распространения класса осваивала водную и воздушную среды обитания.
Подкласс Ихтиозавры (T3 – K, расцвет в J1) – наиболее примитивные морские рептилии, внешне сходные с рыбами и дельфинами. В последние годы для обозначения этого подкласса стал использоваться термин «ихтиоптеригии» (рыбоплавниковые). Характерными особенностями подкласса являются форма черепа, снабженного отверстием для «теменного глаза» и двояковогнутая форма позвонков. Ихтиозавры являлись главными хищниками сублиторали, охотясь на рыб, аммонитов и белемнитов (рис. 70). Длина их достигала 15 м. Систематика ихтиозавров остается дискуссионной, поскольку архаичные черты совмещались у них с прогрессивными (они были живородящими). Иногда к ихтиозаврам относят мезозавров – пресноводных крокодилообразных рептилий длиной до 1 м (C3 – P1).
Характерными чертами подкласса Синаптозавры являлись форма черепа, особенности строения позвонков, ластообразная форма конечностей. Синаптозавры были морскими хищниками, охотившимися на рыб и головоногих моллюсков. Подкласс подразделяется на отряды Завроптеригии и Плакодонты.

Рис. 70. Ихтиозавры
Охотятся на аммонитов
В состав завроптеригий входят группы Плезиозавры и Плиозавры. Плезиозавры (T2 – K) обладали длинной шеей, маленькой головой, веретенообразным телом,… Подкласс Лепидозавры («чешуйчатые ящеры») характеризовались особой формой… Подкласс Архозавры – наиболее разнообразный подкласс рептилий. Они различаются по облику, размерам, строению скелета и…Веерохвостые птицы (K – ныне) имеют короткий хвостовой отдел позвоночника, состоящий из двояковогнутых или седловидных позвонков, к которым веерообразно прикреплены хвостовые перья. По строению клюва веерохвостые птицы делятся на зубастых (K3) и беззубых (современных), имеющих на клюве роговой чехол. Среди птиц известны плотоядные, растительноядные и всеядные формы. Подавляющее большинство птиц способно к полету. Несравненно меньшее распространение имеют бегающие или водоплавающие группы (страусы, пингвины). Полные скелеты ископаемых птиц встречаются крайне редко. Иногда сохраняются перья, скорлупа яиц, следы птиц. Большинство находок происходит из озерных, болотных и морских отложений. а б
Рис. 74. Археоптерикс: а – реконструкция, б – отпечаток на сланце (J)
Класс Млекопитающие (Mammalia), вскармливающие детенышей молоком, резко различны по размерам, облику, образу жизни (Приложение 10). Масса их тела варьирует в широчайших пределах: от землеройки весом в несколько грамм до китов весом в десятки тонн. Характерной особенностью млекопитающих является теплокровность и защитные приспособления, обеспечивающие постоянство температуры тела (волосяной покров или мощная жировая прослойка). Зубы дифференцированы. Разделение млекопитающих на подклассы базируется на способе размножения.
Подкласс Первозвери (T3 – ныне) вскармливают детенышей молоком, но размножаются яйцами. Строение скелета и наличие клоаки, куда открываются пищеварительный тракт, мочевые и половые протоки, сближают их с пресмыкающимися. В современной фауне подкласс представлен отрядом Однопроходные, достоверно известным с неогена. Представители этого отряда – утконос и ехидна – в настоящее время населяют Австралию и Новую Гвинею. Это – небольшие животные (длина тела до 60 см), ведущие наземный и полуводный образ жизни. К вымершим представителям этого подкласса принадлежат отряды Многобугорчатые (T3 – P2) и Докодонты (J3 – K1).

Рис. 75. Морганукодон – одно из самых древних млекопитающих планеты (T)
Условно в данный подкласс (рис.75) включают небольших крысоподобных животных, принадлежащих к отряду Трикодонты. Классификация данного подкласса остается дискуссионной.
ПодклассСумчатые(поздний триас? – поздний мел – ныне) рождают недоразвитых детенышей, которые вскармливаются молоком и донашиваются самкой в особых кожаных сумках. Выводковая сумка поддерживается костями таза. Степень дифференциации зубов и их число различно у разных групп. Из сумчатых наиболее известны кенгуру, опоссумы и коалы. Кроме того, в неформальной номенклатуре часто фигурируют формы, конвергентно сходные с соответствующими группами плацентарных (сумчатые кроты, мыши, куницы, волки). Размеры сумчатых варьируют от нескольких сантиметров до 3 м (кенгуру). Сумчатые представлены в современной фауне трех континентов: Австралии (7 отрядов), Северной и Южной Америк (3 отряда). Древние сумчатые были известны на всех континентах. Предполагается наличие общего предка сумчатых и плацентарных млекопитающих.
ПодклассПлацентарные(Высшие млекопитающие) появились в позднем мелу, а господствующее положение заняли с палеогена (Приложение 10). Мел-палеогеновый рубеж рассматривается как начало господства высших (плацентарных) млекопитающих, являющихся наиболее высокоорганизованными животными. Характерной особенностью этой группы является наличие плаценты – особого органа, осуществляющего связь материнского организма с детенышем до момента его рождения. Другими важнейшими особенностями плацентарных являются сложное строение головного мозга, отсутствие выводковой сумки, рождение беззубых детенышей, способных сосать материнское молоко.
Систематика плацентарных основана на строении зубного аппарата, черепа и конечностей. Выделяется около 30 отрядов, из которых в современной фауне представлено около 20. В раннемеловую эпоху появились насекомоядные, зайцеобразные и приматы, в позднемеловую – древние копытные. В раннепалеогеновых отложениях известны рукокрылые, хищники (креодонты и настоящие хищники), грызуны, непарнокопытные; в среднепалеогеновых – хоботные, китообразные, парнокопытные. Широчайшая радиация, обусловившая наступление «века млекопитающих», начинается с эоцена.
Отряд Насекомоядные (K1 – ныне) – всеядные животные с длиной тела 3,5-22 см. Зубной аппарат дифференцирован, но не специализирован. Мозг маленький, большие полушария гладкие.
Отряд Рукокрылые (P1 – ныне) представлен подотрядами Летучие мыши и Крыланы, различающимися, главным образом, размерами (первые – 2,5-14 см, вторые – до 40 см, в размахе крыльев – до 1,7 м). Один конец летательной кожистой перепонки рукокрылых крепится на пальцах передней конечности, другой – на задних конечностях. В зубном аппарате хорошо развиты клыки; предкоренные и коренные зубы слабо дифференцированы. Большие полушария головного мозга имеют малое количество извилин. По способу питания рукокрылые принадлежат к насекомоядным, растительноядным и кровососущим (вампиры).
Отряд Древние хищники (креодонты), включающий только ископаемые формы, существовал в течение палеогена и раннего неогена. Размеры тела варьировали от 10 см до 2,5 м. Головной мозг имел небольшие размеры, большие полушария были почти гладкими. Короткие конечности были преимущественно пятипалыми. Зубы – бугорчатые или бугорчато-режущие. Креодонты произошли от вымершей группы насекомоядных.
Отряд Настоящие хищники (Фиссипедии) имеют тело размером от 10 см до 4 м. Передние конечности обычно пятипалые, задние – четырехпалые. Характерны плотоядные зубы с режущими гребнями. Большие полушария головного мозга несут многочисленные извилины. Настоящие хищники подразделяются на собакоподобных (палеоген – ныне) и кошкоподобных (средний палеоген – ныне). Собакоподобные, преимущественно, вынюхивают добычу, кошкоподобные – высматривают. В разных филогенетических линиях кошек неоднократно появлялись саблезубые формы.
Отряд Зайцеобразные (K1 – ныне) включает в себя как ископаемые, так и современные группы. К последним принадлежат пищуховые и зайцевые, имеющие размеры тела 30-50 см. Передние конечности пятипалые, задние – четырехпалые. Верхняя челюсть несет постоянно растущие резцы. Клыки – отсутствуют, предкоренные и коренные зубы слабо дифференцированы. Полушария головного мозга почти гладкие, с малым количеством извилин. Ранее грызуны и зайцеобразные включались в один отряд, но позднее было доказано независимое от грызунов происхождение зайцеобразных от одной из групп насекомоядных.
Отряд Грызуны (P1 – ныне) представлен разнообразными группами с размерами тела 5-130 см (беличьи, хомяковые, мышиные и другие). Передние конечности пяти- или четырехпалые, задние – пяти- четырех- или трехпалые. Вторые резцы растут постоянно, клыки и предкоренные зубы отсутствуют. Число коренных зубов зависит от пищи. Полушарии почти гладкие, с малым количеством извилин. Грызуны имели общих предков с насекомоядными. Зубы грызунов, хорошо сохраняющиеся в ископаемом состоянии, используются для стратиграфии континентальных неоген-четвертичных отложений.
Копытные, по-видимому, являются филогенетически сборной группой, включающей отряды, характеризующие разные геологические периоды.
Отряд Кондилартры (K2) представлен животными с длинным хвостом, короткими пятипалыми конечностями, острыми копытами. Размер тела варьировал от 30 см до 2 м. Произошли от насекомоядных, но в палеогене в процессе радиации разделились на группы по способу питания. Одни стали родоначальниками китообразных и ластоногих, другие – предками непарнокопытных, хоботных, парнокопытных.
Отряд Непарнопалых (Непарнокопытных) объединяет несколько современных и ископаемых групп. Длина тела этих животных варьировала от 0,4 до 7 м; высота достигала 2-5 м. Конечности большинства непарнокопытных удлиненные, с различно оформленными копытами. Число пальцев на передних и задних конечностях – три или один, причем средний был наиболее крупным. В процессе эволюции число пальцев уменьшалось за счет редукции боковых.
К современным непарнокопытным принадлежат лошадиные (палеоген – ныне), тапировые (поздний палеоцен – ныне) и носороговые (средний палеоген – ныне); к ископаемым – гиракотериевые и титанотериевые (P2-3), халикотериевые (P2 – Q1), индрикотериевые (P3 –N1). Лошадиные вначале не превышали в высоту 1 м. Они обладали четырехпалыми передними и трехпалыми задними конечностями. Наибольшее распространение имели в Северной Америке, откуда неоднократно мигрировали на территорию Евразии. Носороговые характеризовались короткими массивными конечностями, вначале четырехпалыми, позднее – трехпалыми. Большинство носороговых были безрогими. Зубы характеризовались редукцией клыков и верхних резцов. Нижние резцы превратились в бивни. Места обитания носороговых весьма разнообразны. В раннечетвертичное время широкое распространение имел шерстистый носорог – предмет охоты древнего человека.
Из вымерших непарнокопытных наибольшими размерами характеризовались индрикотериевые. Их максимальные размеры достигали 7 м в длину и 5 м в высоту. Зубной аппарат был частично редуцирован. Передние резцы превратились в бивни, в верхней челюсти направленные вниз, а в нижней – вверх. Объем мозга был невелик, шея длинная. Конечности массивные трехпалые. Индрикотериевая фауна была широко распространена в течение олигоцена и миоцена в лесостепной зоне Евразии.
Специфическим признаком отряда Хоботные, является наличие хобота, образовавшегося за счет сращения носа и верхней губы. Конечности пятипалые, от коротких и средних до длинных и массивных. Отряд был широко распространен в течение неогена и плейстоцена, а в настоящее время почти вымер. Древние хоботные были высотой до 1 м, но в процессе эволюции достигли 4,5-5 м. Изменение размеров и облика связано с переходом от жизни в лесах к обитанию на открытых пространствах. Подразделяются на подотряды Меритерии, Дейнотерии и Слонообразные, из которых последние разделены на мастодонтов и слонов. Хоботные возникли в Африке, а затем распространились в Евразию и Северную Америку.
Отряд Китообразные – вторичноводные млекопитающие, постоянно живущие в водной среде. Дыхание легочное, но могут находиться под водой до 1,5 часов. По внешнему виду конвергентно сходны с рыбами. Роль плавников выполняют сильно укороченные пятипалые передние конечности. Задние конечности и кости таза сильно редуцированы, носовые отверстия смещены назад. Отряд подразделяется на группы Древние, Зубатые и Беззубые киты. Филогенетическое родство зубатых и беззубых китов вызывает сомнение.
Древние киты (эоцен – миоцен) по строению зубного аппарата напоминают кондилартр. К зубатым китам (эоцен – ныне) принадлежат дельфины, кашалоты, нарвалы, клюворылые. Зубы однообразны, число их увеличивается до 240 или уменьшается до двух. Зубатые киты – морские, реже речные или озерные животные, питающиеся рыбой, головоногими моллюсками, ракообразными. Длина тела 1,2 – 20 м. Беззубые (усатые) киты (олигоцен – ныне) вместо редуцированных зубов содержат в ротовой полости многочисленные роговые пластины («китовый ус»). Беззубые киты – морские животные, питающиеся планктоном или стайными мелкими рыбами, процеживая пищу через «усы». Длина тела до 33 м.
Отряд Парнокопытные подразделяется на жвачных и нежвачных. Современные представители жвачных появились в олигоцене (бычьи, оленьи), неогене (оленьковые, жирафовые, антилокапры) и плейстоцене (полорогие: газели, сайгаки, козы, бараны). Из нежвачных современных групп в эоцене появились свинообразные и пекари, неогене – гиппопотамы. Верблюдов и лам обычно выделяют из состава парнокопытных в особый отряд Мозоленогие. Длина тела современных парнокопытных варьирует в пределах 1,5-4,5 м, высота – 2-5,5 м. Кроме того, в составе данного отряда имеются ископаемые группы. Обособление парнокопытных в особую группу обусловлено их отличиями в способе передвижения и мало связано с особенностями строения зубного аппарата.
Отряд Приматы (Приложение 10) произошел в раннемеловую эпоху от группы животных близких к насекомоядным. Он подразделяется на подотряды Полуобезьяны (Низшие приматы) и Обезьяны (Высшие приматы). К первому подотряду (эоцен – ныне) принадлежат лемуры, долгопяты, лори и близкие им формы. Ко второму – относятся широконосые обезьяны Америки, подразделяющиеся на игрунковых и цепкохвостых, а также узконосые обезьяны Старого Света (мартышкообразные и человекообразные). Длина тела обезьян варьирует от 10 см до 1,5-2 м. Пятипалые конечности обладают высокой хватательной способностью. Останки древнейшего представителя высших приматов найдены в эоценовых отложениях Мьянмы (Бирма). В Африке останки высших приматов известны с олигоцена. Предок человека и человекообразных обезьян – дриопитек появился 25-30 млн. лет назад в олигоцене. Был жителем влажных тропических лесов. Вел древесный образ жизни, питаясь растениями и насекомыми (рис.76).

Рис. 76.
Дриопитек (P3).
Человекообразные имеют длинные руки, короткие ноги и сравнительно развитый головной мозг. Около 9 млн. лет назад…Рис. 78. Человек умелый
(Homo habilis)
(3-2 млн. лет назад)

Архантропы третьей группы иногда выделяются в качестве вида H. heidelbergensis. На территории Евразии они продвинулись в северном направлении до Юго-Западной Германии, а возможно и дальше. В Африке в это время обитал H. helmei.

Рис. 80. Человек выпрямленный
(Homo erectus)
(1.74 млн.–130 т.л.н.)
По особенностям строения скелета (в том числе, кистей рук) архантропы весьма сходны с современным человеком, но резко отличаются формой черепа, указывающей на различия в структуре мозга. Череп архантропов характеризовался большими размерами челюстей, значительной толщиной стенок, покатым лбом, нависающими надбровными дугами, выступающим назад затылком. Формы, относимые к H. heidelbergensis отличались от H. erectus большим объемом мозга и округлым затылком. Речевой аппарат и соответствующий центр мозга у архантропов или отсутствовали, или находились в зачаточном состоянии. Поздние архантропы умели пользоваться огнем и изобрели примитивный язык для коллективной охоты.
Орудия архантропов отличаются слабой степенью обработки каменных обломков, главным образом, галек. Типичным орудием было ручное рубило с грубым режущим краем. Кроме того, архантропы делали деревянные копья. По степени обработки каменных орудий их можно отнести к раннему палеолиту. Факт изготовления орудий труда архантропами качественно отличает их от остального органического мира Земли, подтверждая принадлежность к роду Homo.
 Интервал времени 120-35 тыс. лет обычно рассматривается как время распространения палеоантропов, представленных на территории Евразии неандертальцами, систематическое положение которых остается дискуссионным. Одни исследователи относят их к виду H. neanderthalensis, другие – к подвиду H. sapiens neanderthalensis. Иногда к палеоантропам относят вид H. heidelbergensis. Характерной особенностью неандертальцев является их приспособленность к существованию в холодном климате. Это были люди невысокого роста (150-160 см) с широкими плечами, короткими ногами, бочкообразной грудью, мощной мускулатурой. Объем мозга был – 1330 см3. По облику они несколько напоминали представителей современных северных народов – чукчей и эскимосов.
Интервал времени 120-35 тыс. лет обычно рассматривается как время распространения палеоантропов, представленных на территории Евразии неандертальцами, систематическое положение которых остается дискуссионным. Одни исследователи относят их к виду H. neanderthalensis, другие – к подвиду H. sapiens neanderthalensis. Иногда к палеоантропам относят вид H. heidelbergensis. Характерной особенностью неандертальцев является их приспособленность к существованию в холодном климате. Это были люди невысокого роста (150-160 см) с широкими плечами, короткими ногами, бочкообразной грудью, мощной мускулатурой. Объем мозга был – 1330 см3. По облику они несколько напоминали представителей современных северных народов – чукчей и эскимосов.
Рис. 81. Homo sapiens neanderthalensis
(120-35 тыс. лет назад)
Когда в Европе появились кроманьонцы, неандертальцы не менее 5 тыс. лет сосуществовали с ними и, возможно, жили в одних и тех же пещерах. В технике изготовления орудий труда среднего палеолита господствовало изготовление кремневых дисковидных отщепов, использовавшихся затем для получения остроконечников на копья и скребков для обработки шкур. В связи с холодным климатом появилась одежда. В технике обработки орудий труда неандертальцы, видимо, значительно отставали от кроманьонцев. С другой стороны, у них раньше, чем у кроманьонцев пробудились черты гуманизма: племя выделяло пищу больным и старикам. Предметов искусства и украшенийот неандертальцев не осталось. Вероятно, у неандертальцев существовал ритуал погребения. В то же время, по мнению многих исследователей, у неандертальцев имел место каннибализм.
В генотипе современного человека неандертальцы представлены слабо, что позволяет считать их тупиковой ветвью рода Homo. Возможной причиной исчезновения неандертальцев являлась высокая детская смертность. В диете неандертальцев главную роль играла животная пища, что дает основание считать их основными хищниками своего времени. Вероятно, не случайно при захоронении покойников неандертальцы нередко клали вместе с трупом мертвых хищников, а не травоядных животных.
Около 40 тыс. лет назад в Европе появились пришедшие из Африки неоантропы (кроманьонцы), практически не отличавшиеся по облику от современного человека. По сравнению с неандертальцами кроманьонцы характеризовались более высоким уровнем обработки орудий труда (верхний палеолит). Кремневые орудия стали изготавливаться из узких длинных заготовок. Наряду с кремнием широко использовались кости животных, в первую очередь мамонта. Разнообразие орудий труда кроманьонцев отличало их от весьма однообразных орудий неандертальцев. В начале верхнего палеолита появились копьеметалки, а в конце – лук и стрелы. Строятся постоянные зимние общественные жилища: землянки и наземные постройки, начинается одомашнивание животных.
Происходит расцвет верхнепалеолитического искусства. В пещерах Евразии сохранились превосходные образцы наскальной живописи. В слоях стоянок обнаружены статуэтки людей и животных, сделанные из бивней мамонта и известняка. Одежда украшалась бусами и раскрашивалась охрой. Значительной сложностью отличался ритуал похорон. Усопшему клали орудия, каменные ножи, многочисленные украшения. По-видимому, в это время возникают представления о загробном мире.
В течение верхнего палеолита заканчивается освоение человеком континентов. В Австралию человек проник около 60 тыс. лет назад. Возможно, после этого произошло опустынивание континента и исчезновение лесов, поскольку аборигены использовали лес в качестве топлива. В Северную Америку люди проникли через Берингов перешеек около 35 тыс. лет назад, когда произошло осушение Берингии.
Граница палеолит-мезолит (15-13 тыс. лет назад) близка ко времени исчезновения покровного оледенения в Евразии и Северной Америке (около 10 тыс. лет назад). Люди стали заселять освободившиеся территории. Ускорилось одомашнивание животных, появились первые зоны земледелия и скотоводства.
Началом неолита считается изобретение гончарного круга, концом – возникновение металлургии. В связи с неравномерностью развития населения разных территорий неолит наступал в разное время (от 6-5 до 2 тыс. лет до н.э.). Характерными чертами неолита являются высокое качество обработки каменных орудий и оружия, возникновение ткачества и транспортных средств, развитие рыболовства. Около 5 тыс. лет назад появились первые бронзовые изделия, а 3-2,5 тыс. лет назад началась эпоха железа. Однако каменное оружие использовалось и значительно позднее. Саксы в битве при Гастингсе (XI век н.э.) использовали каменные топоры, а зулусы в боях с англичанами в XIX веке пользовались копьями с обсидиановыми наконечниками. Остатки каменных культур наряду с органическими остатками и данными радиогеохронологии широко используются при разработке детальной стратиграфии четвертичного периода.
Вопросы для самоконтроля:
1. Сравнить строение пластинокожих, акантод и хрящевых рыб.
2. Назвать и описать особенности, группы организмов, которая произошла от рыб и дала начало рептилиям.
3. Назовите важнейших представителей современных и древних рептилий, укажите принципы их классификации.
4. Охарактеризовать современную классификацию птиц.
5. Назовите классификационные признаки класса Млекопитающие.
6. Изложить современные представления о происхождении и эволюции человека.
7. Кто такой неандерталец? Кем он являлся?
8. Охарактеризовать историю развития неоантропов.
Часть II. ИСТОРИЧЕСКАЯ ГЕОЛОГИЯ
ОБЩИЕ ПОЛОЖЕНИЯ
Историческая геология ставит своей целью изучение истории развития планеты… Геохронология – календарь геологических событий, охватывающий 4,6 млрд. лет. Её основу составляет радиометрическое…Вопросы для самоконтроля
1.Назвать цель, задачи и основные направления исторической геологии.
2.Объяснить, в чем состоит сциально-экономический эффект историко-геологических исследований.
3. Перечислить этапы развития исторической геологии, назовите особенности каждого этапа.
4.Объяснить роль исторической геологии в формировании элементов научного мировоззрения.
Стратиграфия
2.1. Методы определения относительного возрастаБ
Рис. 82 . Применение стратиграфического метода
Тектоно-стратиграфический метод основан на различиях в степени метаморфизма и дислоцированности горных пород. Применяется при расчленении докембрийских отложений, накопившихся в течение огромных интевалов геологического времени и практически лишенных органических остатков.
Литолого-петрографический метод (литологический анализ), неразрывно связанный с фациальным анализом, рассматривается ниже.
Геофизические методы обычно используются для корреляции разрезов между собой, когда возраст пород определен другими методами. С помощью каротажа скважин прослеживаются по простиранию геологические границы отложений, резко различающихся электрическими, механическими и другими свойствами. Кроме того, при поисках нефтегазоносных месторождений с помощью сейсмических особенностей горных пород удается выявить участки, перспективные для разведки на нефть и газ. Палеомагнитный метод разработан в результате изучения магнитного поля Земли прошлых геологических эпох, зафиксированного в горных породах. Палеомагнитные исследования начались в середине ХХ века. К настоящему времени разработано две методики определения возраста горных пород по палеомагнитным данным: 1) на основе изучения миграции магнитных полюсов, 2) на основе перемагничивания Земли, благодаря чему Северный и Южный магнитные полюса менялись местами (геомагнитные инверсии). При миграции магнитных полюсов (рис. 83) геологический возраст анализируемой горной породы определяется в три этапа. На первом этапе выделяется вектор первичной намагниченности, соответствующийположению магнитного меридиана в момент образования данной породы. Для этого устраняется воздействие современного магнитного поля (вектор остаточной
 |
намагниченности) и магнитных полей, имевших место в течение времени существования данной горной породы (вектор вязкой
Рис. 83. Кривые блуждания полюса в фанерозое для Европы (квадратики) и Северной Америки (кружки) при нынешнем расположении материков; C – кембрийский, 550 млн. лет; S–D – силуро–девонский, 390 млн. лет; S–C1 – силуро–нижнекарбоновый, около 350 млн. лет; С3 – верхнекарбоновый, 300 млн. лет; Р – пермский, 250 млн. лет; Т – триасовый; Т1 – нижнетриасовый, 220 млн. лет; J – юрский, 150 млн. лет; К – меловой, 100 млн. лет
намагниченности). На втором этапе определяется перемещение магнитного полюса в течение предполагаемого времени существования породы. Для этой цели требуется значительное количество разновозрастных образцов, отобранных с более или менее значительной территории.
На третьем этапе определяется точка пересечения магнитного меридиана, установленного в итоге исследований первого этапа, с кривой движения магнитного полюса. Полученная точка указывает возраст анализируемой породы в единицах относительной или абсолютной геохронологии (в зависимости от градуировки кривой движения магнитного полюса). Эта методика весьма трудоемка, требует большого количества образцов, а потому не получила широкого распространения.
Геомагнитные инверсии неоднократно происходили в истории Земли, причем проявлялись неравномерно. Выделяются эпохи сгущения и разряжения инверсий. К настоящему времени на основе изучения геомагнитных инверсий построена палеомагнитостратиграфическая шкала фанерозоя России.
Результаты палеомагнитных исследований используются сторонниками мобилизма для доказательства движения литосферных плит.
Палеонтологический метод, широко используется в стратиграфии. Её отрасль, основанная на использовании этого метода, получила название биостратиграфия. Важнейшим биостратиграфическим методом является метод руководящих ископаемых, сыгравший важную роль в установлении подразделений Международной биостратиграфической шкалы (МБШ). Руководящими формами называют остатки вымерших организмов, живших короткий отрезок времени, но имевших широкое географическое распространение. Оптимальными в этом плане являются конодонты – остатки челюстного аппарата морских червей. В идеальном случае конодонты позволяют решить обе задачи стратиграфии: расчленить толщи горных пород и скоррелировать одновозрастные отложения. Недостатком метода является неодновременность вымирания групп организмов. Например, в Австралии до настоящего времени существуют сумчатые млекопитающие, вымершие на остальных континентах. Для уменьшения данного недостатка было разработано несколько методик: руководящих комплексов, математического анализа и другие.
Основное правило палеонтологического метода – пласты горных пород, содержащие одинаковый комплекс окаменелостей, одновозрастны. С помощью этого метода были, в частности, сопоставлены по возрасту известняки карьеров г. Гурьевска с породами окрестностей г. Праги (Чехия). Обратное правило – пласты, содержащие различные комплексы окаменелостей, разновозрастны не всегда справедливо, так как разный состав окаменелостей может быть вызван как возрастными различиями, так и разной средой обитания. Широкое использование палеонтологического метода позволило исследователям XIX века охарактеризовать органическими остатками отложения каждого периода палеозойской, мезозойской и кайнозойской эр. В целом, стратиграфический и палеонтологический методы геохронологии обеспечили детальное изучение отложений фанерозойского эона, но оказались малопригодными для докембрийских отложений, отличающихся высокой дислоцированностью и в значительной мере лишенных органических остатков.
Все эти методы отвечают на вопрос об относительном (сравнительном) возрасте пластов горных пород. На основе относительной геохронологии в 1881 году на Международном геологическом конгрессе в г. Болонье (Италия) был принят первый вариант Международной биостратиграфической шкалы, содержащий палеонтологические характеристики (списки видов) каждого геологического периода палеозойской, мезозойской и кайнозойской эр.
2.2. Методы определения абсолютного возраста
Геологические процессы отличаются от процессов обыденной жизни чрезвычайной продолжительностью, в силу чего для них оказываются непригодными единицы измерения, применяемые в быту (минуты, часы, сутки, недели, месяцы, годы). Основной единицей измерения продолжительности геологических процессов являются миллионы лет. Только при измерении продолжительности последнего отрезка геологического времени – четвертичного периода – единицей измерения становятся тысячелетия.
Для определения количества времени, прошедшего после какого-либо геологического события, нужны «геологические часы», то есть процесс, во-первых, соизмеримый по длительности с остальными геологическими процессами, во-вторых, характеризующийся постоянной скоростью, не меняющейся от воздействия других процессов. В течение первой половины XIX века обнаружить такой процесс не удавалось. Мощность осадочных толщ, степень метаморфизма горных пород, накопление солей на дне Мирового океана возрастают с течением времени, но, как показали дальнейшие исследования, скорость этих процессов различна на различных участках Земного шара.
Только в конце XIX века трудами французского ученого Беккереля и супругов Пьера и Марии Кюри-Склодовских был обнаружен радиоактивный распад – процесс превращения одного химического элемента в другой, сопровождающийся выделением энергии в виде радиоактивного излучения. Химические элементы, характеризующиеся способностью к радиоактивному распаду, называются радиоактивными (радий, торий, уран и некоторые другие). Установлено, что многие элементы, считающиеся нерадиоактивными, в небольшом количестве содержат радиоактивные изотопы. Время превращения половины какого-либо количества радиоактивного элемента в другой элемент обычно нерадиоактивный (период полураспада) есть величина постоянная, установленная лабораторными исследованиями для каждой пары элементов.
Первые попытки использования радиоактивного распада для определения возраста горных пород относятся к началу ХХ века. Широкое распространение радиометрический (радиологический)метод, известный в качестве «абсолютной геохронологии», получил во второй половине ХХ века. Название «абсолютная» здесь не совсем уместно, так как радиометрический метод (подразделяющийся на несколько частных методик) также не свободен от ошибок.
Использование радиометрического метода предполагает существование в природе замкнутой системы, в которой исходный радиоактивный элемент постепенно превращается в конечный продукт распада, причем вынос начального и конечного продуктов отсутствует. В этом случае, определив лабораторным путем содержания в породе исходного и конечного продуктов распада и зная период полураспада, можно определить его длительность, то есть возраст породы. В действительности начальный и конечный продукты распада частично выносятся в результате процессов метаморфизма, причем их количество зачастую не поддается учету. В силу этого возможны существенные ошибки при определении возраста пород радиометрическим методом. Во избежание ошибок, обычно используется несколько пар элементов и, соответственно, выделяется несколько методик (частных методов) радиометрического датирования.
Таблица 1
| Радиоактивный элемент (изотоп) | Конечный продукт | Период полураспада млрд лет |
| 238U | 206Pb | 4,468 |
| 235U | 207Pb | 0,7038 |
| 232Th | 208Pb | 14,008 |
| 87Rb | 87Sr | 4,88 |
| 40K | 40Ar | 1,3 |
| 14C | 14N | 5750 лет |
Урано-ториево-свинцовый метод основан на распаде 238U→ 206Pb, 235U→ 207Pb, 232Th→ 208Th→ 208Pb. В последние годы определение возраста горных пород чаще всего проводится по отношению U/Pb в кристаллической решетке минерала циркона. Атомы урана частично замещают атомы циркония в кристаллической решетке циркона, а в результате распада урана образуется свинец.
Рубидий-стронциевый метод основан на распаде радиоактивного изотопа 87Rb и превращения его в изотоп 87Sr. Этот метод широко используется при определении геологического возраста древнейших горных пород.
Калий-аргоновый метод, предложенный в 1949 году советским ученым Э.К. Герлингом, основан на распаде радиоактивного изотопа 40К и частичном превращении его в изотоп 40Ar с периодом полураспада 1300 млн. лет.
Радиоуглеродный метод основан на измерении содержания радиоактивного изотопа углерода, присутствующего в органических остатках и породах с высоким содержанием органического вещества. Организмы усваивают радиоактивный углерод из воздуха, а после их отмирания начинается его распад. Период полураспада 14С→14N равен 5750 лет, поэтому радиоуглеродный метод используется для датировки осадков не древнее 60-80 тыс. лет. Этот метод широко применяется для датировки отложений четвертичного периода, а также в археологии.
В течение ХХ века на основе абсолютной геохронологии была установлена продолжительность подразделений Международной биостратиграфической шкалы (МБШ).
2.3. Стратиграфические и геохронологические подразделения
Современная шкала геологического времени является одновременно стратиграфической и геохронологической. С одной стороны, по данным палеонтологического и стратиграфического методов характеризуется органический мир каждого подразделения МБШ, а также его вымирания и возрождения на границах подразделений. С другой – по данным абсолютной геохронологии указываются возраст границ этих подразделений (обычно с точностью до 1-2 млн. лет). Международная шкала геологического времени совершенствуется и в настоящее время. Её последний вариант был принят в 2008 году на Международном конгрессе в Осло (Норвегия).
В России высшим законодательным органом для геологических организаций является Межведомственный стратиграфический комитет (МСК), котрый каждые четыре года пересматривает МБШ и вносит в нее изменения, согласно новым данным науки. По постановлению МСК в 2006 году в России принята Общая стратиграфическая шкала, подразделения которой делятся на стратиграфические и геохронологические.
Таблица 2
| Стратиграфические подразделения | Геохронологические подразделения |
| 1.Акротема | Акрон |
| 2.Эонотема | Эон |
| 3.Эратема | Эра |
| 4.Система | Период |
| 5.Отдел | Эпоха |
| 6.Ярус | Век |
| 7.Зона | Фаза |
| 8.Звено | Пора |
Акротема – наиболее крупное подразделение, появившееся в 1992 году, поскольку подразделения докембрия – архей и протерозой – резко отличаются большей продолжительностью от остальных подразделений.
Эонотема – отложения, образовавшиеся в течение эона, длившегося многие сотни миллионов лет. В составе архейской и протерозойской акротем выделяют по две эонотемы. Рангу эонотемы соответствует также фанерозой.
Эратемы(группы) – крупные стратиграфические подразделения, имеющие значительную мощность, большие площади распространения, сложный состав. Образуются в течение одной эры. Границы между эрами проводятся по резким изменениям в составе органического мира. Выделяют палеозойскую (PZ), мезозойскую (MZ), и кайнозойскую (KZ) эры, входящие в состав фанерозоя.
Система– часть эратемы, образовавшаяся в течение одного периода. Система характеризуется типичными для неё семействами или родами фауны и флоры. Границы между системами палеонтологические или тектонические. Обозначается заглавной буквой латинского алфавита (S, D, P).
Отдел – часть системы, образовавшаяся в течение одной эпохи. Характеризуются отделы определенными родами и видами фауны и флоры. Отдел бывает нижний, средний, верхний; эпохи соответственно – ранняя, средняя, поздняя. Обозначается цифрой (D1).
Ярус – часть отдела, образовавшаяся в течение века, характеризуется определенным комплексом родов и видов ископаемых организмов. Ярус имеет стратотип с определенной палеонтологической характеристикой. Названия ярусов происходят от географических названий мест, где расположен стратотип данного яруса. Индекс яруса обозначается начальной буквой латинского алфавита или двумя буквами (D1 gv).
Зона – часть яруса, накопившаяся в течение одной фазы. Границы её устанавливаются по характерному комплексу видов. Стратотип зоны рекомендуют выбирать в стратотипе яруса. Название зоны и её фаза дается по характерному виду комплекса (зона Altaiophyllum belgebaschicum).
Звено выделяется лишь в четвертичной системе. Пора, за время которой формируются отложения звена, соответствует изменению климата – двум полуциклам: потепление, похолодание. Звено должно иметь стратотип. Четвертичная система слагается из четырех звеньев, которые обозначаются римскими цифрами (QII).
В 1977 году в нашей стране впервые был опубликован Стратиграфический кодекс, выполнение которого обязательно при проведении геологических работ. В кодексе принята стратиграфическая классификация, согласно которой выделяются основные и вспомогательные стратиграфические подразделения. К основным подразделениям относятся: общие, региональные и местные подразделения. При выделении общих стратиграфических подразделений ведущим является палеонтологический метод. Исключение составляет докембрий, расчленение которого осуществляется на основе тектоно-стратиграфического и радиометрического методов. Из общих подразделений ярус, зона и звено должны иметь стратотип – конкретный, наиболее полный разрез данного подразделения, эталон для сравнения. Его можно увидеть, изучить, взять образцы. Для остальных общих стратиграфических подразделений выделение стратотипов не предусмотрено.
Региональные стратиграфические подразделения не имеют всепланетарного охвата. Они устанавливаются для геологического региона, крупного палеобассейна седиментации или палеобиогеографической области. Основной единицей здесь является горизонт, представляющий совокупность одновозрастных свит, по одной из них он получает свое название. Горизонты фанерозойских толщ чаще всего устанавливаются с помощью методов биостратиграфии. Горизонты в докембрийских образованиях, а также в палеонтологически «немых» толщах устанавливаются на основе литолого-фациальных и петрографических особенностей пород. Горизонты четвертичной системы устанавливаются на климатостратиграфической основе. Горизонт имеет стратотип и получает географическое название места, где расположен его стратотип (алчедатский горизонт, сафоновский горизонт). Региональным стратиграфическим подразделением является лона, устанавливаемая по палеонтологическим данным. Название её дается по виду-индексу (например, лона Tabulophyllum schluteri Peetz). Геохронологическим эквивалентом горизонта и лоны является время.
Основным подразделением местной стратиграфической шкалыявляется свита, устанавливаемая на сравнительно небольших территориях. Она объединяет одинаковые или близкие по литолого-фациальным особенностям одновозрастные отложения и отражает определенный этап развития данной территории. Свита имеет стратотип. Свое название она получает по географическому названию стратотипа. Пример: полюдовская свита, тайгонская свита. Свита подразделяется на подсвиты.
Серия – более крупная единица по сравнению со свитой. Она объединяет несколько свит по какому-либо общему признаку (например: морская серия, вулканическая серия). Серии присваивается стратотип одной из ее свит.
Комплекс объединяет несколько серий и имеет собственное название. Это мощная и сложная по составу и структуре совокупность геологических тел. Комплекс отвечает крупному этапу геологического развития обширной территории. Обычно не имеет стратотипа и используется для докембрийских образований. Пример: карельский комплекс.
Кроме основных стратиграфических подразделений существуют ещё вспомогательные, их называют также «термины свободного пользования». Эти подразделения не являются обязательными, их употребляют по желанию. Сюда относятся: толща, пачка, слой, маркирующий горизонт, слои с фауной, флорой. Частные биостратиграфические подразделения – биостратиграфические зоны разных видов выделяются по палеонтологическим признакам. Они имеют стратотип и могут подразделяться на подзоны. Их геохронологический эквивалент – время.
Региональные и местные стратиграфические подразделения сводятся в таблицы, называемые стратиграфическими схемами. Региональная стратиграфическая схема представляет собой корреляцию региональных и местных стратонов с общей стратиграфической шкалой и шкалами смежных регионов. Она состоит из четырех разделов: Общей стратиграфической шкалы, региональных стратиграфичсеских подразделений, корреляции местных разрезов и региональных подразделений смежных регионов. По степени разработанности региональные стратиграфические схемы подразделяются на унифицированные, корреляционные и рабочие. Унифицированные стратиграфические схемы включают в себя все четыре раздела региональной схемы. В корреляционных схемах отсутствует или представлен лишь частично раздел «региональные стратиграфические подразделения». Рабочая схема базируется на недостаточно аргументированных данных.
2.4. Принципы стратиграфии
Вопрос об основных принципах стратиграфии, составляющих её теоретическую основу, неоднократно рассматривался разными исследователями (В. Смитом, Ч. Дарвиным, Т. Гексли, Л. Долло, Л.Л. Халфиным, Д.Л. Степановым и М.С. Месежниковым, С.В. Мейеным и другими). Итоги многолетней дискуссии проанализированы в работе Н.И. Савиной (2002), предложившей выделять четыре принципа.
Первый принцип (принцип последовательности напластования), сформулированный Н. Стеноном, обеспечивает перевод пространственных взаимоотношений геологических тел, залегающих выше или ниже друг друга, во временной режим (моложе или древнее).
Второй принцип (принцип биостратиграфического расчленения и корреляции) гласит, что при корреляции разрезов, если непосредственное прослеживание невозможно, может быть применена система признаков сходства этих отложений (принцип Гексли). Одним из признаков сходства сравниваемых отложений может быть наличие в них одинакового комплекса ископаемых органических остатков (принцип Смита).
Третий принцип позволяет заменять одни признаки другими (например, редкие находки органических остатков заменяются литологическим сходством отложений). Этот принцип издавна применялся в практике геологических исследований, в частности, при корреляции морских и континентальных отложений. На этом принципе основана универсальность МБШ.
Четвертый принцип – объективной реальности и неповторимости стратонов – объединяет два самостоятельных принципа Л.Л. Халфина. Каждый стратон не только объективно отражает среду накопления, но и является уникальным, покольку уникальны (неповторимы) условия его накопления. Этот принцип указывает, что при стратиграфических исследованиях следует учитывать не только временные взаимоотношения стратонов, но и их значимость как объективных источников информации о географической обстановке прошлых геологических эпох.
С точки зрения авторов, следует выделить пятый принцип – необратимость эволюции органического мира (принцип Дарвина-Долло). Согласно этому принципу, вымерший вид не может появиться в процессе эволюции второй раз. Этот принцип был сформулирован на основе эмпирических наблюдений. Сейчас он подтверждён исследованиями генетического аппарата живых организмов: утеря генотипа невосполнима естественным путем.
Вызывает сомнения сформулированный Ч. Дарвиным принцип неполноты геологической летописи. Этот принцип справедлив для каждого частного разреза, но, по-видимому, не может быть признан в глобальном масштабе. Невозможно представить, чтобы процессы денудации были повсеместными (где-то имело место осадконакопление).
Вопросы для самоконтроля
1. Укажите отличие палеонтологического метода относительной геохронологии от стратиграфического.
2. Перечислите пары элементов, используемые в радиометрии («абсолютной геохронологии»).
3. Назовите источники ошибок в определении абсолютного возраста.
4. Назовите основные группы стратиграфических подразделений.
5. Охарактеризуйте специфику стратиграфических схем.
6. Перечислите основные принципы стратиграфии.
Фациальный анализ
Понятие «фация» появилось в геологической литературе в XIX веке. В настоящее время это понятие используется и в географии, и в геологии. Географы… Геология имеет дело с осадками и горными породами, поэтому геологи… В процессе фациального анализа изучаются все элементы современной физико-географической обстановки (рельеф, климат,…Рис. 85. Палеобиоценоз прибрежной области.
Все организмы связаны совместным проживанием и цепью питания

Рис. 86 . Тафоценоз побережья Австралии
Волнами переотложены остатки наземных растений,
водорослей, морских ежей, губок.
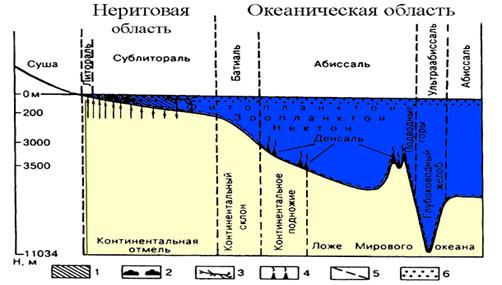
Рис. 87. Биономические зоны моря со сгущениями жизни.
1 – наибольшая концентрация жизни, 2 – рифы,3 – скопление планктонных водорослей, 4 – денсаль (зона жизни организмов, существующих за счет хемосинтеза) , 5 – бентосная пленка жизни, 6 – планктонная пленка жизни
населен организмами и приповерхностный слой (до глубины 50-80 м) батиальной и абиссальной зон (рис. 87, 1). Здесь обитает богатый фито- и зоопланктон, питающиеся им организмы, а также хищники.
На континентах ископаемые остатки организмов встречаются сравнительно редко и лишь в относительно молодых породах (начиная с девонского периода). Главными факторами, определяющими условия накопления континентальных осадков, являются климат и рельеф. Обычно встречаются отпечатки древних растений и значительно реже – остатки животных (пресноводные моллюски, насекомые, ракообразные). В отложениях мезо-кайнозоя нередки остатки позвоночных.
Фации нормальноморских бассейнов
Неритовая область, соответствующая шельфу (глубины до 200 м, иногда до 500 м), превышает по удельной биомассе абиссаль в 200 тысяч раз. Она… Отложения неритовой области представлены терригенными, органогенными,… К батиальной области (глубины приблизительно от 200 до 2000 м) относится морское дно, соответствующее континентальному…Рис. 90 . Осадки, скапливающиеся у подножия континентального склона
Абиссальная область соответствует ложу Мирового океана, в котором выделяют крупные океанические котловины, подводные хребты, возвышенности, плато. Для неё характерно полное отсутствие света, пониженная температура воды (от 1,87 до 4°С), громадное давление и повышенное содержание углекислоты, так как глубже 4,5-5 км карбонаты переходят в раствор. Все это неблагоприятно влияет на расселение организмов, однако жизнь на абиссальных глубинах существует. В основном это пелагические формы – рыбы, моллюски, планктон. Из бентосных форм характерны иглокожие, брахиоподы, двустворчатые моллюски. Много сидячих форм с длинными стеблями (губки, восьмилучевые кораллы). Для организмов характерно замещение известкового скелета кожистым, хитиновым или кремнистым. У костистых рыб понижено окостенение скелета.
Абиссальная зона до глубин 4,5-5 км покрыта карбонатными илами, состоящими из раковин фораминифер и водорослей (фитопланктона). На глубинах более 4500 м, где карбонаты растворяются, накапливаются кремнистые осадки: красная глубоководная глина и радиоляриевые илы. Состав осадков по латерали в значительной мере контролируется климатическим фактором. В тропическом и субтропическом поясах наибольшее распространение имеет карбонатный глобигериновый (фораминиферовый) ил, образовавшийся за счет скопления скелетных остатков глобигерин, населяющих приповерхностный слой Мирового океана. В Атлантическом океане за счет теплого Северо-Атлантического течения область накопления глобигериновых илов распространяется далеко на север. В холодных морях северной и южной окраин Тихоокеанской впадины накапливаются кремнистые диатомовые илы – скопления скелетных остатков микроскопических водорослей (диатомей).
Красная глубоководная глина, имеющая красновато-коричневый цвет и сложенная частицами размером менее 0,005 мм является весьма характерным типом осадков абиссали. Она имеет эоловое происхождение и состоит в основном из космической и вулканической пыли, зубов акул. Красновато-коричневая окраска обусловлена наличием окислов железа. Скорость накопления красной океанической глины очень мала (не более 1мм в 1000 лет). С красной океанической глиной обычно связаны железомарганцевые конкреции, местами покрывающие 70-90% поверхности дна. В будущем добыча железа и марганца на планете будет производиться за счет разработки залежей этих конкреций, и первые шаги в этом направлении уже сделаны. В рифтовых зонах срединно-океанических хребтов абиссальной области происходит излияние базальтовых лав.
В составе осадков ультраабиссали (глубины от 6 до 11 км) также преобладает красная глубоководная глина. В конце прошлого века здесь обнаружили «оазисы» жизни, которые располагаются вокруг «черных курильщиков». Это высокие донные образования, из верхушек которых поступают перегретые воды – гидротермы, черного цвета, так как содержат сульфиды. Живые организмы, обитающие вокруг гидротерм, питаются за счет бактерий, использующих сульфиды для хемосинтеза. В основном это черви, полихеты, 7 видов рыб, иглокожие, членистоногие. Здесь обнаружен особый тип животных – погонофоры. Это крупные червеобразные животные (длиной до 1,5 м), живущие в гибких цилиндрических трубках из белка и хитина (рис. 91). Питание погонофор происходит за счёт симбиоза с хемотрофными бактериями, поэтому органов пищеварения у них нет. Такой способ питания присущ многим животным глубоководных «оазисов».

Рис. 91. Погонофоры на фоне «черных курильщиков».
Впервые были открыты в 1977 году на Галапагосских островах
Переходные фации.
в а б Эстуариями называются устья рек, расширенные в… Лагуна – часть моря, отделенная косой или барьерным рифом. Заливом называется… и богатые органическимвеществом илы, а также накапливаются бокситы и железные руды. В аридном климате происходит…Континентальные фации
1. Малые мощности отложений (до первых километров). 2. Резкая смена фаций по латерали и разрезе. 3. Климатическая зональность.Вопросы для самоконтроля
1. Дайте определение фации.
2. Охарактеризуйте литологический и биономический методы фациального анализа.
3. Охарактеризуйте фации нормальноморских бассейнов, переходные и континентальные.
4. Назовите факторы, в наибольшей мере влияющие на распределение организмов в морских бассейнах.
5. Назовите особенности палеогеографических карт по сравнению с географическими.
ТЕКТОНИЧЕСКИЙ АНАЛИЗ
Состав и строение Земного шара кардинально меняются с глубиной (от поверхности к центру). Оболочки Земли получили название геосфер. В какой-то мере… Важной задачей исследования земных недр является изучение закономерностей… В последние десятилетия вместо подразделения «земная кора» (мощность от 5-10 до 30-60 км) обычно употребляется…Рис. 93. Схема движения литосферных плит и строения земной коры.
1 – верхняя мантия (стрелками показаны конвекционные движения), 2 – срединно-океанические хребты (зона спрединга), 3 – базальтовый слой океанической коры, 4 - гранитный слой континентальной коры, 5 – вулканияеские горы над зонами субдукции, 6 – базальтовый слой континентальной коры, 7 – молодые складчатые горы, 8 – острова, 9 - 10 – глубоководные желоба (зоны субдукции), 11 – островные дуги
При погружении происходит её разогрев за счет трения. Морские осадки, соскабливающиеся с плиты (процесс аккреции), частично
расплавляются, превращаясь в магму, частично подвергаются метаморфизму. Становится понятным, почему часть гранитов имеет магматическое происхождение, а другая образовалась за счет процессов метаморфизма.Первоначально тектоника плит рассматривалась как альтернатива учению о тектонически пассивных и активных геоструктурах – платформах и геосинклиналях. Это учение было разработано во второй половине XIX века Д. Холлом, Д. Дана и А.П. Карпинским. Позднее были выявлены черты сходства обеих концепций. В частности, представления о формировании гранитов в зоне субдукции в какой-то мере соответствуют концепции этапности развития геосинклинали, превращающейся в результате консолидации в участок земной коры континентального типа.
По мере накопления фактического материала стало очевидным, что тектоника плит, основанная на признании ведущей роли горизонтальных дислокаций, не отражает в полной мере геологические процессы недр Земли. В частности, была установлена значительная роль плюмов – радиальных столбообразных формирований флюидов диаметром до 100 км – в массоэнергообмене Земного шара. Возник особый термин «плюмтектоника», подчеркивающий их роль в геологических процессах. В настоящее время не вызывает сомнений многообразие вертикальных и горизонтальных перемещений в пределах мантии. Можно предполагать, что они происходят и в ядре. Таким образом, термин «геодинамика», объединяющий комплекс радиальных и тангенциальных движений в недрах Земного шара, оказался весьма удачным. С этих позиций рассматриваются ниже основные геосферы Земли.
Внутренние геосферы Земли
Внутреннее ядро («ядрышко») радиусом около 1200 км является твердым, так как плотность его 11 т/м3 (рис. 94). Даже при том огромном давлении,… Мантия (мощность до 2900 км) состоит, в основном, из силикатов, то есть по…Рис. 94 . Внутренние геосферы Земли
В вопросе об астеносфере остается много неясного.Возможно, она имеет не сплошной, а прерывистый характер.
Нижняя часть верхней мантии характеризуется большей скоростью прохождения сейсмических волн, указывающих на большую плотность вещества с глубиной. До недавнего времени мантия подразделялась на верхнюю и нижнюю мощностью, соответственно, около 640 и 2200 км. В верхней мантии выделялись нижняя и верхняя части, граница между которыми проводилась на уровне 400-410 км. В
последние годы выделена средняя мантия, ограниченная глубинами 840 и 1700 км. Это подразделение характеризуется специфичным минеральным составом, подтверждающим правомерность его выделения. Кроме того, в низах нижней мантии близ её границы с ядром выделяется слой переменной мощности, названный слой D''.
В отличие от ядра вещество мантии выходит на земную поверхность, образуя зоны интенсивного вулканизма, обычно приуроченные к зонам спрединга и субдукции. Однако некоторое количество вулканов находится в центральной части литосферных плит, вдали от их границ. Характерным примером является цепь вулканических Гавайских островов протяженностью около 6000 км, возможно, трассирующая глубинный разлом, достигающий нижней границы нижней мантии (слоя D''). Вулканы Гавайских островов, а также островов Рождества, Реюньона, Исландии рассматриваются как «горячие точки», извергающие высокотемпературную лаву, связанную с веществом нижней мантии. Некоторые «горячие точки» связаны с выходами плюмов, «прожигающих» твердую оболочку Земного шара. Сейчас считается, что по составу нижняя мантия представлена минералами, содержащими O, Si, Mg, Fe, и в значительно меньших количествах – Ca, Al, Na, K. Лабораторные исследования минералов при высоких давлениях показали, что ион Fe2+ в таких условиях значительно уменьшился в объеме, что привело к изменению всех свойств содержащего его минерала. Очевидно, что изменения вещества в условиях гигантских давлений и температур затрагивает его структуру, а не химический состав. Это видно на примере графита, который в определенных условиях переходит в алмаз.
В ходе геологического развития Земли состав мантии существенно изменился на архейско-протерозойской границе за счет увеличения в её составе сиалической составляющей и, вероятно, неоднократно менялся в дальнейшем.
Литосфера– твердая оболочкаЗемли (земная кора и часть верхней мантии) сложенная горными породами. По латерали в составе твердой оболочки выделяются три области: континенты, океанические впадины и приходная зона – геосинклиналь, начинающая развитие в океанической впадине, а заканчивающая – в пределах континента. Процесс образования твердой оболочки происходит следующим образом. В зоне спрединга (рис. 93, 2) в результате поднятия вещества мантии образуется «базальтовый» слой земной коры океанического типа (рис. 93, 3). Верхняя его часть сложена базальтами (мощность 1-3 км), ниже залегают интрузивные породы основного и ультраосновного состава. Во время движения литосферной плиты к окраинам океанической впадины на ней накапливается осадочный слой, обогащенный калием и натрием. В зоне субдукции океаническая литосферная плита погружается под континентальную, на которой в результате аккреции (соскабливания), метаморфизма и частичного расплавления морских осадков образуется гранитный слой (рис. 93, 4). Каждое подразделение твердой оболочки характеризуется специфичным комплексом горных пород – «набором» геологических формаций. Их изучение является предметом формационного анализа. 4.2.2. Земная кора океанического типа
Наиболее достоверные данные получены по формациям земной корыокеанического типа, слагающим океанические впадины. Существенный вклад в их изучение внесли американские ученые, осуществлявшие на протяжении нескольких десятилетий подводное бурение в океанических впадинах. Многоступенчатый процесс формирования океанических впадин можно представить, сведя воедино его стадии, происходящие в различных областях Земного шара. На первом этапе (стадии) развития океанической впадины под воздействием глубинного массоэнергообмена в литосфере возникала зона напряжений, в которой формировались грабенообразные структуры. Их примерами могут служить Рейнский грабен, разделяющий Вогезы и Шварцвальд, впадина Телецкого озера в Горном Алтае, Ферганская долина в Тянь-Шане. Крупнейшим из современных грабенов является впадина озера Байкал. Если напряжение в зоне сохранялось, то формировалась цепь грабенов. Примером является система Великих Африканских озер в Юго-Восточной Африке. На следующем этапе грабены соединялись, образуя узкую глубокую впадину с земной корой океанического типа. В настоящее время примером такой впадины является Красное море, отделяющее Аравийский полуостров от Африканского континента. При дальнейшем расширении грабена Красного моря образуется сооружение типа Мозамбикского пролива, отделяющего от Африки о. Мадагаскар, а затем – океаническая впадина.
По латерали области распространения океанической коры подразделяются на океанические платформы, территориально более или менее совпадающие с ложем Мирового океана, срединно-океанические хребты и зоны трансформных разломов. Океанические платформы (рис.93) по характеру рельефа делятся на котловины и подводные асейсмичные хребты. В котловинах глубина океанов составляет 3-6 км, иногда достигая 7 км. В Атлантическом океане выделяется 7 котловин, в Индийском – 5, в Тихом – 9, в Северном Ледовитом – 4. Рельеф днища котловин определяется мощностью осадочного слоя, меняющейся в широких пределах, в зависимости от притока терригенного материала на единицу площади океанической впадины. В Тихоокеанской впадине, где он сравнительно невелик, развит крупнохолмистый и холмистый подводный рельеф за счет вулканических построек высотой до нескольких сотен метров. Наиболее крупные вулканические постройки, достигающие поверхности океана или даже возвышающиеся над ней в виде островов, получили название подводных гор. В Индоокеанской впадине большая мощность осадочного слоя наблюдается близ устьев крупных рек типа Ганга, где формируются мощные конуса выноса. В силу этого значительное распространение имеет равнинный рельеф океанического дна. Океанические платформы Атлантического океана, характеризующегося еще большим притоком терригенного материала, имеют равнинный рельеф (волнистые и плоские равнины). Наибольшая мощность осадочного слоя наблюдается в Северном Ледовитом океане. Асейсмичные хребты (не путать со срединно-океаническими хребтами) высотой 1-2 км характеризуются линейными очертаниями. Вершины хребтов, возвышающиеся над поверхностью океана, образуют острова (рис. 93, 8).
Осадочный слой океанических платформ подробно описан в главе «Фациальный анализ». Он сложен горизонтально залегающими неметаморфизованными осадками: илами и красной глубоководной глиной. Вследствие движения литосферных плит от зоны спрединга до зоны субдукции в осадочном слое нет пород древнее средней юры (возрастом около 150 млн. лет). «Базальтовый» слой океанических платформ подразделяется на собственно базальтовый и габбро-перидотитовый, видимо, близкий по химико-минералогическому составу подстилающей его мантии. Полезными ископаемыми осадочного слоя океанических платформ являются железомарганцевые конкреции, отлагающиеся на глубинах около 4000 м.
Срединно-океанические хребты – главная на Земле область спрединга (рис. 93, 2) – образуют гигантскую горную систему, протягивающуюся через все океанические впадины и занимающую около 1/6 их суммарной площади. Длина горной системы – около 60 тыс. км, ширина – от 500 до 2000 км, превышение над абиссальной равниной – 3-4 км. В поперечном сечении срединно-океанического
хребта (рис. 95) выделяется четыре зоны: две фланговых, гребневая и осевая. Фланговые зоны шириной в сотни километров характеризуются низкогорным рельефом, обусловленным малой мощностью осадочного слоя. По мере удаления от осевой зоны срединно-океанического хребта, мощность осадков увеличивается, и низкогорный рельеф постепенно переходит в рельеф холмистой равнины. Гребневая зона, разбитая трансформными разломами на узкие (1-10 км шириной) блоки, возвышается над фланговыми зонами на 1,5-2 км. Осевая зона в типичном виде представляет собой рифт – узкую (25-30 км) U-образную крутосклонную депрессию (грабен) глубиной 5-7 км. Она характеризуется мощными излияниями базальтовой лавы, повышенным тепловым потоком из глубин Земли, высокой сейсмичностью, наличием
Рис. 95. Срединно-Атлантический
Хребет
гидротермальных источников. На некоторых участках срединно-океанических хребтов осевая зона выражена не рифтом, а поднятием,
образовавшимся за счет лавы. Описанное выше строение характерно для срединно-океанических хребтов Атлантического, Северного Ледовитого и, отчасти, Индийского океанов. В Тихом океане и на юге Индийского океана зоны спрединга выражены в рельефе не хребтами, а пологосклонными возвышенностями шириной до 1000 км – срединно-океаническими поднятиями (Южно- и Восточно-Тихоокеанское).
Трансформные разломы, субперпендикулярно пересекающие срединно-океанические хребты, смещают в горизонтальном направлении их осевые рифты на десятки и сотни километров (рис. 95).
Застывшая лава осевой зоны срединно-океанических хребтов рассматривается некоторыми исследователями как особая геологическая формация. Её древним аналогом, возможно, являются офиолиты – комплекс магматических пород основного и ультраосновного состава, нередко подстилаемый серпентинитами. Наличие офиолитов в пределах континентов рассматривается как доказательство превращения земной коры океанического типа в континентальную. Осадочный слой в зонах трансформных разломов и срединно-океанических хребтов имеет весьма ограниченное распространение и часто отсутствует. «Базальтовый» слой характеризуется уменьшенной мощностью базальтов, в силу чего его габбро-перидотитовая составляющая подходит близко к поверхности дна. Здесь наблюдаются повышенные значения силы тяжести и усиленный тепловой поток из недр Земли. Некоторые авторы выделяют особый – георифтогенальный – тип земной коры, характерный для срединно-океанических хребтов и зон трансформных разломов.
Земная кора континентального типа
В настоящее время в пределах континентов выделяются континентальные платформы, пассивные окраины континентов, складчатые и глыбовые горы. Каждое… К континентальным платформам с докембрийским фундаментом (древним платформам)… В пределах плиты выделяются синеклизы, антеклизы и авлакогены (рис. 96). Синеклизами называются области понижения…Земная кора геосинклинального типа
Область развития земной коры геосинклинального типа характеризуется крайней расчлененностью рельефа. Вершины вулканических островных дуг или цепи… Структурами земной коры геосинклинального типа являются мио- и… Формирование геосинклинального типа земной коры начинается с момента достижения литосферной плитой зоны субдукции на…Вопросы для самоконтроля
2. Охарактеризовать специфические особенности континентальной и океанической земных кор. 3. Назвать основные типы движения литосферных плит. 4. Назвать основные особенности и главные достижения концепции мобилизма.История развития планеты Земля
5.1. Догеологический этап (стадия) Догеологический этап развития Земли, начавшийся около 4,6 млрд. лет назад в результате взрыва сверхновых звезд,…Архей (AR)
Физико-географические условия архея коренным образом отличались от современных. Климат архея был значительно жарче современного, хотя количество…Рис. 97. Физико-географические условия архея
5.2.1.1. Ранний архей (AR1)
Раннеархейский эон архейского акрона представлен комплексом сильно дислоцированных отложений, высоких степеней метаморфизма, описанных в геологической литературе в качестве «серых гнейсов». По химизму они отвечают магматическим породам среднекислого состава, но, по-видимому, имеют осадочное происхождение. В настоящее время эти породы обнаружены на щитах всех древних платформ.
Распространение пород раннего архея (AR1).Наиболее изучен комплекс «серых гнейсов» на Северо-Американском континенте в пределах Канадского щита: в районе озера Верхнее, на полуострове Лабрадор, в провинции Слайв (район Большого Невольничьего озера), на юго-западе Гренландии (серия Исуа). Нижнеархейские породы провинции Слайв и серии (формации) Исуа считаются древнейшими породами планеты (возраст около 4,1 млрд. лет). На Восточно-Европейской платформе нижнеархейские породы комплекса «серых гнейсов» известны на Балтийском и Украинском щитах. На Балтийском щите они распространены в Карелии и Восточной Финляндии, а также в районе г. Мурманск. На Украинском щите, входившем в состав Южнорусского щита, «серые гнейсы» известны в бассейне среднего течения р. Днепра. На Сибирской платформе к «серым гнейсам» отнесена алданская серия (мощность до 15 км), распространенная в центральной части Алданского щита. Предполагается наличие этих пород и на Анабарском щите. Породы комплекса «серых гнейсов» обнаружены также на Китайско-Корейской платформе и на материках Южного полушария (Африке, Австралии, Антарктиде, Южной Америке), рассматриваемых как «обломки» Гондваны (рис. 98).
«Серые гнейсы», по-видимому, представляют собой реликты древнейшей земной коры континентального (сиалического) типа, выходящей в настоящее время на поверхность в разных районах Земного шара. Действительные границы раннеархейского континента (или континентов) и окружающих океанических впадин остаются неясными, вследствие многократных последующих перемещений литосферных плит. Образование «серых гнейсов», по-видимому, произошло в результате частичной гранитизации еще более древней земной коры океанического типа в ходе саамской складчатости. Таким образом, различия в строении твердой оболочки Земли, возможно, проявлялись уже в раннеархейское время.
Существование в раннем архее гидросферы обосновывается водным происхождением железистых кварцитов (джеспилитов) серии Исуа. Наличие гидросферы подтверждает существование первичной (более плотной, чем в настоящее время) атмосферы, о составе которой имеются разные точки зрения. И.А. Резанов считает, что она, в основном, состояла из водорода. По мнению других ученых в первичной атмосфере преобладали углекислый газ и, возможно, азот, а в качестве примесей присутствовали аммиак, сероводород, пары воды.
Наличие в породах серии Исуа графита, характеризующегося близким к современному соотношением изотопов 13С/12С указывает на его биогенное происхождение, то есть на существование жизни. Присутствие в формации Исуа джеспилитов, формирующихся при чередовании окислительной и восстановительной обстановок, подтверждает это предположение, поскольку в архее свободный кислород образовывался только за счет жизнедеятельности цианоидей. По мнению В.И. Вернадского, возникновение жизни на Земле следует рассматривать как процесс формирования единой системы – биосферы, характеризующейся более или менее равновесным состоянием отдельных её элементов (продуцентов, консументов и редуцентов). В этой связи можно предполагать, что жизнь является одной из форм существования материи и извечно существует во Вселенной (гипотеза панспермии). Согласно этой гипотезе, жизнь на Земле появилась, когда возникли более или менее благоприятные условия. В настоящее время достоверные следы жизни в космосе не обнаружены, и гипотеза панспермии не подтверждена фактическим материалом.
5.2.2.2. Поздний архей (AR2)
В позднеархейское время (3,5-2,5 млрд. лет), по-видимому, произошло растяжение (спрединг) и последующее дробление раннеархейской земной коры континентального (сиалического) типа, представленной «серыми гнейсами». Образовались впадины с океанической (симатической) земной корой. В них накапливались вулканиты базальтового состава и метаосадочные породы, в том числе, железистые кварциты. В дальнейшем в результате метаморфизма и интенсивных дислокаций этих пород образовались зеленокаменные пояса, пронизанные плутонами гранитов. Зеленокаменные пояса достигают в длину 1000 км при ширине до 200 км, причем их современные границы существенно отличаются от первоначальных. Позднеархейские зеленокаменные пояса распространены на всех щитах древних платформ, занимая вместе с массивами «серых гнейсов» значительную часть их территории (рис. 98) .
 Рис. 98. Распределение пород архея и раннего протерозоя в составе континентальной земной коры
Рис. 98. Распределение пород архея и раннего протерозоя в составе континентальной земной коры
В Южном полушарии (юг Африки, королевство Свазиленд) наиболее изучен зеленокаменный пояс Барбертон. Суммарная мощность зеленокаменных отложений, объединяемых в надсерию Свазиленд (стратотип среднего и верхнего архея), близка к 15 км. В силу высокой дислоцированности надсерии, эта цифра является приблизительной. Основание надсерии сложено вулканитами основного и ультраосновного состава. Подушечная отдельность лав, вероятно, образовавшихся из вещества мантии, говорит о подводном характере вулканизма. Стратиграфически выше залегает толща вулканитов разного состава (от базальтов до риолитов), содержащая горизонты кремнистых пород. Она согласно перекрывается слабометаморфизованной толщей туфов, сланцев, железистых кварцитов. Еще выше, несогласно, с конгломератами в основании, залегает грубообломочная терригенная толща, описанная в качестве надгруппы Модис. Эти отложения, уже являются протоплатформенным образованием. Зеленокаменные породы позднеархейского возраста известны также на западе Австралийского континента (блоки Иилгарн и Пилбара), на территории Индостана и Антарктиды.
Значительное распространение они имеют и на щитах платформ Северного полушария. На Балтийском щите Восточно-Европейской платформы зеленокаменные средне-верхнеархейские породы описаны в качестве лопия (мощность 3-5 км). Они обнажаются между Ладожским и Онежским озерами, протягиваясь на север до Белого моря, известны на территории Карелии и Финляндии. Преобладают осадочные породы: песчаники с горизонтами конгломератов, кремнистые и графитистые сланцы, мраморизованные доломиты, железистые кварциты (железорудное месторождение Костомукша). Породы сильно дислоцированы и подверглись метаморфизму амфиболитовой стадии. Граниты, прорывающие лопий, имеют возраст 2,7 млрд. лет. В последнее время к позднему архею относят и вышележащие отложения, описанные в качестве сумия. Это – слабодислоцированная вулканогенно-осадочная толща (мощность 2-4 км) несогласно, с конгломератами в основании, перекрывающая лопий. Метаморфизм пород не превышает зеленосланцевой стадии. Накопление сумия произошло в условиях протоплатформенного режима.
На Украинском щите, по-видимому, входившем в состав Южнорусского щита, средне-верхнеархейские зеленокаменные толщи развиты в пределах Среднеприднепровского блока, где они перекрывают «серые гнейсы». В низах разреза преобладают вулканиты, в верхах – метаморфизованные осадочные породы. Общая мощность средне-верхнеархейских толщ близка к 7 км.
На Алданском щите Сибирской платформы средне-верхнеархейские толщи описаны в качестве субганского комплекса (мощность около 7 км), перекрывающего «серые гнейсы». Низы его сложены основными вулканитами, средняя часть – мраморизованными доломитами. Верхи, представленные удоканской серией, сложены железистыми кварцитами и терригенными породами широкого гранулометрического спектра. Они принадлежат к отложениям протоплатформенного этапа.
На Канадском щите Северо-Американской платформы наиболее крупный пояс зеленокаменных пород (Абитиби) развит в районе озера Верхнего. Еще несколько зеленокаменных поясов располагается в районе Большого Невольничьего озера (провинция Слайв). Мощность слагающих их отложений варьирует от 0,5 до 12 км.
Анализ разрезов верхнеархейских отложений Южного и Северного полушарий показывает, что низы отложений этого хроностратиграфического интервала сложены породами «базальтового» слоя, а верхи – осадочного, причем и те и другие прорваны массивами гранитов. Такая же последовательность перекрывающих отложений характерна для современной геосинклинальной (переходной) зоны. Верхнеархейские породы отличаются от собственно геосинклинальных отложений большей степенью метаморфизма, но в целом аналогия сохраняется. Некоторые исследователи отождествляют низы разреза зеленокаменных поясов с офиолитами.
Органический мир позднего архея был представлен прокариотами – бактериями и цианобионтами (рис. 1. Часть I), следами жизнедеятельности которых являются остатки хлорофилла, достоверно доказанные в породах этого возраста. К полезным ископаемым среднего и позднего архея, кроме железистых кварцитов, принадлежат месторождения лития, бериллия, золота, хрома, никеля и кобальта.
Протерозой (PR)
Протерозойский акрон (акротема) подразделяется на ранне- и позднепротерозойский эоны (эонотемы). Поздний протерозой в свою очередь делится на рифей… 5.2.2.1. Ранний протерозой (PR1)Рис. 99. Джеспилит – железистый кварцит
Первый в истории Земли экологический кризис, кардинально изменивший органический мир произошел около 2 млрд. лет назад. Благодаря гравитационной дифференциации вещества железо сконцентрировалось в ядре, и накопление джеспилитов прекратилось. Свободный кислород начал поступать в атмосферу и постепенно количество его достигло точки Пастера (1% от современного содержания). По выражению А.Г. Заварзина, на рубеже раннего и позднего протерозоя атмосфера «вывернулась наизнанку»: вместо кислородных «карманов», образовывавшихся вокруг цианобионтных матов, появились анаэробные «карманы» в местах разложения органики. Уменьшилась роль прокариот, а господствующее положение позднее заняли эукариоты. Раньше считалось, что смена восстановительной обстановки на окислительную произошла в течение рифея, когда широкое распространение получили терригенные красноцветные породы, накапливавшиеся на континентах в аэробных условиях. Новейшие исследования показали, что смена обстановок произошла не позднее ранне-позднепротерозойского рубежа.
Климат раннего протерозоя по оценкам американских ученых был значительно теплее современного. Среднегодовая температура Земли была близка к 60ºС (в настоящее время она приближается к 15ºС). Также как и в настоящее время наблюдалась климатическая зональность. Во второй половине раннего протерозоя произошло оледенение, имевшее, судя по новейшим данным, глобальный характер. Следы раннепротерозойского оледенения обнаружены на юге Африки, на Индостане, в Австралии, на Канадском и Балтийском щитах, в Юго-Восточной Азии. Возможными причинами оледенения считаются расположение континентов вблизи Южного полюса, воздействие космоса и другие. В число причин оледенения следует включить и понижение содержания углекислого газа в конце раннего протерозоя, вызвавшее усиление теплоотдачи планеты. Наряду с зонами оледенения выделялись области аридного климата, о существовании которых свидетельствуют эвапориты, известные на Канадском и Алданском щитах.
Главным полезным ископаемым раннего протерозоя являются железистые кварциты (джеспилиты), которые в XXI веке могут оказаться основным источником железа. Крупнейшими районами добычи железистых кварцитов являются Криворожье, Курская магнитная аномалия, кряж Каражос в бассейне р. Амазонки, Западная Австралия, Канадский щит. В нижнепротерозойских отложениях Бразилии, Южной Африки, Канады известны месторождения марганца, кобальта, ванадия, титана и хрома. Крупные запасы золота и урана заключены в нижнепротерозойских конгломератах Южной Африки. Медистые песчаники удоканской серии Сибирской платформы содержат запасы меди.
5.2.2.2. Поздний протерозой (PR2)
Согласно принятой в России Общей стратиграфической шкалы позднепротерозойский эон (1,65-0,542 млрд. лет) подразделяется на рифей и венд (продолжительность, соответственно, около 1,0 и около 0,09 млрд. лет). Многие исследователи относят венд к фанерозою, помещая его в составе палеозойской эры ниже кембрия. В зарубежных стратиграфических схемах ранний и средний рифей фигурируют в качестве среднего протерозоя, а к позднему протерозою относят поздний рифей и венд. В данной работе приняты подразделения Общей стратиграфической шкалы. Существенные изменения на рубеже раннего и позднего протерозоя рассматриваются как проявления карельской складчатости. Она вызвала смену протоплатформенного режима на платформенный, и обусловила консолидацию ряда подвижных зон.
Рифей (R)
В составе эталонного разреза рифея снизу вверх выделено три серии: бурзянская (R1), юрматинская (R2) и каратавская (R3) общей мощностью 14-15 км.… К началу рифея на Земном шаре четко выделялись платформы и разделявшие их… Органический мир рифея характеризовался появлением эукариот (рис. 1, часть I), обладавших кислородным типом дыхания.…Рис. 100. Образование Родинии (Пангеи-1), 600 млн лет назад
Климат. Широкое распространение в раннем и среднем рифее высокомагнезиальных известняков и хемогенного кремнеобразования указывает на мягкий и очень теплый климат. По оценкам американских геологов, среднегодовая температура в течение рифея составляла 40-50ºС. В конце среднего рифея произошло понижение температуры земной поверхности, фиксированное сокращением накопления карбонатов и прекращением хемогенного кремненакопления. В позднем рифее произошла дифференциация климата: в экваториальной зоне он остался теплым или даже жарким, а в приполярных зонах стал нивальным. Подтверждением являются горизонты тиллитов, указывающие на покровное оледенение в Северном и Южном полушариях. В подвижных (горно-складчатых) поясах сформировалось горно-долинное оледенение.
Полезные ископаемые рифея представлены месторождениями железа, меди, урана, полиметаллов, а также нефтегазоносными толщами. Осадочные месторождения железных руд известны на Южном Урале, Енисейском кряже, Северной Корее, Австралии, Центральной Африке. Месторождения медных и медно-никелевых руд магматогенного происхождения известны в Канаде, США, Центральной и Южной Африке. Крупные месторождения урана обнаружены в Канаде, Австралии, Центральной Африке. Месторождения полиметаллов гидротермального происхождения найдены в США и Канаде. На Урале за счет вторичного обогащения высокомагнезиальных доломитов образовались залежи магнезита. В рифейских отложениях Сибирской платформы обнаружены крупные запасы нефти и газа.
Венд (V)
Органический мир.Для выделения венда большое значение имело открытие Р. Сприггом и М. Глесснером в 40-50-х годах ХХ века так называемой «эдиакарской… По современным представлениям вендская фауна является древнейшей группой… 1. Мягкотелый облик организмов, лишенных раковин и панцирей. В ископаемом состоянии мягкотелые организмы редко…Основные особенности развития Земли в докембрии
В течение докембрия произошли важнейшие события геологической истории Земли: сформировались геосферы Земного шара, зародилась жизнь, появились… В нашей стране докембрий подразделяется на архейский и протерозойский акроны…Вопросы для самоконтроля
2. Осветить гипотезу образования нашей планеты. 3. Назвать причины жаркого климата раннего архея. 4. Перечислить подразделения архея древних платформ.Палеозойская эра (эратема)
Палеозойская эра (табл. 4) имеет в своем составе шесть периодов (кембрий, ордовик, силур, девон, карбон, пермь). Большинство исследователей считают более правомерным делить её на ранний, соответствующий господству морского режима, и поздний, отличающийся преобладанием наземных условий. Это расчленение палеозоя (по три периода в каждом подразделении) принято в настоящей книге.
Ранний палеозой
Органический мир кембрийского периода характеризуется господством двух групп морской скелетной фауны беспозвоночных: трилобитов из типа… На прочие группы органического мира кембрия приходилось около 10% его… Таблица 4Рис. 102. Расположение континентов и океанов в палеозое
Тектонический режим и осадконакопление.К началу кембрийского периода земная кора подразделялась на континентальные платформы и океанические впадины, характеризовавшиеся корой океанического типа. На границах платформ и океанических впадин располагались геосинклинали и геосинклинальные пояса, превращавшиеся после консолидации в горно-складчатые сооружения (рис. 102). Тектоническая жизнь характеризовалась распространением во второй половине кембрия салаирской фазы каледонской складчатости, в результате которой к концу периода активность большинства геосинклинальных зон существенно снизилась.
ВСеверном полушарии наиболее полные разрезы кембрия Восточно-Европейской платформы вскрываются по берегам Финского залива. В районе Санкт-Петербурга нижний кембрий представлен балтийской серией, сложенной, в основном, синей глиной (мощность100-150 м). Это – отложения мелководного моря, практически лишенные органических остатков и содержащие кристаллики пирита, указывающие на сероводородное заражение бассейна. Среднекембрийские отложения распространены на Восточно-Европейской платформе слабо. Во второй половине кембрия платформа стала областью преимущественной денудации. Осадконакопление продолжалось только на её северо-западной окраине (район г. Осло), где мощность средне-верхнекембрийских отложений достигает 75 м.
Несравненно более широкое распространение имеют кембрийские осадки Сибирской платформы, отсутствующие лишь на территории щитов. Суммарная мощность кембрийских отложений достигает 2-3 км, причем они содержат обильную и разнообразную ископаемую фауну. По составу и строению кембрийских толщ выделяют три типа разреза. На востоке платформы (бассейн среднего течения рр. Лены и Алдана) развиты отложения нижнего и среднего отделов кембрия, представленные, в основном, органогенными известняками, общей мощностью 1,5-2 км. Известняки изобилуют остатками археоциат и трилобитов. На северо-западе платформы (окраины Анабарского щита) кембрийские отложения представлены тремя отделами, а их мощность варьирует от 1,2 до 2,5 км. Наряду с известняками здесь заметную роль играют глинисто-карбонатные, реже существенно глинистые породы. В составе органических остатков преобладают трилобиты; реже встречаются беззамковые брахиоподы и археоциаты. На юго-западе платформы развит третий тип разреза, также представленный тремя отделами кембрийской системы. Этот тип разреза отличается преобладанием лагунного режима. Нижний отдел характеризуется значительным распространением соленосных толщ (Лено-Вилюйский солеродный бассейн). Мощность прослоев и пластов каменной соли варьирует от нескольких сантиметров до 75 м, а суммарная мощность кембрийских отложений достигает 2-3 км.
НаКитайской платформе морские отложения кембрия занимают около 2/3 её площади. Максимум трансгрессии пришелся на вторую половину кембрия. В это время накапливались карбонатные илы, в различной мере содержащие примесь терригенного материала. В начале трансгрессии происходило накопление фосфоритов.
На Северо-Американской платформе в первой половине кембрийского периода преобладали процессы денудации, а в конце среднего кембрия произошла трансгрессия. Море проникло в западные и южные районы, где началось накопление терригенно-карбонатных толщ. В позднем кембрии областью осадконакопления стала большая часть платформы. Суммарная мощность кембрийских отложений близка к 300 м.
В Южном полушарии суперконтинент Гондвана в кембрийское время, в основном, являлся областью денудации, а морские трансгрессии распространялись лишь на его окраины. В бассейне р. Амазонки и в пределах Сахары накапливались существенно грубообломочные отложения мощностью 400-600 м. На севере полуострова Индостан, в Соляном кряже распространены соленосные отложения мощностью около 500 м. Восточная окраина Австралийского континента являлась областью накопления морских терригенно-известняковых отложений.
Крупнейшими океанами в кембрийском периоде, располагавшимися во впадинах с земной корой океанического типа, были Япетус, Палеоазиатский, Палеотетис, Палеопацифика и, возможно, Арктический. Япетусв какой-то мере соответствовал Северной Атлантике. Палеоазиатский океан, разделявший Восточно-Европейскую, Сибирскую и Китайскую платформы, протягивался от Северного Урала через Казахстан и Алтае-Саянскую складчатую область до Забайкалья и Приамурья. Палеотетис отделял Гондвану от платформ Северного полушария. Палеопацифика более или менее соответствовал Тихому океану, а Арктический – Северному Ледовитому.
На границах океанических впадин и континентов располагались геосинклинали и геосинклинальные пояса, превратившиеся впоследствии в горно-складчатые сооружения (рис. 102). Вдоль границы впадины палеоокеана Япетус с Северо-Американской платформой, в пределах Северо-Атлантического геосинклинального пояса, располагались Аппалачская и Иннуитская геосинклинали. В Аппалачской геосинклинали выделялись области мио- и эвгеосинклинального режима, в Иннуитской – большее распространение имел миогеосинклинальный режим. От юго-восточной окраины Гренландии до Северной Норвегии и Шпицбергена протягивалась Грампианская геосинклиналь. В её пределах также преобладал миогеосинклинальный режим, характеризовавшийся накоплением преимущественно известняково-терригенных толщ мощностью 2-7 км.
Миогеосинклинальным режимом характеризовался Средиземноморскийгеосинклинальный пояс, расположенный в пределах Палеотетиса и территориально соответствовавший Альпийско-Гималайскому складчатому поясу. Во впадине Палеоазиатского океана значительное распространение имел Урало-Монгольский геосинклинальный пояс, в пределах которого интенсивным осадконакоплением характеризовались хребты Чингиз (Чингизтау) и Тарбагатай. В раннем и среднем кембрии здесь в условиях эвгеосинклинального режима накапливались терригенно-вулканогенные толщи общей мощностью 4000-5000 м. Восточнее, в пределах Алтае-Саянской складчатой области (АССО) в кембрийском периоде выделялись тектонически стабильные области – срединные массивы – и разделявшие их прогибы, характеризовавшиеся эвгеосинклинальным режимом. Примером прогибов являлась Западная Тува, отличавшаяся в первой половине кембрия исключительно высокой тектонической активностью. В результате салаирской фазы складчатости эвгеосинклинальный режим в АССО в значительной мере прекратил существование, и в средне-позднекембрийское время имел место перерыв в осадконакоплении. На кембро-ордовикском рубеже осадконакопление в Западной Туве возобновилось и в условиях миогеосинклинального режима продолжалось до начала раннего девона. За это время накопились известняково-терригенные толщи мощностью до 10-11 км.
На восточной окраине Сибирской и западной окраине Американских платформ располагалось Тихоокеанское геосинклинальное кольцо, подразделявшееся на Западно-Тихоокеанский и Восточно-Тихоокеанский геосинклинальные пояса. В их пределах выделялись области мио- и эвгеосинклинального режимов. Миогеосинклинальный режим был развит в обоих геосинклинальных поясах. В Западно-Тихоокеанском поясе он известен во Вьетнаме и Лаосе, а также в Российском Приморье. В Восточно-Тихоокеанском поясе он был распространен в Северных Кордильерах. Эвгеосинклинальный режим в Западно-Тихоокеанском поясе был развит на Шантарских островах, Тасмании и Новой Зеландии. В Восточно-Тихоокеанском поясе эвгеосинклинальные отложения распространены на Тихоокеанском побережье Северной Америки, где их мощность достигает 8000 м.
Климат.Кембрийский период характеризовался господством экваториального и тропического климата. Образовывались коры выветривания каолинитово-гидрослюдистого типа с месторождениями марганца и железа. Зоны аридного климата выделяются накоплением соленосных и гипсо-ангидритовых отложений.
Полезные ископаемые. В раннем кембрии главную роль играют фосфориты и каменная соль. Фосфоритоносные бассейны находятся в Каратау (Средняя Азия), в провинции Юнань (Юго-Западный Китай), в Северном Вьетнаме. Солеродные бассейны известны на Сибирской платформе (Лено-Вилюйский бассейн) и в Соляном хребте на севере полуострова Индостан. В кембрийских отложениях Алжирской Сахары заключены крупные месторождения нефти. В нижнекембрийских отложениях Кемеровской области обнаружено крупнейшее в России Усинское месторождение марганца, расположенное в Кузнецком Алатау к северо-востоку от г. Междуреченска (открыто профессором Томского политехнического университета К.В. Радугиным). Месторождения железных руд известны на территории Горной Шории и Кузнецкого Алатау.
Ордовикский период (система, О).Название периода дано по древнему племени ордовиков, населявших полуостров Уэллс на западе Великобритании. Первое предложение о выделении этой системы (периода) было сделано английским геологом Лапвортом в 1879 году. На геологических картах России ордовикская система фигурирует с 1951 года. Согласно решению Межведомственного стратиграфического комитета в нашей стране система подразделена на три отдела и шесть ярусов. Международным геологическим конгрессом ордовикская система утверждена в качестве самостоятельного подразделения в 1960 году. За международный стандарт принята зональная граптолитовая шкала Великобритании. По данным на 2008 год начало ордовикского периода датируется 490 млн. лет, конец – 443 млн. лет (продолжительность 57 млн. лет). В конце ордовика произошла таконская фаза каледонской складчатости.
Органический мир ордовика, также как и кембрия, характеризуется господством морской фауны беспозвоночных, населявшей мелководную зону, причем разнообразие её многократно возросло благодаря обширной трансгрессии. По сравнению с кембрием, в составе органического мира обозначилась роль животных с карбонатным скелетом, из которых многие (строматопораты, табуляты, гелиолитиды, ругозы) являлись рифостроителями (см. рис. 30, 37, 38. Часть I). Формирование твёрдых скелетов позволило многим бентосным животным приподниматься над морским дном и кормиться в богатых пищей водах. В первую очередь это были иглокожие - морские пузыри и морские лилии (рис. 61. Часть I). Среди брахиопод господствовали замковые, строившие карбонатную раковину. У ордовикских трилобитов выработалась способность к свертыванию при приближении опасности для защиты лишенной панциря брюшной части тела. Из моллюсков были распространены головоногие с прямой или слабоизогнутой раковиной (орто- и эндоцератиты) – главные хищники морских бассейнов (рис. 50. Часть I). Для стратиграфии ордовика важнейшей группой являются граптолиты – колониальные организмы, характеризующиеся планктонным или бентосным образом жизни. Первые единичные граптолиты появились в конце кембрия, но широкое распространение они получили в ордовикском периоде. Среди них выделяются дендроидные граптолиты (дендроидеи) и граптолоидеи (рис. 64. Часть I). Тогда же появились первые рыбоподобные позвоночные – бесчелюстные, покрытые костным панцирем (рис. 65. Часть I). В конце ордовика по последним данным появились первые хрящевые рыбы. Флора была представлена сине-зелеными, багряными и зелеными водорослями (рис. 5.Часть I).
Тектонический режим и осадконакопление.В начале ордовикского периода на платформах Северного полушарияпозднекембрийская регрессия сменилась обширной трансгрессией. На Восточно-Европейской платформе отложения ордовика изучены по естественным обнажениям и скважинам в трех районах: на южной периферии Балтийского щита (Эстония, Московская синеклиза), на юго-западной окраине платформы (Приднестровье) и на северо-востоке (Большеземельская тундра). Наиболее детально они описаны на территории Эстонии. Здесь в основании ордовика залегает пачка фосфоритоносных оболовых песчаников (мощность 30 м). Вследствие огромного количества фосфорсодержащих раковин оболид (фораминиферы) песчаники разрабатываются для получения фосфоритов. Стратиграфически выше залегают отложения среднего и верхнего ордовика, сложенные известняками (мощность 200-250 м). В среднем ордовике выделяется горизонт горючих сланцев (кукерситов), используемых для получения жидкого топлива. Западнее (в Швеции), в составе ордовикских отложений преобладают черносланцевые породы (граптолитовые сланцы), содержащие ураноносный прослой. По-видимому, здесь находилась наиболее глубоководная зона бассейна. На юго-западной окраине платформы ордовикские отложения имеют терригенно-карбонатный состав.
На Сибирской платформе ордовикские отложения мощностью до нескольких сотен метров развиты по окраинам Тунгусской синеклизы и на юго-западе платформы. Скважинами они прослежены на значительной площади в западной и центральной частях платформы. В составе отложений преобладают карбонатные породы, изобилующие остатками морской фауны (главным образом, брахиопод). На юге платформы в составе ордовикских отложений встречаются фосфориты. На Сибирской платформе ордовикская трансгрессия по размаху уступала раннекембрийской.
На Китайскойплатформе преобладают терригенно-карбонатные отложения нижнего и среднего ордовика, изобилующие остатками морской фауны; в позднем ордовике она стала областью регрессии. На Северо-Американской платформе в составе ордовикских отложений (мощность первые сотни метров) преобладают карбонатные породы. На восточной окраине платформы в них содержится терригенный материал, поступавший с Аппалачей. В середине ордовика Северо-Американская платформа пережила кратковременную регрессию. По размаху ордовикская трансгрессия на Северо-Американской платформе значительно превосходила кембрийскую.
На Гондване трансгрессия получила ограниченное распространение. В южноамериканской части Гондваны карбонатные породы известны на границе с Восточно-Тихоокеанской геосинклиналью и в Амазонской впадине. В Сахаре распространены пески с прослоями галечников и глин; их мощность варьирует от 500-800 м до 2-2,5 км. На территории Австралии устойчивый морской бассейн существовал у границы с Западно-Тихоокеанским геосинклинальным поясом, периодически распространяясь в западном и северо-западном направлениях. В конце ордовика на Гондване сформировалось покровное оледенение, обусловленное её положением вблизи Южного полюса. Антарктида оставалась областью преимущественной денудации.
В ордовикском периоде существовали те же океаны, что и в кембрии (Япетус, Палеоазиатский, Палеотетис, Палеопацифика и, возможно, Арктический). Как и кембрий, ордовик считается талассократическим этапом развития Земли.
На окраинах океанов вдоль границ земной коры континентального и океанического типов формировались геосинклинальные пояса и области. В пределах Аппалачской и Иннуитской геосинклиналей Северо-Атлантическогогеосинклинального пояса выделялись эв- и миогеосинклинальная зоны. В эвгеосинклиналях накапливались вулканогенно-терригенные толщи мощностью до 4 км, в миогеосинклиналях – терригенно-карбонатные отложения значительно меньшей мощности. Таконская фаза каледонской складчатости, проявившаяся в этих геосинклиналях, не обеспечила их окончательную консолидацию. В Грампианскойгеосинклинали низы ордовика представлены существенно вулканогенными отложениями мощностью около 1,2 км. Вверх по разрезу вулканиты постепенно замещаются известняково-терригенными отложениями общей мощностью около 3 км. Суммарная мощность ордовикских отложений полуострова Уэллс на западе Великобритании близка к 4 км. В конце ордовика в результате таконской фазы каледонской складчатости произошло поднятие территории и обмеление палеобассейна.
В Средиземноморском геосинклинальном поясе ордовикские отложения, распространенные сравнительно слабо, представлены миогеосинклинальными толщами, детально изученными в XIX веке на территории Чехии геологом И. Баррандом. Вулканогенные образования, характерные для эвгеосинклинальных областей, распространены в Центральном Французском массиве, в Вогезах, Шварцвальде.
В Урало-Монгольскомгеосинклинальном поясе кембро-ордовикский рубеж был отмечен заложением на байкальском фундаменте Уральской ветви пояса, соответствующей современному Уралу. В его пределах выделялись западная и восточная зоны, характеризующиеся, соответственно, мио- и эвгеосинклинальным режимом. На западном макросклоне Урала накопились терригенно-карбонатные отложения мощностью 500-2000 м. На восточном макросклоне сформировались вулканиты спилитовой формации мощностью несколько километров.
В Казахстанской ветви Урало-Монгольского пояса сформировались две области эвгеосинклинального режима. Одна из них протягивалась от истоков р. Ишим через Джезказган до Северного Тянь-Шаня, образуя в плане дугу выпуклостью на запад. Другая область располагалась к востоку от Караганды (хребты Чингизтау, Тарабагатай, Джунгарский Алатау). Здесь мощность вулканогенно-осадочных ордовикских толщ достигает 10 км. Эвгеосинклинальный режим завершился таконской фазой каледонской складчатости, сопровождавшейся внедрением гранитоидов в зонах разломов.
В Алтае-Саянской складчатой области (АССО), располагавшейся восточнее Казахстана, ордовикские отложения представлены осадочными толщами (флишевой формацией), характеризующими режим, близкий к миогеосинклинальному.
По обоим берегам Палеопацифики располагались Западно- и Восточно-Тихоокеанский геосинклинальные пояса. В Западно-Тихоокеанском геосинклинальном поясе эвгеосинклинальные условия существовали на территории Японии и юго-восточной окраины Китая. Западнее (ближе к Сибирской платформе) располагалась миогеосинклинальная зона; мощные терригенные отложения ордовикского возраста распространены, в частности, на Верхоянском хребте. На южной окраине Западно-Тихоокеанского пояса, в Восточной Австралии, в ордовике накапливались эвгеосинклинальные толщи. В конце ордовика здесь проявилась таконская фаза складчатости. ВВосточно-Тихоокеанскомгеосинклинальном поясе эвгеосинклинальная зона развита на западном побережье Северной Америки. Восточнее (ближе к Северо-Американской платформе) располагалась миогеосинклинальная зона, имевшая более широкое распространение. В её пределах накапливались карбонатные толщи мощностью до 2 км. На Южно-Американском континенте эвгеосинклинальная зона отсутствовала, а в миогеосинклинальной – накапливались терригенные отложения.
Климат.В течение ордовика произошли существенные изменения климата. Раннеордовикское время характеризовалось теплым климатом с некоторым уклоном в аридность. В среднем ордовике произошло похолодание, сопровождавшееся гумидизацией. В позднем ордовике продолжалось похолодание и произошла некоторая аридизация. Во второй половине позднего ордовика началось покровное оледенение Гондваны, возможно обусловленное её перемещением в высокие широты. В самом конце периода, вероятно, вновь имело место потепление, вызвавшее таяние ледников. Не исключено, что климатические изменения, обусловившие колебания уровня Мирового океана, были вызваны падением астероидов.
Важнейшими полезными ископаемыми ордовикского периода являются нефть Канзаса и Оклахомы (США), горючие сланцы Прибалтики, оолитовые железные руды о. Ньюфаундленд, фосфориты оболового горизонта Эстонии и Ленинградской области.
Силурийский период (система, S).Силурийская система установлена в 1835 году английским геологом Р. Мурчисоном в провинции Уэллс на западе Великобритании. В современном объеме силурийская система фигурирует в России с 1951 года, а Международным геологическим конгрессом этот объем был утвержден в 1960 году. Начало силурийского периода датируется цифрой 443 млн. лет назад, окончание – 418 млн. лет, продолжительность 35 млн. лет. В России силурийская система подразделяется на два отдела, а последние – на два яруса каждый. Во второй половине силура произошла позднекаледонская фаза каледонской складчатости.
Органический мир силурийского периода эволюционировал по нескольким направлениям: в составе морской фауны возросла роль рифостроителей (строматопорат, табулят, ругоз, мшанок), увеличилось её разнообразие, активизировалось освоение органическим миром континентальных пространств. За счет появления новых семейств (спирифериды, продуктиды) существенно обновился состав замковых брахиопод, нередко являвшихся породообразующим материалом. Для силурийского периода характерно появление брахиопод со сложно устроенным ручным аппаратом. Усилилась дифференциация иглокожих на криноидей и морских ежей, характеризовавшихся, соответственно, прикрепленным и подвижным образом жизни. Из членистоногих начинают вымирать трилобиты, зато широкое распространение получили гигантские ракоскорпионы – эвриптериды (рис. 103, 3). Некоторые ракообразные приспособились жить на суше (рис. 103, 8). К концу периода возросла роль конодонтов как важнейшей стратиграфической группы. Граптолиты были представлены, в основном, осеносными граптолоидеями. В конце периода началось их вымирание. Широкое распространение имели головоногие моллюски (подклассы Орто- и Эндоцератоидеи). В составе фауны увеличилась роль позвоночных: наряду с бесчелюстными (рис. 103, 7) интенсивно развивались рыбы (рис. 103; 4, 5). Получили распространение внутриконтинентальные опресненные водоемы, населенные брахиоподами, двустворчатыми моллюсками и ракообразными.

Рис. 103. Реконструкция водоёма силурийского периода. 1 – риниофиты (куксония), 2 – примитивные плауновидные, 3 – эвриптерид, 4 – телодонты, 5 – акантоды; панцирные бесчелюстные: 6 – птераспис, 7 – цефаласпис; 8 – первые наземные ракообразные
Во второй половине силура на окраинах водоемов появились первые высшие растения – риниофиты (род Куксония). Особенности строения риниофитов указывают на их полуводный образ жизни (рис. 103, 1). В самом конце силура появляются примитивные плауновидные (рис. 103, 2).
Тектонический режим и осадконакопление.В пределах платформ Северного полушария в раннесилурийское время продолжалась ордовикская трансгрессия, а в конце периода в результате позднекаледонской складчатости началась регрессия, продолжавшаяся в раннем девоне. На Восточно-Европейской платформе силурийские отложения выходят на дневную поверхность в Прибалтике, Приднестровье и на Тиманском кряже. Скважинами они прослежены на территории Польши и Украины, а также в окрестностях Ярославля и Волгограда. Наиболее детально изучен разрез силура Эстонии (мощность 400-500 м). В его составе преобладают известняки, а в самых верхах появляются красноцветные песчаники, указывающие на начало регрессии. На северо-западной окраине платформы (Литва, Польша и Южная Швеция) распространены маломощные тонкообломочные отложения с остатками граптолитов, характерные для глубоководных бассейнов.
На Сибирской платформе силурийские отложения резко несогласно залегают на ордовикских осадках. Нижнесилурийские отложения представлены широким спектром морских осадков от глубоководных граптолитовых сланцев до мелководных доломитов с прослоями гипса и ангидрита. Верхнесилурийские осадки сохранились лишь на северо-западе платформы. В Тунгусской синеклизе они без видимого несогласия перекрываются пестроцветными отложениями нижнего девона. На силурийско-девонском рубеже к Сибирской платформе присоединились восточные окраины Алтае-Саянской складчатой области: Восточный и Западный Саяны, Минусинский и Тувинский межгорные прогибы.
Китайская платформа в силурийском периоде расширилась в южном направлении за счет причленения к ней горно-складчатых сооружений, образовавшихся в результате консолидации сопредельных геосинклиналей. НаСеверо-Американской платформе мощность силурийских отложений варьирует от первых сотен метров до 1,2-1,5 км. В течение большей части периода накапливались карбонатные толщи, в составе которых значительную роль играли рифогенные образования. В конце силура началось накопление соленосных толщ, указывающее на регрессию и аридизацию климата. Солеродные бассейны занимали северную часть платформы: территорию штата Мичиган, Гудзонов залив, окраины Канадского щита.
Центральная часть суперконтинента Южного полушария – Гондваны– в силуре являлась областью денудации, а на окраинах накапливались мощные терригенно-карбонатные толщи. На южноамериканской части суперконтинента мощность терригенно-карбонатных силурийских толщ варьирует от 100 до 800-1200 м. На Африканском континенте в начале силура произошло углубление океанического бассейна: песчаниковые отложения ордовика сменились граптолитовыми сланцами; в конце силура началась регрессия. Австралийская часть Гондваны являлась областью денудации.
Из океанов в начале силура продолжали существовать Япетус, Палеоазиатский, Палеотетис, Палеопацифика, Арктический (?). Таким образом, в первой половине силура продолжался талассократический этап развития Земли, характерный для кембрийско-ордовикского времени. Во второй половине периода соотношение континентов и океанических впадин существенно изменилось. В результате позднекаледонской фазы каледонской складчатости произошла консолидация большей части Северо-Атлантического геосинклинального пояса, сопровождавшаяся отмиранием впадины палеоокеана Япетус. На западной окраине палеоокеана продолжалось развитие Иннуитской и Аппалачской геосинклиналей, подразделявшихся на эв- и миогеосинклинальные прогибы. В конце силура эти геосинклинали испытали консолидацию, но в Аппалачской геосинклинали она носила временный характер.
На восточной окраине Япетуса – в Грампианской геосинклинали – силурийские отложения залегают на подстилающих толщах со структурным несогласием, отличаясь от них значительно меньшей степенью метаморфизма. Накопление силурийских осадков происходило в условиях миогеосинклинального режима. Выделяется два разнофациальных комплекса отложений. Первый комплекс (мощность около 3000 м) представленный терригенно-известняковыми отложениями, характеризовал шельфовую зону. Разрез этих отложений на полуострове Уэллс (юго-запад Великобритании), изобилующий остатками ископаемой фауны (в том числе граптолитами), является стратотипом силурийской системы. Второй комплекс – маломощные граптолитовые сланцы – формировался в глубоководной зоне палеобассейна.
Во второй половине силурийского периода началась консолидация Грампианской геосинклинали, завершившаяся на силурийско-девонском рубеже превращением её в горно-складчатое сооружение. В раннедевонское время образовались горно-складчатые сооружения Ирландии, Великобритании, Скандинавского полуострова и Западного Шпицбергена, оказавшиеся «мостом», соединившим Восточно-Европейскую и Северо-Американскую платформы.
Силурийские отложения Средиземноморского геосинклинального пояса наиболее детально изучены в районе Праги (Чехия), где они согласно перекрываются нижнедевонскими отложениями. Этот разрез принят в качестве международного стандарта силурийско-девонской границы.
В силурийском периоде уменьшились размеры впадины Палеоазиатского океана за счет образования каледонид Северо-Западного Казахстана и Северного Тянь-Шаня. Консолидация началась на ордовикско-силурийском рубеже таконской фазой складчатости и завершилась на силурийско-девонском рубеже позднекаледонской фазой складчатости. В итоге к началу девона Урало-Монгольский (Урало-Охотский) геосинклинальный пояс оказался расчлененным на Уральскую (Урало-Южнотяньшаньскую) и Монголо-Охотскую геосинклинальные области, территориально разобщенные Казахстанско-Северотяньшаньским блоком земной коры континентального типа, иногда называемым Казахстанским макроперешейком.
В составе Уральской геосинклинальной области выделяются западная (миогеосинклинальная) и восточная (эвгеосинклинальная) зоны, территориально соответствующие макросклонам этой горной системы. На западном склоне Урала и на Новой Земле накапливались карбонатные и карбонатно-глинистые осадки (мощность 500-1500 м), изобилующие остатками ископаемой фауны. На восточном склоне отлагались лавы и туфы, чередующиеся с кремнистыми сланцами и известняками суммарной мощностью до 5000 м. На обоих склонах Урала встречаются участки распространения черных глинистых граптолитовых сланцев, возможно, характеризующие глубоководные зоны палеоокеана. Ископаемая фауна западной и восточной частей силурийского палеобассейна была практически одинаковой. Таким образом, Уральский палеобассейн в течение рассматриваемого периода, несмотря на различие палеотектонических режимов, представлял собой единое целое.
В Казахстанской части Урало-Монгольского пояса во второй половине силурийского периода эвгеосинклинальный режим, существовавший с раннего кембрия по ранний силур, завершился позднекаледонской складчатостью, обеспечившей консолидацию этой территории. Казахстанско-Северотяньшаньский блок стал областью земной коры континентального типа, в пределах которого нижне-среднедевонские отложения полого залегают на сильно дислоцированных силурийских породах. По особенностям геологического строения эта территория принадлежит к межгорным прогибам.
По берегам Палеопацифики располагались Западно- и Восточно-Тихоокеанский геосинклинальные пояса. В Западно-Тихоокеанском геосинклинальном поясе силурийские отложения имеют ограниченное распространение, причем миогеосинклинальные отложения распространены больше, чем эвгеосинклинальные, известные только на юго-восточной периферии Сибирской платформы и на территории Японии. Миогеосинклинальные осадки представлены терригенными породами широкого гранулометрического спектра и, отчасти, известняками. Позднекаледонская фаза складчатости, проявившаяся в Юго-Восточном Китае (на территории Катазии), обусловила несогласное залегание среднедевонских красноцветов на подстилающих породах.
В Восточно-Тихоокеанском геосинклинальном поясе эвгеосинклинальные отложения развиты лишь на узкой прибрежной полосе Северо-Американского континента. Далее к востоку располагалась область миогеосинклинального режима. На западном побережье Южной Америки развиты лишь миогеосинклинальные отложения. В силурийских отложениях центральной части Анд встречена толща тиллитов мощностью 140 м, по-видимому, указывающая на существование здесь горного оледенения.
Климат.В силурийском периоде отчетливо выделялись области гумидного и аридного климата. Северный аридный пояс занимал северную часть Северной Америки. Южный аридный пояс охватывал центральную часть Северной Америки, частично Восточно-Европейскую платформу и запад Австралии. Влажный экваториальный климат существовал в пределах Новой Земли, Урала, Центрального Казахстана, Алтае-Саянской складчатой области. Областью холодного климата, возможно, являлись Анды, где обнаружены грубообломочные, похожие на тиллиты, толщи.
Полезные ископаемые силурийского периода представлены на территории США месторождениями нефти, осадочных железных руд, каменной соли. С гранитоидами связаны месторождения золота в Северном Казахстане и Кузнецком Алатау; с ультраосновными интрузиями – месторождения хромита на Урале и асбеста в Канаде.
Основные особенности развития Земли в раннем палеозое
Тектонический режим раннего палеозоя характеризовался наличием платформ (континентов) с земной корой континентального (сиалического) типа и… К раннепалеозойским платформам Северного полушария относятся… Океанами были Япетус, Палеоазиатский, Палеотетис, Палеопацифика, возможно, Арктический. Геосинклинальными поясами,…Вопросы для самоконтроя
2. Укажите, какие особенности строения риниофитов свидетельствуют об их полуводном образе жизни. 3. Перечислите океаны и континенты, существовавшие к началу раннего… 4. Охарактеризуйте динамику климата раннего палеозоя.Поздний палеозой
Девонский период (система, D). Девонская система выделена в 1839 году А. Седжвиком и Р. Мурчисоном в Англии (графство Девоншир). Расчленение на… Органический мир.К началу девонского периода вымерли или в значительной мере… К девонскому периоду относится одно из важнейших событий эволюции органического мира Земли – появление и развитие…Рис. 103. Образование гигантского суперконтинента Пангея-2
 В Северном полушарии последовательно увеличивались размеры Лавренции за счет включения в её состав Сибирской и Китайской платформ, в силу чего к концу пермского периода Лавренция превратилась в Лавразию. На территории Восточно-Европейской платформы в первой половине раннепермской эпохи существовал теплый мелководный морской бассейн, характеризовавшийся нормальной соленостью воды и накоплением карбонатных осадков. Восточнее, на границе с Предуральским прогибом, на сотни километров в субмеридиональном направлении протягивалась система барьерных рифов. В формировании их участвовали строматопораты, мшанки, кораллы, морские лилии. Размеры рифогенных построек варьировали от нескольких метров до сотен метров в высоту при ширине до 2-3 км. Часто они являются ловушками для нефти. Еще восточнее, в пределах Предуральского прогиба, накапливались глубоководные глинисто-
В Северном полушарии последовательно увеличивались размеры Лавренции за счет включения в её состав Сибирской и Китайской платформ, в силу чего к концу пермского периода Лавренция превратилась в Лавразию. На территории Восточно-Европейской платформы в первой половине раннепермской эпохи существовал теплый мелководный морской бассейн, характеризовавшийся нормальной соленостью воды и накоплением карбонатных осадков. Восточнее, на границе с Предуральским прогибом, на сотни километров в субмеридиональном направлении протягивалась система барьерных рифов. В формировании их участвовали строматопораты, мшанки, кораллы, морские лилии. Размеры рифогенных построек варьировали от нескольких метров до сотен метров в высоту при ширине до 2-3 км. Часто они являются ловушками для нефти. Еще восточнее, в пределах Предуральского прогиба, накапливались глубоководные глинисто-
карбонатные илы.
Во второй половине приуральской эпохи на восточной периферии Восточно-Европейской платформы в пределах Предуральского прогиба произошло засолонение бассейна, обусловившее накопление пластов галита и ангидрита. В северной части прогиба (район Соликамска) кроме галита накапливались и калийные соли. Таким образом, на большей части Предуральского прогиба в биармийскую эпоху накапливалась эвапоритовая формация. Севернее, в бассейне р. Печоры климат был достаточно влажный, и происходило накопление угленосной паралической формации. В татарскую эпоху в пределах Предуральского прогиба накапливались красноцветные и пестроцветные породы континентального происхождения с редкими остатками фауны и флоры. Таким образом, в течение пермского периода в пределах Предуральского прогиба и сопредельной части Восточно-Европейской платформы накапливались морские и лагунно-континентальные отложения. Последние в зависимости от типа климата были представлены гипсо-соленосными или угленосными толщами. Суммарная мощность пермских отложений Предуральского прогиба достигает 2500-3000 м. Прогиб формировался в среднекаменноугольно-пермское время, но максимум прогибания пришелся на приуральскую эпоху.
Несколько иная картина наблюдалась в западной части Восточно-Европейской платформы. Здесь в континентальных отложениях татарского отдела залегает глинисто-известняковая пачка, соответствующая кратковременной позднепермской трансгрессии. Вверх по стратиграфической нормали эти отложения вновь сменяются красноцветными песчаниками. В конце пермского периода территория Восточно-Европейской платформы представляла собой огромную песчаную пустыню, обитатели которой населяли оазисы. Один из них был в бассейне Северной Двины, где В.П. Амалицкий обнаружил захоронение парейазавров и иностранцевий.
Территория герцинид Западной Европы в течение пермского периода характеризовалась переходом от орогенного режима к платформенному. В Германской впадине приуральские отложения со структурным несогласием, обусловленным судетской фазой складчатости, залегают на породах раннекаменноугольного возраста. Эти отложения представлены красноцветными, континентальными известняково-вулканогенно-терригенными породами, изобилующими остатками ракообразных, двустворок, рыб и земноводных. В низах толщи заметную роль играют вулканиты основного состава. Мощность приуральских отложений варьирует от нескольких десятков метров на окраинах впадины до 1200 м в её центральной части. Значительная дислоцированность приуральских отложений позволяет считать, что Германская впадина во время их накопления характеризовалась режимом межгорного прогиба.
Татарские отложения Германской впадины отделены от биармийских поверхностью структурного несогласия, хронологически соответствующей заальской фазе герцинской складчатости. Они залегают практически горизонтально и, соответственно, относятся к чехлу Западно-Европейской платформы. В составе верхнепермских отложений Германской впадины последовательно сменяются четыре пачки. В основании залегают конгломераты мощностью в несколько метров. Выше лежит пачка черных тонкослоистых битуминозных аргиллитов примерно такой же мощности со скоплениями сульфидов меди, цинка, серебра (медистые сланцы). Еще выше залегает известняково-доломитовая пачка мощностью до нескольких десятков метров (цехштейн). Разрез верхней перми заканчивается гипсо-соленосной пачкой мощностью несколько сотен метров.
На территории Северо-Американской платформы пермские отложения распространены меньше каменноугольных. Приуральские и биармийские отложения представлены морскими осадками, татарские – лагунными соленосными. В татарское время на платформе устанавливается континентальный режим. Таким образом, в западной части Лавразии (к западу от Урала) в течение пермского периода нарастала регрессия, происходившая в условиях прогрессирующей аридизации климата.
К востоку от Урала располагалось несколько впадин, характеризовавшихся интенсивным угленакоплением (Кузнецкий, Минусинский, Тунгусский угольные бассейны). Детальное описание истории геологического развития Кузнецкого бассейна, как примера таких впадин, приведено ниже.
На территории Тунгусской синеклизы Сибирской платформы во второй половине пермского периода развивается трапповый магматизм, с наибольшей силой проявившийся на пермо-триасовом рубеже («сибирские траппы»). В составе трапповой формации преобладают лавовые тела основного, иногда ультраосновного состава, а также толщи туфов. Мощность отдельных покровов и потоков достигает первых десятков метров, а суммарная мощность трапповой формации – 2000 м. Лавовые покровы наслаиваются, в силу чего возникает ступенчатость рельефа (трапп – по-английски «лестница»). Кроме того, формируются силы, дайки и подводящие каналы. Таким образом, в составе трапповой формации эффузивные и интрузивные отложения генетически объединяются в вулкано-плутонические ассоциации. Некоторые авторы считают пермо-триасовый вулканизм Сибирской платформы причиной массового вымирания органического мира на палеозойско-мезозойском рубеже.
На Китайской платформе во второй половине пермского периода угленакопление сменилось отложением гипсо-соленосных толщ, происходившим в континентальной обстановке при аридном климате. Каледониды Казахстана в течение пермского периода переживали эпоху активизации. На хребте Чингизтау, характеризовавшимся в каменноугольное время субплатформенным режимом, в пермском периоде возобновилась вулканическая деятельность.
В Южном полушарии в пермское время несколько расширилась территория Гондванского суперконтинента за счет причленения к нему герцинид Восточной Австралии и Южной Африки (Капские горы). Большая часть суперконтинента оставалась сушей, а морской режим существовал лишь на окраинах Гондваны, преимущественно в приуральскую00 эпоху. В Африке морской режим существовал на севере Сахары, в Восточной Африке, на Аравийском полуострове и острове Мадагаскар. Кроме того, морские отложения известны на полуострове Индостан. В мезозойскую эру на этой территории образовалась впадина Индийского океана. Кратковременные вторжения моря отмечены в начале перми и в Южной Америке (бассейн р. Парана).
На суше Годваны в пермском периоде накапливалась гондванская серия. В основании её залегают тиллиты, отложившиеся еще в каменноугольном периоде. На территории Австралии формирование ледниковых отложений продолжалось и в начале пермского периода. Вверх по разрезу тиллиты сменяются паралической угленосной толщей, а последняя – лимнической. Отложения гондванской серии нередко содержат следы дождевых капель, изобилуют остатками пресноводных животных и наземных растений. Сходство этих отложений, распространенных на разобщенных морями материках, подтверждает предположение, об образовании в пермском периоде единого суперконтинента.
Из геосинклинальных поясов в пермском периоде продолжали развитие Средиземноморский и Тихоокеанское геосинклинальное «кольцо», подразделяющееся на Западно- и Восточно-Тихоокеанский пояса. Средиземноморский геосинклинальный поясзначительно уменьшился в результате судетской и астурийской фаз герцинской складчатости. Оставшаяся область геосинклинального режима обычно описывается в качестве геосинклинали Тетис. Типичные разрезы пермских отложений этой геосинклинальной области располагаются в Карнийских Альпах, Динарах, Сицилии. В Карнийских Альпах низы нижнепермского отдела представлены известняковой толщей мощностью до 600 м. Выше с резким угловым несогласием залегает красноцветная песчаниково-вулканогенно-сланцевая пачка мощностью около 150 м, содержащая растительные остатки. Во второй половине раннепермской эпохи здесь прошли процессы складкообразования, сопровождавшиеся интрузивной деятельностью. Верхнепермские отложения представлены известняково-терригенной толщей мощностью до 600 м, содержащей остатки морской фауны и наземных растений.
На территории Турции, Ирана, Афганистана пермские отложения представлены маломощными известняковыми толщами, накапливавшимися в пределах срединных массивов. На восточной окраине геосинклинали Тетис, в пределах Памиро-Алая, господствовал эвгеосинклинальный режим. Здесь отлагались известняково-вулканогенно-терригенные толщи мощностью до 5000 м. Судя по составу органических остатков, в раннепермское время существовал единый морской бассейн, протягивавшийся от Южной Европы до Памира. В конце перми он распался на несколько водоемов.
В Западно-Тихоокеанском геосинклинальном поясе Тихоокеанского геосинклинального «кольца» эвгеосинклинальный режим в пермском периоде существовал в областях, приближенных к Палеопацифике (Корякское нагорье, Сихотэ-Алинь, Японские острова, Индонезийский архипелаг). Западнее, в пределах Верхоянья, располагалась область миогеосинклинального режима. В Восточно-Тихоокеанском геосинклинальном поясе эвгеосинклинальный режим существовал в прибрежной зоне Северо-Американского континента, где накапливались известняково-вулканогенные толщи мощностью до 3500 м. Восточнее, в миогеосинклинальной зоне, отлагались известняково-терригенные осадки мощностью до 5000 м, нередко залегающие на подстилающих толщах с угловым несогласием. В южно-американской части Восточно-Тихоокеанского пояса в начале пермского периода исчезло горное оледенение Анд. Продолжалось, начавшееся в каменноугольном периоде, накопление преимущественно континентальных толщ молассового облика, представленных известняково-вулканогенно-терригенными отложениями общей мощностью до 3000 м. В конце пермского периода произошла мощная складчатость, сопровождавшаяся внедрением гранитных интрузий.
Климатпермского периода в целом был теплым, хотя в самом начале периода на юго-востоке Австралии существовало покровное оледенение, а на территории Анд – горное. Температура морской воды в Приуралье (по кораллам) составляла 26-28ºС, в Закавказье (по брахиоподам) – 19-21ºС, в Памиро-Алае (по фузулинидам) – 19-24ºС. Климатическая зональность пермского периода характеризовалась наличием экваториального, двух тропических и двух умеренных поясов. Тропические пояса отличались аридностью климата. В западной части северного аридного пояса на границе штатов Оклахома, Нью-Мексика, Канзас и Техас располагался один из крупнейших солеродных бассейнов. В восточной части этого пояса соленакопление происходило в Германской, Днепрово-Донецкой и Прикаспийской впадинах, в Приуралье, Западном Казахстане и Средней Азии. Южный аридный тропический пояс представлен гипсоносными красноцветными отложениями Южной Америки и Северо-Западной Африки.
Экваториальный пояс, находившийся между северным и южным тропическими аридными поясами, включал в себя Центральную Америку, Южную Европу, центральные и юго-восточные районы Китая. Здесь произрастали древовидные папоротники и каламиты, лишенные признаков сезонной цикличности – годовых колец.
Южнее и севернее тропических аридных поясов располагались области умеренного климата с более или менее равномерным увлажнением. В них входила большая часть Северо-Американского континента, кроме его южной окраины. В Евразии этот тип климата господствовал в Казахстане, Монголии, Северном Китае. Во второй половине пермского периода имела место аридизация климата, но существенного изменения климатической зональности не наблюдалось. В целом, пермский период характеризуется геократическим режимом.
Полезные ископаемые. В пермском периоде, отличавшимся широким распространением аридного климата, образовались крупные месторождения солей. Из месторождений калийных солей наиболее крупными являются Верхнекамское в Приуралье, месторождения Прикаспия, Германии, Делавэрского бассейна на юге США. Крупнейшим месторождением поваренной соли является Артемовское на севере Донбасса. Для областей гумидного климата характерным полезным ископаемым является уголь. Огромные запасы каменного угля заключены в пермских отложениях Печорского, Таймырского, Минусинского, Кузнецкого и Тунгусского бассейнов, а также в угольных бассейнах Китая, Индии, Австралии. Пермский возраст имеют некоторые нефтеносные горизонты Волго-Уральской антеклизы и ряда месторождений США. Крупными месторождениями газа являются Шебелинское (Украина), Вуктыльское (Коми), Гронинген (Нидерланды), Хьюготон (США). Среди рудных полезных ископаемых наиболее значимыми являются: месторождения меди (Мансфельд, Германия; Джезказган, Казахстан), олова (Корнуолл, Великобритания), урана (Германия, Центральный Французский массив, ЮАР), ртути (Никитовское месторождение, Украина).
Геологическое развитие территории Кузбасса
На геологической карте Кузнецкий прогиб (Кузбасс) вырисовывается как область распространения угленосных палеозойских и мезозойских толщ, причем… Угленакопление в Кузбассе началось в раннем девоне и продолжалось с перерывами… В течение раннекаменноугольного (турнейско-визейского) этапа Кузбасс являлся частью обширного палеобассейна,…Особенности развитя Земли в позднем палеозое
В составе морской фауны позднего палеозоя утратили господствующую роль трилобиты, граптолиты и некоторые группы головоногих моллюсков. Широкое… Угленакопление стало характерной особенностью позднего палеозоя: в это время… Тектоническая жизнь позднего палеозоя характеризовалась сокращением площади океанической коры при соответствующем…Вопросы для самоконтроля
1. Охарактеризуйте изменения, произошедшие в органическом мире в течение позднего палеозоя. Каковы причины кризиса P-T?
2. Перечислите процессы, обеспечившие почвообразование в позднем палеозое.
3. Укажите, как повлияли процессы углеобразования на состав атмосферы и климата позднего палеозоя.
4. Перечислите структурные этажи Донбасса.
5.Перечислите материки, входившие в состав Лавразии и Гондваны в позднем палеозое.
Мезозойская эра (эратема)
Мезозойская эра длительностью 180 млн. лет состоит из трех периодов: триасового, юрского и мелового (табл. 5) Таблица 5Рис. 104 . Распад гигантских континентов, начавшийся в триасе, продолжался на протяжении всего мезозоя.
Стрелками показаны движения литосферных плит.
Геосинклинальный режим в триасовом периоде существовал в Средиземноморском поясе, значительно сократившимся в размерах, и в пределах «Тихоокеанского кольца», состоящего из Западно- и Восточно-Тихоокеанского поясов. Они характеризовались, в основном, морским режимом.
Средиземноморский геосинклинальный поясв триасовом периоде протягивался в субширотном направлении от юго-западных окраин Европы до Индокитая. Тектоно-седиментационный режим в его пределах был весьма различен: одни области характеризовались эвгеосинклинальным режимом (Альпы, Большой Кавказ), другие – представляли собой срединные массивы (Закавказье), где накапливались карбонатные толщи сравнительно небольшой мощности. В Альпах, возможно, существовало два эвгеосинклинальных прогиба. В Западных Альпах преобладал морской режим, и происходила вулканическая деятельность. В Восточных и Южных Альпах широкое распространение имели гипсо-соленосные толщи. На Большом Кавказе триас представлен морскими терригенно-известняковыми отложениями общей мощностью около 1500 м.
На восточной окраине Средиземноморского геосинклинального пояса триасовые отложения распространены на северо-западе Вьетнама, в бассейне р. Черной. Низы нижнего триаса представлены здесь красноцветной вулканогенно-терригенной толщей. Вверх по разрезу она сменяется существенно известняковыми отложениями, а последние – черносланцевой толщей. Накопление морских отложений, иногда содержащих вулканиты, закончилось в начале позднего триаса. Суммарная мощность геосинклинальных отложений триасового периода приближается к 5500 м. В середине позднетриасовой эпохи в результате индосинийской фазы мезозойской складчатости произошли процессы консолидации. В конце позднетриасовой эпохи осадконакопление возобновилось и продолжалось до середины мелового периода. В условиях межгорного прогиба накопилась вулканогенно-терригенная угленосная толща мощностью более 6000 м.
Западно-Тихоокеанский геосинклинальный пояспротягивался от северо-восточных окраин России до Новой Зеландии и Тасмании. На Северо-Востоке России в его состав входили Яно-Колымская и Анюйско-Чукотская геосинклинали, разделенные Колымским (Омолоно-Колымским) срединным массивом. В Яно-Колымской геосинклинали общая мощность триасовых отложений, представленных мелко-тонкообломчными породами, достигает 7000-7500 м. Отложения изобилуют остатками морской фауны. В Анюйско-Чукотской геосинклинали, отличие от Яно-Колымской, широкое распространение имеют вулканиты. В пределах Колымского срединного массива мощность триасовых отложений, представленных, в основном, известняками, не превышает 1000 м. Вулканиты развиты в окраинных частях срединного массива.
В Восточно-Тихоокеанском геосинклинальном поясеэвгеосинклинальный режим существовал лишь на территории Кордильер, где накапливались многокилометровые вулканогенно-терригенные толщи морских отложений. Восточнее (ближе к платформе) в пределах Скалистых гор располагалась область миогеосинклинального режима, характеризовавшаяся меньшей мощностью отложений и отсутствием вулканитов. Различие в составе бентосной фауны Западно- и Восточно-Тихоокеанского поясов рассматривается как косвенное подтверждение существования в триасовом периоде глубоководного океана Палеопацифика, в какой-то мере соответствующего современному Тихому океану.
Климат. Большинство исследователей триасового периода выделяет в качестве главных областей морского режима Бореальный бассейн и море Тетис, разделенные обширной сушей. В состав Бореального бассейна входила акватория Северного Ледовитого океана и приполярные области (Северо-Восток России, бассейн низовий р. Лены, Таймыр, Шпицберген, Гренландия, Арктическая Канада). В пределах бассейна накапливались вулканогенно-терригенные отложения. Единство Бореального бассейна подтверждается сходством видового состава фауны отдельных его частей. В море Тетис накапливались карбонатные осадки с широким распространением эвапоритов. Средняя температура воды варьировала здесь в пределах 21-26ºС, в то время как в Бореальном бассейне она приближалась к 15ºС. В целом, триасовый период характеризовался теплым климатом, а существовавшая в это время зональность, в основном, обусловливалась степенью влажности.
Предполагается, что по степени влажности в триасовом периоде выделялись четыре зоны: экстрааридная (зона пустынь), умеренно-аридная (зона сухих саванн), переменно-влажная и равномерно-влажная. Пустынные условия преобладали в Северной Америке, Северной Африке, Северной Австралии. В этих регионах распространены красноцветные континентальные отложения, а среди морских осадков – эвапориты и доломиты. Умеренно-аридные условия существовали на севере Северной Америки, в Северной Европе, в Западной и Южной Сибири, Монголии, в Северо-Восточном Китае. Здесь появились засухоустойчивые ассоциации растений, а кое-где (вероятно по долинам рек и озер) – комплексы влаголюбивых растений. Переменно-влажный климат существовал на северо-востоке Лавразии, в Приморье, на востоке Австралии и северо-западе Канады. В пределах этой области формировались сероцветные терригенные толщи и коры выветривания. Равномерно-влажный климат существовал в Центральной Америке, на Аляске, в Канаде, Сахаре и Северо-Восточной Азии. К концу триасового периода температура ещё несколько повысилась.
Полезные ископаемые. Нефтегазоносные месторождения триасового возраста известны в Тимано-Печорской области, Алжирской Сахаре, Арктической Канаде, на Аляске. Запасы углей обнаружены на Урале (Челябинский буроугольный бассейн), в США (Южно-Аппалачский бассейн), Китае, Австралии. Осадочные руды урана обнаружены в штате Колорадо (США). Запасы каменной и калийной солей известны в Альпах. С трапповой формацией Тунгусской синеклизы связаны алмазоносные трубки взрыва, месторождения меди, никеля, кобальта, железных руд, графита. С раннетриасовыми корами выветривания связаны месторождения минеральных красок в Средней Азии и Сибири.
Юрская система (период, J) в современном объеме впервые была выделена немецким естествоиспытателем А. Гумбольдтом в 1822 году в качестве «формации юры». В 1829 году французский геолог А. Броньяр описал «формацию» как самостоятельную систему. В настоящее время юрская система подразделяется на три отдела и 11 ярусов. Нижний отдел системы иногда называют лейасом, средний – доггером, верхний – мальмом. Продолжительность юрского периода составляет около 62 млн. лет (208 - 145,8 млн. лет).
Органический мир. Морская фауна юрского периода характеризовалась широчайшим распространением одной из групп головоногих моллюсков – аммонитов. На основе изучения этой быстро эволюционировавшей группы было выделено более 40 аммонитовых зон, последовательно сменявшихся в течение рассматриваемого периода. Широкое распространение в юрском периоде имела и другая группа головоногих моллюсков – вымерший надотряд Белемниты. Значительно сократилось распространение наутилоидей. Кроме того, были распространены двустворчатые и брюхоногие моллюски, иглокожие (морские лилии и морские ежи), брахиоподы (ринхонеллиды и теребратулиды), шестилучевые кораллы, фораминиферы. Из водных позвоночных широчайшее распространение, как и в триасовом периоде, имели рыбы, ихтиозавры и ластоногие рептилии.
В фауне суши господствовали рептилии, главным образом динозавры. В юрском периоде благодаря мягкому влажному климату произрастала пышная наземная растительность, обеспечивавшая благоприятную кормовую базу для растительноядных динозавров. Некоторые из них достигали 20-30 м в длину при весе 50-60 тонн. Американские палеонтологи предполагают существование сейсмозавра, вес которого мог достигать 100-120 тонн. Динозавры жили колониями и отличались определенной заботой о потомстве. По-видимому, они обладали более высоким уровнем психического развития, нежели это предполагалось раньше и, следовательно, более развитым мозгом. Гигантские размеры динозавров, возможно, обеспечивали постоянную температуру тела, необходимую для нормального функционирования мозга. Кроме того, увеличение размеров тела растительноядных динозавров способствовало защите от хищников. В то же время это вызывало увеличение размеров хищников.
Продолжалось освоение наземными рептилиями воздушного океана, началом которого стало появление в триасовом периоде «летающей ящерицы» – лонгисквамы. В первой половине юры появились первые представители летающих ящеров (птерозавров) – рамфоринхи. Некоторые палеонтологи считают, что наличие у птерозавров шерсти доказывает существование у них терморегуляции. В конце юры появилась еще одна группа птерозавров – птеродактили, освоившие планирующий полёт. В юрском периоде появились представители класса птиц. До недавнего времени предком птиц считался археоптерикс. Сейчас многие исследователи предполагают, что предковой формой птиц являлся протоавис, обитавший в конце триасового периода, а археоптерикс был представителем птицеподобных динозавров.
Дискуссия вокруг археоптерикса, в конечном счете, подтвердила разнообразие направлений эволюции органического мира. Из множества путей эволюции, реализуемых в природе различными группами органического мира, наиболее прогрессивным является тот, который сопровождается кардинальной перестройкой организма. Фаунистическая группа, испытавшая такую перестройку, получает в дальнейшем широкое распространение. Примером являются птицы, возникшие за счет кардинальной перестройки одной из групп рептилий. Прочие, приспособившиеся к полетам группы рептилий (например, лонгисквамы, рамфоринхи, птеродактили) в процессе дальнейшей эволюции вымерли. Вымирание происходило тем быстрее, чем менее значительной была перестройка организма. Лонгисквамы, обладавшие крыльями дополнительно к двум парам конечностей, жили более короткий отрезок геологического времени, чем птерозавры, характеризовавшиеся более глубокой перестройкой организма.
Млекопитающие юрского периода являлись одной из групп наземной фауны, имевшей широкое распространение в течение мезозойской эры. По числу видов мезозойские млекопитающие превосходили рептилий. Юрская флора отличается от триасовой исчезновением кордаитов, «семенных папоротников», каламитов. Господствующее положение заняли представители голосеменных: хвойные, гинкговые, цикадовые.
Тектонический режим и осадконакопление. В юрском периоде платформенный режим существовал в Лавразии и Гондване, однако сухопутная связь между этими суперконтинентами нарушилась, в силу чего данный период считается временем распада Пангеи II. Областями геосинклинального режима являлись Средиземноморский, Западно- и Восточно-Тихоокеанский пояса. Из геосинклинальных поясов в течение юрского периода море неоднократно вторгалось на территорию платформ. Максимум трансгрессии пришелся на позднеюрскую эпоху. В Северном полушарии трансгрессия с особой силой проявилась на Восточно-Европейской, Западно-Европейской и Западно-Сибирской платформах, в Южном полушарии – в Восточной Африке, Западной Австралии и на Мадагаскаре. Наличие позднеюрских морских отложений на территории Гондваны, по-видимому, указывает на начало её распада на отдельные континенты, разделенные впадинами Индийского и Атлантического океанов. Мощность юрских трансгрессий позволяет считать этот период талассократическим этапом развития Земли.
Тектоническая жизнь в юрском периоде была достаточно активной. На платформах, как и в триасовом периоде, продолжался рифтогенез, сопровождавшийся вулканической деятельностью. К рифтогенным структурам принадлежат Датско-Польский авлакоген и грабенообразные впадины Забайкалья. На юрский период пришелся максимум мезозойской (киммерийской) складчатости, начавшейся на восточной окраине Средиземноморского геосинклинального пояса в конце триасового периода. С наибольшей силой она проявилась в позднеюрскую эпоху на территории Кордильер, где описана в качестве невадийской фазы.
В состав Лавразии в юрское время входили разновозрастные платформы Северного полушария. На Восточно-Европейской платформе раннеюрские толщи представлены песчано-глинистыми отложениями континентального происхождения, развитыми на Украине и в Прикаспийской впадине. В среднеюрскую эпоху началась южная трансгрессия со стороны моря Тетис и северная – со стороны Бореального бассейна, территориально соответствовавшего Северному Ледовитому океану. В поздней юре большая часть платформы стала дном моря. На Московской синеклизе верхнеюрские отложения, представленные песчано-глинистыми отложениями с остатками морской фауны (мощность около 90 м), несогласно перекрывают среднекаменноугольные известняки.
На Сибирской платформе морские отложения юрского возраста распространены на северных и восточных окраинах в пределах Енисейско-Хатангского (Хатангского), Лено-Анабарского и Приверхоянского прогибов. На Западно-Сибирской плите нижне-среднеюрские отложения представлены песчано-глинистыми континентальными осадками. В позднеюрскую эпоху в результате трансгрессии Бореального бассейна накопились терригенные отложения, изобилующие остатками морской фауны. В это время Западная Сибирь представляла собой огромный морской залив, ограниченный с трех сторон холмисто-увалистой сушей (Урал, Казахстан, Алтай и большая часть Сибирской платформы). Южнее залива располагалась система межгорных впадин, протягивающаяся от Центрального Казахстана до Северного Китая (Карагандинская, Кузнецкая, Канско-Ачинская, Минусинская, Зейско-Буреинская). Во впадинах происходило угленакопление.
На Западно-Европейской платформе сравнительно полный разрез юрской системы наблюдается в пределах Германской впадины. Здесь представлены все три отдела общей мощностью 450-750 м. В составе нижнеюрских отложений доминируют глинистые сланцы, средне- и верхнеюрский отделы сложены известняками. Накопление этой толщи происходило в морском бассейне при теплом или даже жарком климате. Сравнительно небольшая мощность отложений и слабая дислоцированность указывают на платформенный режим области их накопления. Североамериканская часть Лавразии в юрском периоде, в основном, являлась сушей. Морские отложения, чередующиеся с континентальными, развиты лишь на западе платформы, на окраинах Кордильер.
В конце юры закладываются впадины Атлантического и Индийского океанов. Известняки позднеюрского возраста, залегающие непосредственно на океанических базальтах, обнаружены в Северной Атлантике в районе Багамских островов. Таким образом, оказались разделенными евразиатская и североамериканская части Лавразии. В Южной Атлантике юрские морские отложения обнаружены у берегов Западной Африки. Морские отложения юрского возраста, указывающие на зарождение Индийского океана, обнаружены в пределах «Мозамбикского рукава», у побережья Индостана, в Западной Австралии. Бореальный бассейн юрского возраста более или менее соответствовал Северному Ледовитому океану, но резко отличался от него теплым климатом.
Геосинклинальный режим в юрском периоде существовал в пределах Средиземноморского, Западно- и Восточно-Тихоокеанского поясов.Средиземноморский пояс к началу юрского периода сократился за счет герцинской и индосинийской складчатостей. Областями осадконакопления здесь являлись прогибы и срединные массивы. В прогибах накапливались многокилометровые толщи вулканогенно-терригенных пород с пластами известняков и яшм, в срединных массивах мощность юрских отложений, нередко угленосных, не превышает сотен метров.
Примером прогибов являются Северный и Южный прогибы на северном и южном макросклонах Большого Кавказского хребта. В Северном прогибе нижне-среднеюрские отложения глубоко метаморфизованы; их мощность достигает 12000 м. В Южном прогибе степень метаморфизма меньше, а мощность не превышает 5000 м. Однако в Южном прогибе, в отличие от Северного, среднеюрские отложения представлены не осадочными толщами, а вулканитами. Таким образом, вопрос о том, какой из этих прогибов отвечает признакам эвгеосинклинали, а какой миогеосинклинали остается открытым. Этот пример показывает, что разделение геосинклинальных прогибов на эв- и миогеосинклинали в ряде случаев носит условный характер. Верхнеюрские отложения Северного и Южного прогибов резко несогласно залегают на нижне-среднеюрских, что указывает на проявление на средне-позднеюрском рубеже складчатости. Она, однако, не уничтожила различий тектонического режима Северного и Южного прогибов. Областями геосинклинального режима в юрском периоде оставались также Памир и Тибет.
В Западно-Тихоокеанском геосинклинальном поясе продолжали развитие Яно-Колымская и Анюйско-Чукотская геосинклинали, разделенные Колымским срединным массивом. Вулканизм в это время наблюдался на границе срединного массива и геосинклиналей. В поздней юре произошла консолидация геосинклиналей, сопровождавшаяся внедрением гранитоидов. В это же время на границе с Сибирской платформой заложился Приверхоянский прогиб.
В Восточно-Тихоокеанском геосинклинальном поясе областью эвгеосинклинального режима являлись Западные Кордильеры, где накапливались вулканогенно-терригенные отложения мощностью до 6000 м. В позднеюрскую эпоху здесь произошла невадийская фаза мезозойской складчатости, обусловившая возникновение углового несогласия внутри позднеюрских отложений. В конце юрского периода эвгеосинклинальный режим сохранялся на Береговых хребтах, где накапливались мощные терригенные отложения. Областью миогеосинклинального режима в юрском периоде являлась территория Скалистых гор.
Климат юрского периода, вероятно, был значительно теплее современного: следов континентального оледенения в приполярных областях не обнаружено. Для юрского периода, в некоторой мере условно, намечается пять климатических поясов: экваториально-тропическо-субтропический, два аридных и два умеренных. Раннеюрская эпоха характеризовалась преобладанием на Земном шаре экваториально-тропическо-субтропического климата, фиксированного широким накоплением ископаемых углей и бокситов. Термический экватор в это время, по-видимому, располагался в пределах Средиземноморского геосинклинального пояса. Сравнительно небольшое распространение имели области аридного климата. В дальнейшем, в течение юрского периода происходит аридизация и похолодание климата. В средней юре несколько увеличивается область субтропического климата при соответствующем сокращении области экваториально-тропического. Резко расширяется область умеренного климата. В позднеюрской эпохе климатические пояса несколько смещаются в южном направлении и приобретают черты сходства с современными, но среднегодовые температуры были выше. В целом, климатические изменения в течение юрского периода не были значительными. В этой связи не вполне понятна интенсивная изменчивость юрских аммоноидей.
Полезные ископаемые. Теплый влажный климат юрского периода обеспечивал благоприятные условия для процессов угле- и бокситообразования. В юрских отложениях заключено около 1/6 мировых запасов ископаемых углей. Крупнейшими буроугольными бассейнами являются Канско-Ачинский, Убоганский, Иркутский. Крупные запасы каменного угля содержатся в юрских отложениях Южно-Якутского бассейна, угольных бассейнов Грузии, Китая, Австралии. Месторождения бокситов юрского возраста известны на Урале, в Тургайском прогибе, Средней Азии, Енисейском кряже, Средиземноморье. Крупнейшие месторождения нефти юрского возраста найдены на территории Саудовской Аравии, Предкавказья, Средней Азии, Западной Сибири. Газоносные месторождения обнаружены в Баренцевом море. Крупные месторождения осадочных железных руд известны на юге Западной Сибири, Англо-Парижском и Польско-Германском бассейнах.
Мощная интрузивная деятельность во второй половине юрского периода обеспечила формирование месторождений олова, молибдена, вольфрама, золота в пределах Западно- и Восточно-Тихоокеанского геосинклинального поясов. С юрским вулканизмом связано формирование месторождений марганца на Балканах и в Калифорнии, а также меди – в Закавказье.
Меловая система (период, К)в качестве особого подразделения Международной стратиграфической шкалы была выделена бельгийским ученым О. д’Аллуа в 1822 году в Англо-Парижском бассейне. Свое название она получила по широкому распространению в составе отложений белого писчего мела. Двучленное строение меловой системы утверждено Международным геологическим конгрессом в 1885 году, однако в некоторых европейских государствах она расчленяется на три отдела. Ярусное расчленение системы, в основном, произведено в XIX веке. Согласно современным представлениям каждый из двух отделов расчленен на 6 ярусов. Продолжительность периода, по данным 2008 года, составляет около 80 млн. лет (145-65 млн. лет).
Органический мир. Морская фауна беспозвоночных мелового периода характеризуется широким распространением аммоноидей, белемнитов, двустворчатых и брюхоногих моллюсков, морских ежей, губок, мшанок, шестилучевых кораллов, фораминифер. Среди аммоноидей появляются группы, отличающиеся от ранее существовавших строением раковины и межкамерной перегородки. Получает распространение гигантизм: некоторые виды достигают 2 м. На рубеже раннего и верхнего мела значительно обновляются белемниты. Среди двустворчатых моллюсков господствуют иноцерамы, а в тропических морях – рудисты, образовывавшие рифовые скопления. Шестилучевые кораллы участвуют в рифообразовании вместе с рудистами. Значительное распространение в середине периода получили брюхоногие моллюски. Снижается разнообразие брахиопод, но возрастает разнообразие мшанок. Среди морских ежей выделяются группы правильных и неправильных. Во второй половине мелового периода возрастает разнообразие губок. Важным элементом органического мира морских бассейнов мелового периода были микроскопические водоросли – наннопланктон. Среди морских позвоночных большое распространение получают лучеперые и костистые рыбы. Среди водных рептилий исчезают ихтиозавры, но продолжается развитие плезиозавров. Значительное распространение получают гигантские водные ящерицы – мозозавры, которые обитали в большом диапазоне глубин, в силу чего в их скелетах часто заметны следы кессонной болезни.
В органическом мире суши господствующую роль играли растительноядные и хищные динозавры, нередко достигавшие значительных размеров. Среди растительноядных динозавров наиболее крупными были гадозавры (утконосые динозавры), среди хищников и трупоядных – тиранозавры, вес которых достигал 2 тонн. Во второй половине мелового периода, в связи с появлением покрытосеменных растений, некоторые группы растительноядных динозавров приспособились к питанию семенами и приобрели конвергентное сходство с птицами (клювоголовые рептилии). В конце периода появились змеи и крокодилы, получившие широкое распространение в кайнозойской эре. Млекопитающие в меловом периоде были представлены сумчатыми, населяющими в настоящее время Австралию, Северную и Южную Америку. Птицы мелового периода принадлежали к группам ящерохвостых и веерохвостых. Современные птицы произошли от веерохвостых.
В раннем мелу наземная растительность характеризовалась господством голосеменных, представленных цикадовыми, гинкговыми, беннетиттовыми, хвойными. Во второй половине периода появились и получили широкое распространение покрытосеменные растения, многие из которых сохранились до настоящего времени (дуб, платан, лавр, магнолия). В непосредственной связи с наземной растительностью протекала эволюция насекомых, широко распространенных в меловом периоде.
Мел-палеогеновый рубеж ознаменовался одним из крупнейших вымираний органического мира Земли. По подсчетам палеонтологов вымерло более 100 семейств морских беспозвоночных и примерно такое же количество таксонов наземных животных и растений. Вопрос о причинах периодических вымираний (и, соответственно, обновлений) остается открытым. Большинство исследователей объясняет эти события воздействием космического фактора.
Тектонический режим и осадконакопление. В начале мелового периода имела место небольшая регрессия. По-видимому, она была связана с раннемеловой (колымской) фазой мезозойской складчатости, обусловившей отмирание геосинклинального режима на значительной части Западно-Тихоокеанского пояса. На платформах эта фаза складчатости проявилась глыбовыми движениями в Забайкалье, Монголии, Китае. Трапповый магматизм наблюдался в это время в Южной Америке (бассейн р. Параны) и на Земле Франца-Иосифа. Во второй половине раннего мела регрессия сменяется мощной трансгрессией, в ходе которой окончательно оформились океанические впадины (Атлантическая и Индоокеанская), разделившие Лавразию на Северную Америку и Евразию, а Гондвану – на континенты Южного полушария. Трансгрессия, распространившаяся на большую часть современных континентов, фиксирована накоплением карбонатных толщ, в том числе белого писчего мела. Широкое распространение трасгрессии позволяет считать меловой период талассократическим этапом развития Земли.
В конце мелового периода произошла первая (ларамийская) фаза альпийской складчатости. Трапповый магматизм в это время проявился на плато Декан (полуостров Индостан) и в Западной Африке. Продолжалось развитие океанических впадин, однако связь между континентами (по крайней мере, эпизодическая) бесспорно существовала. Об этом свидетельствует сходство наземных рептилий и млекопитающих разных континентов.
В Северном полушарии в результате распада Лавразии образовались Евразия и Северная Америка. В состав Евразии входило несколько разновозрастных платформ. На Восточно-Европейской платформе в течение раннемеловой эпохи находился меридионально ориентированный пролив, соединявший Бореальный бассейн и океан Тетис. Здесь накапливались тонкообломочные отложения с желваками фосфоритов. В начале позднего мела пролив утратил связь с Бореальным бассейном и превратился в обширный залив океана Тетис, восточная граница которого проходила по меридиану Аральского моря. Монголия и Алтае-Саянская складчатая область являлись сушей, населенной динозаврами. В пределах Западно-Сибирской плиты располагался залив Бореального моря, и накапливались нефтегазоносные толщи мощностью более 2000 м. Морским режимом характеризовалась также территория Хатангского прогиба, а Таймыр, возможно, являлся в это время островом. На востоке Сибирской платформы в пределах Вилюйской синеклизы в меловом периоде в условиях наземного режима сформировалась угленосная толща. За счет разрушения гор Верхояно-Чукотской области Западно-Тихоокеанского пояса в Приверхоянском прогибе накопилась терригенные осадки.
Северо-Американский континент, более или менее соответствовавший Северо-Американской платформе, в начале мелового периода испытывал трансгрессию с юга из Мексиканского залива и с севера – из Бореального бассейна. На юге континента накапливались терригенно-карбонатные отложения, на севере – терригенные. В конце мелового периода в связи с ларамийской фазой альпийской складчатости началась регрессия. Накопилась мощная континентальная угле-нефтегазоносная толща, содержащая отпечатки наземных растений и костей динозавров.
Континенты Южного полушария («обломки» Гондваны) в раннемеловую эпоху, в основном, представляли собой высоко поднятую сушу. Морской режим существовал на востоке Африки, на Мадагаскаре, на западе Индостана, в Австралии. В Центральной Африке в наземных условиях отлагались озерно-болотные и аллювиальные отложения. Позднемеловая трансгрессия, в основном, проявилась в Африке, где образовался пролив, соединивший Северо-Африканский бассейн с Гвинейским заливом Атлантического океана. В пределах Конго и Нигерии в это время отлагались континентальные, иногда угленосные толщи. В Южной Америке (на территории Амазонии) угленосные отложения накапливались в течение позднемеловой эпохи.
К концу мелового периода уже имелись все современные океаны. Палеопацифика превратилась в Тихий океан. Доказательством существования южной части Атлантической впадины является смена вверх по разрезу континентальных отложений нижнего мела морскими осадками на атлантических побережьях Африки и Южной Америки. Распределение глубин в пределах впадин приближалось к современному: мелководные отложения Атлантического и Индийского океанов распространены вблизи континентов, глубоководные – в центральных частях впадин. Большинство исследователей считает, что в раннемеловую эпоху в океанах отсутствовали холодные придонные воды, в силу чего циркуляция океанических вод была слабой. Это приводило к заражению сероводородом глубоководных осадков позднего мела.
В результате образования срединных океанических хребтов часть океанской воды могла быть вытолкнута в зону шельфа. Это повысило базис речной эрозии и, соответственно, уменьшило её интенсивность, благодаря чему сократился твердый сток в океаны. Вместо терригенных илов стали накапливаться карбонатные, превратившиеся затем в белый писчий мел. Повышение уровня Мирового океана, возможно, стало причиной позднемеловой трансгрессии.
В пределах Средиземноморского геосинклинального пояса выделяется три области: Альпийско-Гималайская, Восточно-Азиатская и Индонезийская. Наиболее изучена западная часть Альпийско-Гималайской области. Она подразделяется на осевую зону, характеризовавшуюся существованием цепи эвгеосинклинальных прогибов, и две миогеосинклинальные зоны, расположенные севернее и южнее. В состав осевой зоны входили эвгеосинклинальные прогибы Центральных Альп, Динарид, Понтийских гор, Малого Кавказа. В северную миогеосинклинальную зону входили Пиренеи, Северные Альпы, Карпаты, Горный Крым, Большой Кавказ, Копетдаг. Южная миогеосинклинальная зона включала Северный Атлас, Андалузские горы, Южные Альпы, Восточную Адриатику, южные склоны Восточного Тавра, Загрос.
Примером динамики распределения зон является Кавказско-Крымско-Карпатская область. К осевой зоне принадлежал Малый Кавказ, в состав которого входили ээвгеосинклинальный прогиб и Закавказский срединный массив. В раннемеловую эпоху в эвгеосинклинальном прогибе произошло складкообразование и внедрение гранитоидных интрузий, а во второй половине периода – внедрение магм основного и ультраосновного состава. В пределах срединного массива накапливались маломощные толщи известняков.
В пределах Большого Кавказа продолжали развитие миогеосинклинальные прогибы, заложившиеся в юрском периоде. В Северном прогибе накапливалась терригенно-известняковая толща мощностью более 1100 м, в Южном – флишевая толща мощностью до 5500 м. В позднем мелу увеличилось поднятие, разделявшее эти прогибы, в результате чего они оказались оттесненными, соответственно, в северном и южном направлениях. В конце позднего мела Кавказ стал областью денудации, но затем здесь возобновилось осадконакопление. В Горном Крыму в начале мелового периода установился субплатформенный режим: нижнемеловые отложения (мощность 100 м) несогласно залегают на верхнеюрских песчаниках; верхнемеловые – представлены известняками (мощность 200-400 м). В Восточных Карпатах в миогеосинклинальных условиях накапливались флишевые толщи (мощность до 2000 м).
В Восточно-Азиатской области орогенная фаза развития началась еще в конце триасового периода в результате индосинийской фазы мезозойской складчатости. Этот процесс продолжался и в последующее время. В конце мелового периода здесь в условиях платформенного режима происходило накопление известняков.
Западно-Тихоокеанский геосинклинальный пояс в меловом периоде характеризовался отмиранием геосинклинального режима на территории Верхояно-Чукотской области и большей части Сихотэ-Алиня. Эти регионы превратились в горно-складчатые сооружения, в пределах которых областями осадконакопления (в том числе, угленакопления) оставались межгорные прогибы. Во второй половине раннемеловой эпохи произошла колымская фаза мезозойской складчатости, сопровождавшаяся внедрением гранитоидов. В это время обособился Чукотско-Катазиатский вулканогенный пояс, ограниченный с запада зоной глубинных разломов. Произошло отмирание Приверхоянского передового прогиба.
В Восточно-Тихоокеанском геосинклинальном поясе после невадийской фазы мезозойской складчатости областью горного рельефа стали Центральные Кордильеры. Восточнее располагалась область миогеосинклинального режима, западнее (в пределах Береговых хребтов) – эвгеосинклинального. Во второй половине раннемеловой эпохи вновь произошла фаза складчатости, а в середине позднемеловой – следующая, сопровождавшаяся внедрением на Аляске гранитоидных интрузий. В результате этих фаз областью горного рельефа стала вся территория Кордильер. В конце мелового периода в результате ларамийской фазы альпийской складчатости прекратила существование область миогеосинклинального режима, занимавшая территорию Скалистых гор.
Климат. В начале мелового периода имела место некоторая аридизация климата, а в конце раннемеловой эпохи произошло кратковременное похолодание. В целом, географическая зональность климата была близка к современной, но термический режим отличался более высокими температурами. Также как и в юрском периоде выделялись экваториально-тропический гумидный пояс, два аридных и два умеренных пояса. Экваториально-тропический пояс, возможно, имел более широкое распространение, нежели в настоящее время, поскольку включал в себя значительную часть современных субтропиков. С другой стороны, не исключено, что значительная часть территории экваториально-тропического пояса характеризовалась не гумидным, а аридным климатом. Зонами умеренного климата в меловом периоде являлись области, принадлежащие в настоящее время к нивальному поясу. Для морей северной гумидной зоны характерна бореальная фауна, а для континентов – холоднолюбивые растения и углеобразование, широко проявившееся в Евразии и Северной Америке. Наиболее холодный климат, судя по растительным остаткам, наблюдался в Антарктиде, где средние температуры были близки к 15ºС.
Полезные ископаемые. Важнейшими полезными ископаемыми осадочного происхождения, заключенными в отложениях меловой системы являются угли, железные руды, бокситы, фосфориты, белый писчий мел, нефтегазоносные толщи. В меловых отложениях содержится более 1/5 мировых запасов углей. В России крупнейшими угольными бассейнами мелового возраста являются Ленский и Зырянский. Оолитовые железные руды образуют крупный железорудный бассейн на юго-востоке Западно-Сибирской плиты. Месторождения фосфоритов известны на Восточно-Европейской платформе. Цепь этих месторождений прослеживается также на южном побережье Средиземного моря (от Марокко до Сирии). Месторождения бокситов известны в Тургайском прогибе, на Енисейском кряже и Южном Урале. Месторождения солей обнаружены в Туркменистане и Северной Америке. Нефтегазоносные месторождения известны в Западной Сибири, в Ливии, Кувейте, Нигерии, по берегам Мексиканского залива, в Канаде. С кислыми интрузиями Тихоокеанских геосинклинальных поясов связаны месторождения золота, олова, свинца, вольфрама, сурьмы, ртути. В кимберлитовых трубках мелового возраста заключены месторождения алмазов в Южной Африке и Индии.
Особенности развития Земли в мезозойской эре
Органический мир мезозойской эры, вследствие гиганских обновлений на пермо-триасовом и мел-палеогеновых рубежах, резко отличался от органического… Моря мезозоя характеризовались господством головоногих моллюсков, а суша –… В мезозойской эре продолжался круговорот элемента углерода в оболочках Земного шара, начавшийся в девонском периоде…Кайнозойская эра
В соответствии с решениями Международных геологических конгрессов и Межведомственного стратиграфического комитета России кайнозойская эра делится на… Таблица 6Рис. 105.Схема оледенений четвертичного периода
В настоящее время вопрос о причинах оледенений остается открытым, в силу чего нет ясности и в вопросе об их синхронности. С некоторой долей условности считаются синхронными оледенения Северного полушария: в Альпах, на Восточно-Европейской платформе, в Северной Азии и Северной Америке. В Альпах ледниковые эпохи выделяются в качестве гюнца, минделя, риса и вюрма, на Восточно-Европейской платформе – в качестве раннечетвертичного (варяжского и окского), среднечетвертичного, позднечетвертичного (валдайского) оледенений. Наибольшей площадью характеризовалось среднечетвертичное оледенение, подразделяющееся на днепровскую (максимальную) и московскую стадии. Во время днепровской стадии ледник, двигавшийся двумя языками, достигал широт Днепропетровска и Волгограда. На северо-востоке Восточно-Европейской платформы ледники наступали с Тиманского кряжа, Новой Земли, Полярного Урала. В Западной Европе в это время ледники покрывали Британские острова, спускаясь южнее Берлина и Варшавы.
В Северной Азии оледенение охватывало наиболее обеспеченную осадками территорию от Урала до низовьев р. Лены. Вследствие недостатка влаги его распространение было меньшим, чем в Европе. Максимум оледенения пришелся на средний неоплейстоцен. В связи с малой мощностью снежного покрова широкое распространение получило подземное оледенение – многолетняя мерзлота. В значительной мере она сохранилась до настоящего времени.
На Северо-Американском континенте покровным оледенением было занято около 2/3 территории. Число оледенений оценивается по-разному (от четырех до шести). Ледовый покров состоял из 4 ледников (Гренландский, Лабрадорский, Кьювотинский и Кордильерский), которые в ледниковые эпохи объединялись, а в межледниковья разобщались. Южная часть континента (южнее Великих озер) и его северо-западная окраина были свободны ото льдов. На современный рельеф наибольшее воздействие оказало последнее (висконсинское) оледенение. Кое-где сохранились следы предшествующих оледенений (небрасского, канзасского и наиболее значительного – иллинойского).
Существенное воздействие оказало четвертичное оледенение на перигляциальные области Северного полушария, где в ледниковые эпохи формировались лессы, а в межледниковья – почвенные горизонты. В зависимости от характера эпох (ледниковые или межледниковые) менялась влажность климата внеледниковых областей: в ледниковые эпохи климат был влажным, в межледниковья – теплый семиаридный. В свою очередь, характер климата влиял на уровень воды в древних озерах. Современный уровень Мертвого моря на 400 м ниже уровня Мирового океана, а во время ледниковых эпох он был значительно выше этого уровня.
Резкие климатические колебания усиливали циркуляцию атмосферы, в связи с чем в четвертичном периоде увеличилось количество терригенных осадков, поступающих в Мировой океан в результате эолового переноса. Колебания уровня Мирового океана, достигавшие в четвертичном периоде первых сотен метров, также способствовали усиленному притоку в океаны терригенного материала. В итоге, средняя скорость осадконакопления в Мировом океане в четвертичное время была в несколько раз большей, чем в палеогене. Одновременно увеличилось карбонатонакопление в экваториальной зоне и кремненакопление – в умеренно холодной.
Полезные ископаемые четвертичного периода подразделяются на несколько групп. Из россыпей важны месторождения золота, платины, касситерита, алмазов, изумрудов. Среди осадочных руд сохраняют значение бобовые железные руды озерно-болотного происхождения. Всё большее значение приобретают железомарганцевые конкреции. Из нерудных полезных ископаемых важны стройматериалы, месторождения которых во избежание транспортных расходов необходимо осваивать в каждом районе. Из горючих полезных ископаемых имеют значение залежи торфа. Подземные воды являются главным источником питьевого водоснабжения. Лёд Антарктиды может стать источником пресных вод для засушливых районов Земного шара.
Интенсивное изменение природы продолжается и в настоящее время, но его изучением занимается не историческая геология, а науки, характеризующие современное состояние окружающей среды (география, биология, экология и другие). В последние 1,5-2 столетия важнейшей природопреобразующей силой стала деятельность человека. Современное человечество не может существовать, не изменяя природную среду. Однако эти преобразования не должны переходить определенный рубеж, за которым начинается прогрессирующее разрушение природы, предопределяющее гибель человечества. В настоящее время создается принципиально новая отрасль знаний – учение о взаимоотношении человека с окружающей средой. От его дальнейшего развития и, главное, выполнения рекомендаций, в значительной мере зависит будущее человечества.
Особенности развития Земли в кайнозойской эре
Тектоническая жизнь кайнозойской эры характеризуется продолжением альпийской складчатости, начавшейся в позднем мелу (ларамийская фаза). В… Вопросы для самоконтроля:Заключение
Историческая геология, изучающая историю развития планеты Земля, оперирует гигантскими интервалами времени: миллионами и миллиардами лет. В дальнейшем, когда объектом научных исследований станет возраст звездных систем и галактик, возрастные интервалы увеличатся в десятки и сотни раз. В какой-то мере историческая геология аналогична астрономии, имеющей дело с гигантскими расстояниями (сотни световых лет). Обобщение данных исторической геологии и астрономии позволяет наметить некоторые общие закономерности пространственно-временного изменения окружающей действительности, условно названной материей. По-видимому, наиболее общей закономерностью пространственно-временного развития материи является закон периодичности, проявляющийся сочетанием элементов цикличности и направленности. С этой позиции многократно использованный образ «развития по спирали» в полной мере сохраняет иллюстративную значимость. Формы проявления периодичности весьма разнообразны.
Наиболее ярко периодичность проявилась в развитии живого вещества планеты, характеризовавшегося последовательным увеличением видового разнообразия. В процессе роста разнообразия периодически происходили массовые вымирания, в течение которых число видов резко сокращалось. Затем оно постепенно восстанавливалось на новой основе (за счет появления новых групп органического мира). После этого наступала эпоха расцвета этих групп, а затем эпоха их вымирания. Крупнейшие вымирания на венд-кембрийском, пермо-триасовом и мел-палеогеновом рубежах объясняются по-разному (возможно, имело место сочетание нескольких факторов), но сам факт их существования не подвергается сомнению. Периодически менялся и климат Земли. В истории её развития выделяются эпохи оледенения, в течение которых резко понижалась глобальная температура. При этом уменьшение количества лучистой энергии Солнца подчеркивалось увеличением абляции за счет роста ледников.
Проявлением периодичности тектонических процессов является периодическое возникновение Пангеи. Предполагается, что за время существования Земли области распространения земной коры континентального типа несколько раз объединялись, образуя Пангею. Оболочечное (зонарное) строение Земли также может рассматриваться как одна из форм периодичности, поскольку имеет место скачкообразное изменение плотности вещества на границах оболочек, происходящее на фоне его медленного роста с глубиной. Наконец, проявлением периодичности, по-видимому, является периодическое изменение свойств веществ в результате изменения их атомного строения, положенное в основу периодического закона Менделеева. Кристаллическое строение вещества, вероятно, также может рассматриваться как периодическое чередование в пространстве ионов и атомов.
Представления о периодичности развития Земли отражены в пульсационной гипотезе, сформулированной в 30-е годы ХХ столетия В.А. Обручевым и М.А. Усовым. В последние годы эти представления развивались Е.Е. Милановским. Возможно, на основе пульсационной гипотезы и положительных аспектов геодинамики, будет разработана новая теория, являющаяся очередным приближением к научной Истине.
СПИСОК ЛИТЕРАТУРЫ
1. Бейли Д., Седдон Т. Доисторический мир. – М.: Росмэн, 1995. – 160 с.
2. Белоусов В.В. Тектоника плит и тектонические обобщения // Геотектоника. – 1991. - № 2. – С. 3–12.
3. Верзилин Н.Н. Живое вещество как определяющий фактор развития палеогеографических обстановок и геологических процессов в истории Земли // Современные геологические проблемы учения В.И. Вернадского о биосфере. Труды Ленинградского общества естествоиспытателей. – Л.: Изд-во ЛГУ, 1990. – С. 129–155.
4. Владимирская Е.В., Кагарманов А.Х., Спасский Н.Я. и др. Историческая геология с основами палеонтологии. – Л.: Недра, 1985. – 423 с.
5. Вронский В.А., Войткевич Г.В. Основы палеогеографии. – Учебное пособие. – Ростов-на-Дону: Феникс, 1997. – 570 с.
6. Гор Р. Экологические кризисы в истории Земли // За рубежом. – 1989. – № 38. – С. 20–21.
7. Гумерова Н.В. Историческая геология. – Учебное пособие. – Томск: ИПФ, ТПУ, 2011. 106 с.
8. Добрецов Н.Л. Основы тектоники и геодинамики. Учеб. пособие / Новосиб. гос. ун-т. Новосибирск, 2011. – 492 с.
9. Еськов К.Ю. Удивительная палеонтология: история Земли и жизни на ней / К.Ю. Еськов. – М.: ЭНАС-КНИГА, 2012. – 312 с.
10. Историческая геология. – Учебник для студ. высш. учеб. заведений / Н.В. Короновский, В.Е. Хаин, Н.А. Ясаманов. – 4-е изд., стер. – М.: Издательский центр «Академия», 2008. – 464 с.
11. Каныгин А.В. Автогенетическая концепция эволюции биосферы и проблема глобальных экологических кризисов // Актуальные вопросы геологии и географии Сибири / Материалы научной конференции Томского госуниверситета, 1998. – Т. 1. – С. 221–225.
12. Клив Э. Моллюски. Мир животных. – Минск.: Белфаксиздат, 1996. 72 с.
13. Кныш С.К., Гумерова Н.В., Полиенко А.К. Основы структурной, исторической и региональной геологии. – Учебное пособие. – Томск: ИПФ, ТПУ, 2011. 307 с.
14. Латыпов Ю.Я. Экосистема кораллового рифа // Наука в России, 2008 – № 2. – С. 97-106.
15. Лобье Л. Оазисы на дне океана. – Л.: Гидрометеоиздат, 1990. 156 с.
16. Милановский Е.Е. Глобальная цикличность геологического развития Земли в фанерозое и проблемы великих вымираний //Историческая и региональная геология в системе геологического образования / Под ред. А.Х. Кагарманова. – СПб.: Горный ин-т, 1999. – С. 34–35.
17. Михайлова И.А., Бондаренко О.Б. Палеонтология. Ч. 1 –Учебник. – М.: Изд-во МГУ, 1997. – 448 с.
18. Наугольных С.В. Символ Гондваны // Природа, 2012. – № 12. – С. 49–59.
19. Неручев С.Г. Периодичность крупных геологических и биологических событий фанерозоя. – Геология и геофизика, 1999. Т. 40. – № 4. – С. 493–511.
20. Несис К.Н. Кембрийская революция шла быстрее, чем думали // Природа, 1994. – № 2. – С. 94–95.
21. Океаны. Энциклопедический путеводитель. – М.:ЗАО «Махаон», 2007. – 304 с.
22. Парфенова М.Д. Историческая геология с основами палеонтологии. – Учебное пособие. – Томск: Изд. ТПУ, 1998. – 547 с.
23. Подобина В.М., Ксенева Т.Г. Микропалеонтология. – Учебное пособие. – Томск: Томский государственный университет, 2006. – 316 с.
24. Подобина В.М., Родыгин С.А. Историческая геология. – Учебное пособие. – Томск: Изд. НТЛ, 2000. – 262 с.
25. Расницын А.П. Когда жизнь и не думала умирать // Природа, 2012. – № 9. – С. 39–48.
26. Садил Й., Пешек Л. Планета Земля. – Прага: Артия, 1968. – 191 с.
27. Савина Н.И. Основы и методы стратиграфии. – Учебное пособие. – Томск: Изд. ТГУ, 2002. – 196 с.
28. Соколов Б.С. Очерки становления венда. – М.: КМКЛтд., 1997. – 156 с.
29. Сорохтин О.Г., Ушаков С.А. Глобальная эволюция Земли. – М.: МГУ, 1991. – 445 с.
30. Удивительная планета Земля. Иллюстрированный атлас эволюции. – Франция: ЗАО «Издательский дом Ридерз Дайджест», 2003. – 320 с.
31. Федонкин М.А. Загадки вендской фауны // Природа, 1989. № 8. – С. 59–72.
32. Холодов В.Н. Геология и мифотворчество // Природа, 2012. –
№ 6. – С. 25–44.
33. Чумаков Н.М. Климатический парадокс позднего докембрия // Природа, 1992. – № 4. – С. 34–41.
34. Энциклопедия для детей. Геология. – М.:Аванта+, 2001, 688 с.
35. Якобсон К.Э. Парадоксы венда // Природа, 1993. – № 12. – С.
26–32.
36. Январёв Г.С. Историческая и региональная геология. – Учебное пособие. – М.: Недра, 1993. – 271 с.
37. Ясаманов Н.А. Популярная палеогеография. – М.: Недра, 1985. – 136 с.
38. Spinar Z., Burian Z. Leben in der Urzeit. – Leipzig. Jena. Berlin: Urania–Verlag, 1973. – 228 p.
Указатель рисунков, заимствованных у других авторов
Интернет-ресурсы
Рис. 5 а – http://ru-aqua.ru, рис. 5 б –http:// lib.rus.ec.ru, рис. 12, в – http:// Webattach.mail.yandex.net, рис. 28-34 –https://ru.wikipedia.org/wiki/, рис. 43 – http://ru. wikipedia.org/wiki/, рис. 44, г –http://content.foto.mail.ru/mail/deov-popova/1321, рис. 48 –www.planktonchronicles.org/, рис. 59 –http:// Webattach.mail.yandex.net/,
Список литературы - надо
СОДЕРЖАНИЕ
| ПРЕДИСЛОВИЕ ……………………………………………………………….. | ||
| ВВЕДЕНИЕ………………………………………………………………………. | ||
| ЧАСТЬ I. | ПАЛЕОНТОЛОГИЯ ……………………………………. | |
| 1. | Основные понятия и термины…………………………. | |
| 2. | Современная классификация органического мира…. | |
| 2.1. | Надцарство доядерные организмы. Прокариоты (Procaryota)………………………...………………………... | |
| 2.1.1. | Царство Бактерии (Bacteria)………………………………. | |
| 2.1.2. | Царство Цианобионты (Cyanobionta)…………………….. | |
| 2.2. | Надцарство Ядерные организмы или Эукариоты (Eucaryota)……………………...………………………… | |
| 2.2.1. | Царство Растения (Phyta)…………………………………. | |
| 2.2.1.1. | Подцарство Низшие растения (Thallophyta)… | |
| 2.2.1.2. | Подцарство Высшие растения (Telomophyta)………….... | |
| 2.2.2. | Царство Грибы (Fungi)……………………………………. | |
| 2.2.3. | Царство Животные (Zoa)…...…………………………… | |
| 2.2.3.1. | Подцарство Одноклеточные (Простейшие) (Protozoa).…. | |
| 2.2.3.2. | Подцарство Многоклеточные (Metazoa)………………… | |
| ЧАСТЬ II. | ИСТОРИЧЕСКАЯ ГЕОЛОГИЯ……………………….. | |
| 1. | Общие положения……………………………………… | |
| 1.1. | Историческая геология в системе естествознания ……… | |
| 1.2. | Этапы развития исторической геологии…………………. | |
| 2. | Стратиграфия……………………………………...…… | |
| 2.1. | Методы определения относительного возраста..………... | |
| 2.2. | Методы определения абсолютного возраста..………........ | |
| 2.3. | Стратиграфические и геохронологические подразделения…………………………………………… | |
| 2.4. | Принципы стратиграфии………………………………….. | |
| 3. | Фациальный анализ……………………………………… | |
| 3.1. | Литологический метод фациального анализа……………. | |
| 3.2. | Биономический метод…….……………………………….. | |
| 3.2.1. | Фации нормальноморских бассейнов…..………………… | |
| 3.2.2. | Переходные фации…………………………….………… | |
| 3.2.3. | Континентальные фации….………………………………. | |
| 3.3. | Палеогеографические карты……………………………… | |
| 4. | Тектонический анализ………………………..…………. | |
| 4.1. | Формационный анализ……………………………………. | |
| 4.2. | Современные представления о строении Земли………… | |
| 4.2.1. | Внутренние геосферы Земли……………………………… | |
| 4.2.2. | Земная кора океанического типа…………………………. | |
| 4.2.3. | Земная кора континентального типа…………………… | |
| 4.2.4. | Земная кора геосинклинального типа……………………. | |
| 5. | История развития планеты Земля……………………... | |
| 5.1. | Догеологический этап (стадия)……………………………. | |
| 5.2. | Докембрий……………………………………………………. | |
| 5.2.1. | Архей (AR)……...………………………………………....... | |
| 5.2.1.1. | Ранний архей (AR1)…….………………………………........ | |
| 5.2.2.2. | Поздний архей (AR2)…….………………………………...... | |
| 5.2.2. | Протерозой (PR).……………………………………………. | |
| 5.2.2.1. | Ранний протерозой (PR1)...………………………………… | |
| 5.2.2.2. | Поздний протерозой (PR2).………………………………… | |
| 5.2.2.3. | Рифей (R)…………………...……………………………….. | |
| 5.2.2.4. | Венд (V)….………………………………………………….. | |
| 5.2.3. | Основные особенности развития Земли в докембрии.…… | |
| 5.3. | Фанерозой…………………………………………………… | |
| 5.3.1. | Палеозойская эра (эратема) (PZ)…………………………… | |
| 5.3.1.1. | Ранний палеозой (PZ1)……….…………………………… | |
| 5.3.1.2. | Основные особенности развития Земли в раннем палеозое…………………………………………………….. | |
| 5.3.1.3. | Поздний палеозой (PZ2)……...…………………………… | |
| 5.3.1.4. | Основные особенности развития Земли в позднем палеозое………………………………………………………. | |
| 5.3.2. | Мезозойская эра (эратема) (MZ)………………………....... | |
| 5.3.2.1. | Особенности развития Земли в мезозойской эре…..……… | |
| 5.3.3. | Кайнозойская эра (эратема) (KZ)………………………….. | |
| 5.3.3.1. | Особенности развития Земли в кайнозойской эре……….. | |
| ЗАКЛЮЧЕНИЕ…………………………………………………………………… | ||
| СПИСОК ЛИТЕРАТУРЫ…………………………………………....................... | ||
| УКАЗАТЕЛЬ РИСУНКОВ, ЗАИМСТВОВАННЫХ У ДРУГИХ АВТОРОВ… | ||
| ПРИЛОЖЕНИЯ ………………………………………………………………….. | ||
| СОДЕРЖАНИЕ………………………………………………………………….… |