рефераты конспекты курсовые дипломные лекции шпоры

- Раздел История
- /
- К.Ю. Еськов История Земли и жизни на ней
Реферат Курсовая Конспект
К.Ю. Еськов История Земли и жизни на ней
К.Ю. Еськов История Земли и жизни на ней - раздел История, К.ю. Еськов ...
К.Ю. Еськов
История Земли и жизни на ней
Благодарности Пользуюсь случаем выразить глубокую признательность своим коллегам, геологам и биологам - О.А. Афанасьевой, В.Ю.…Содержание
· Авторское предуведомление ...
· Возраст Земли и Солнечной системы. Абсолютный и относительный возраст. Геохронологическая шкала ...
(дополнительная). Несколько слов о методологии науки. Принцип актуализма, "Бритва Оккама" и презумпции. Проверка теории: верификации и фальсификации ....
· Образование нашей планеты: "холодная" и "горячая" гипотезы. Гравитационная дифференциация недр. Происхождение атмосферы и гидросферы ...
· Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция ...
· Происхождение жизни: абиогенез и панспермия. Гиперцикл. Геохимический подход к проблеме ....
(дополнительная). Термодинамические подходы к сущности жизни. Второе начало термодинамики, энтропия и диссипативные структуры...
· Ранний докембрий: древнейшие следы жизни на Земле. Маты и строматолиты. Прокариотный мир и возникновение эукариотности ...
· Поздний докембрий: возникновение многоклеточности. Гипотеза кислородного контроля. Эдиакарский эксперимент ...
(дополнительная). Взаимоотношения хищника и жертвы в экологическом и эволюционном масштабах времени ...
· Кембрий: "скелетная революция" и пеллетный транспорт. Эволюция морской экосистемы: кембрий, палеозой и современность ...
· Ранний палеозой: "выход жизни на сушу". Появление почв и почвообразователей. Высшие растения и их средообразующая роль. Тетраподизация кистеперых рыб ...
· Поздний палеозой - ранний мезозой: криоэры и термоэры. Палеозойские леса и континентальные водоемы - растения и насекомые ...
· Эволюция наземных позвоночных (1): поздний палеозой - ранний мезозой. Анамнии и амниоты. Две линии амниот - тероморфная и завроморфная ...
· Эволюция наземных позвоночных (2): поздний мезозой. Завроморфный мир. Маммализация териодонтов. Динозавры и их вымирание ...
· Мезозойские биоценотические кризисы. "Ангиоспермизация мира" (средний мел) и "Великое вымирание" (конец мела). Импактные и биотические гипотезы ...
· Кайнозой: наступление криоэры. Новые типы сообществ - тропические леса и травяные биомы. Эволюция млекопитающих и появление человека ...
· Четвертичный период (антропоген): Великое оледенение. Ледниковая теория. Перигляциальные сообщества и мамонтовая фауна ...
(дополнительная). Историческая биогеография. Викариантная модель и концепция "оттесненных реликтов". Фитоспрединг ...
· Рекомендуемая литература
· Словарь терминов
· Словарь названий живых и ископаемых организмов
Авторское предуведомление
Поскольку учебный курс этот предназначен не для "среднестатистического школьника", а для людей, собирающихся связать свою судьбу с наукой,… Эйнштейн как-то заметил (вполне справедливо), что если ученый не в состоянии… Хочу также предупредить, что в ряде случаев я буду излагать факты и обобщения последних лет, которые в принципе могут…Возраст Земли и Солнечной системы. Абсолютный и относительный возраст. Геохронологическая шкала.
Необходимо заметить, что людей первоначально заинтересовал возраст не Земли как небесного тела, а именно обитаемой Земли - как сейчас сказали бы,… Установление же времени существования Солнца - после того как были открыты… Необходимо было найти источник, питающий своей энергией Солнце - иначе вообще рушился закон сохранения энергии. И вот…А (дополнительная). Несколько слов о методологии науки. Принцип актуализма, "Бритва Оккама" и презумпции. Проверка теории: верификации и фальсификации.
Принцип актуализма (термин этот был введен в 1830 году Ч.Лайелем) заключается в том, что при любых реконструкциях событий прошлого мы исходим из того, что в те времена должны были действовать такие же законы природы, что и ныне; сам Лайель кратко формулировал его как "Настоящее есть ключ к прошлому". И пускай, к примеру, в докембрии существовали экосистемы, не имеющие современных аналогов - но камень-то, надо думать, и тогда падал на землю с ускорением 9,8 м/сек2, вода замерзала при нуле градусов Цельсия, а молекула хлорофилла исправно поглощала кванты света... А, собственно говоря, почему? Вопрос этот вовсе не так уж прост.
Непосредственно в прошлое заглянуть невозможно, машина времени - это несбыточная мечта человечества. Любые наши суждения о прошлом есть лишь более или менее вероятные предположения, основанные на интерпретации фактов и событий современности. Динозавры (столь полюбившиеся широкой публике после "Юрского парка") - это, вообще-то говоря, лишь куски песчаника, напоминающие своей формой кости современных рептилий; все же остальное - чистые домыслы. Понятное дело, что цена домыслам режиссера С.Спилберга и академика от палеонтологии Л.П.Татаринова несколько разная, однако экспериментально проверить нельзя ни первые, ни вторые - ни сегодня, ни в будущем. Поэтому для начала нам следует решить для себя принципиальный вопрос: познаваемо ли прошлое вообще? При этом необходимо признать, что на логическом уровне проблема неразрешима, то есть это вопрос не разума, а веры.
Если ответ будет "нет", то мы можем дальше по собственному усмотрению населять прошлое атлантами и лемурийцами, разумными спрутами и крылатыми огнедышащими драконами, а можем, наоборот, отрицать существование всего, что не упомянуто - черным по белому - в Ветхом Завете. Пожалуйста; мы теперь находимся в сфере мифологии, можно ни в чем себе не отказывать. Отправившись по этому пути, мы с неизбежностью должны придти к отрицанию существования Хеопса, Ивана Грозного, а то и товарища Сталина - чем они в этом смысле лучше динозавров?
Если же мы примем, что прошлое принципиально познаваемо (а подавляющее большинство людей решают для себя этот вопрос именно так), и останемся при этом на позициях рационального мышления (то есть будем полагаться не на "откровения свыше", а на свои собственные наблюдения и умозаключения), то упомянутый выше кусок песчаника немедленно превратится в бедренную кость тиранозавра. Структура ее поверхности позволит нам сделать выводы о местах прикрепления мышц, и соответственно, о типе походки, скорости передвижения и возможных способах охоты; внутренняя структура кости - о характере кровоснабжения, и соответственно, о возможной теплокровности этих существ. Ископаемая древесина с годичными кольцами позволит заключить, что климат в этом месте тогда был сезонным, а ископаемый коралловый риф - что температура окружающей его морской воды превышала 20 градусов. Все эти выводы будут основаны на аналогиях - на том, как ведут себя кости позвоночных, древесина и коралловые рифы в наши дни. Но вправе ли мы исходить из такой предпосылки? Не только вправе - мы обязаны поступать именно так, и вот почему.
Мы уже оговорили, что действуем в сфере рационального мышления. Рациональный тип мышления - отнюдь не единственно возможный; бывает мышление художественное, мистическое, религиозное, и т.п. Надо отчетливо осознавать, что ни одно из них не "хуже" и не "лучше" остальных - они просто разные, и имеют свои собственные "своды законов". Мы вольны в выборе типа мышления - но, раз выбрав, обязаны будем в дальнейшем подчиняться определенным правилам.
Одним из фундаментальных принципов рационального мышления является "Бритва Оккама" (по имени английского философа XIII века); сам Оккам формулировал его как "Не умножай сущностей сверх необходимого". Применительно к правилам научного исследования это означает следующее: выбирая одну из нескольких гипотез, объясняющих некое явление, надо начинать с самой простой из них, и только убедившись в том, что она "не работает", переходить к более сложной, повторяя эту процедуру до тех пор, пока не будет найдено простейшее удовлетворительное объяснение.
Приведем такой пример. На тихоокеанском острове Пасхи имеются циклопические статуи, которые, казалось бы, не могли быть воздвигнуты примитивным племенем, населяющим остров в наши дни. Можем ли мы высказать гипотезу, что статуи эти поставлены пришельцами с другой планеты? Конечно, можем. Однако, находясь в рамках рационального подхода, мы вправе принять подобное объяснение лишь после того, как будут исчерпаны все более простые - "земные" - гипотезы. Тур Хейердал, сделавший успешную попытку установить пасхианскую статую с помощью лишь тех средств, что есть в наши дни в распоряжении тамошних аборигенов, действовал строго в рамках "Бритвы Оккама" - хотя наверняка не задумывался над этим. Последнее весьма существенно: дело в том, что принцип "Бритвы Оккама" (и впоследствии развившийся из нее принцип парсимонии) для любого ученого, по крайней мере, в сфере естественных наук, настолько фундаментальны, что обычно его просто не замечают - как мы не замечаем воздуха, которым дышим.
Возвращаясь к методам реконструкции картин далекого прошлого, отметим, что с этой точки зрения актуализм - стремление в исторических реконструкциях отталкиваться от современных аналогов - совершенно корректен. Существование же в прошлом принципильно иных, чем ныне действующие, законов природы будет той самой "избыточной сущностью", которую и отсекает "Бритва Оккама". Собственно говоря, прошлое вообще познаваемо ровно настолько, насколько точные аналогии былым ситуациям существуют в современности. Однако в следующих главах мы регулярно будем сталкиваться и с такими совокупностями фактов, для объяснения которых нам придется предполагать, что в природе существовали и ситуации, ныне совершенно немыслимые, как-то: экосистемы, не имеющие в своем составе хищников; ландшафты, которые не являются ни сушей, ни морем, а чем-то средним; атмосферная циркуляция, при которой число конвективных ячеек отлично от нынешнего. Являются ли такие реконструкции отступлением от принципа актуализма? Нет, не являются, и вот почему.
Дело в том, что принцип актуализма не является аксиоматическим утверждением. Аксиома - это принимаемое без доказательств положение, на основе которого строится внутренне непротиворечивая система взглядов. Если мы принимаем аксиому "Через точку, лежащую вне прямой, можно провести одну и только одну прямую, параллельную данной", то получаем внутренне непротиворечивую геометрию Евклида. А если принять, что через такую точку можно провести несколько прямых не пересекающих данную, то возникнет геометрия Лобачевского, столь же внутренне непротиворечивая, что и "нормальная", евклидова.
Выше мы уже сталкивались с одним аксиоматическим утверждением - законом напластования ("если один слой горных пород лежит на другом, то верхний слой образовался позднее, чем нижний"), на котором основана такая внутренне непротиворечивая система взглядов, как стратиграфия. Поэтому если бы вдруг удалось доказать (напрягите воображение!) что вышележащий слой может образоваться прежде нижележащего - это означало бы полное разрушение картины Мира, что создана стратиграфией.
Принцип же актуализма принадлежит к совершенно иному типу утверждений - презумпциям. Всем известна используемая в юриспруденции презумпция невиновности. Она может быть сформулирована так: поскольку большинство людей не являются преступниками, то каждый отдельно взятый человек должен считаться невиновным до тех пор, пока не доказано обратное. Последнее - чрезвычайно важно: в презумпции изначально заложена возможность опровержения; она лишь устанавливает очередность, в которой следует рассматривать соответствующие гипотезы (применительно к презумции невиновности это означает, что обвиняемый не обязан ничего доказывать - это дело обвинителя).
Палеонтолог А.П.Расницын показал, что этот тип утверждений используется в естественных науках чрезвычайно широко, хотя практически всегда - в неявном виде. Например, постоянно практикуемое биологами определение степени родства организмов по степени их сходства - не что иное как презумпция, которую можно сформулировать так: "Более сходные между собой организмы должны считаться более близко родственными между собой до тех пор, пока не доказано обратное (т.е. конвергентное возникновение этого сходства)". В дальнейшем мы будем часто сталкиваться с этим типом логических конструкций. Одной из презумпций и является принцип актуализма, который может быть переформулирован таким образом: в процессе исторического исследования мы должны исходить из того, что любые системы в прошлом функционировали так же, как их современные аналоги, до тех пор, пока не доказано обратное.
Раз уж зашла речь о научном мышлении, то следует рассказать о взглядах одного из крупнейших философов XX века, математика по базовому образованию - К.Поппера. Он одним из первых задался вопросом: когда теорию можно считать научной? Поппер сразу уточняет: "Меня интересовал не вопрос о том, "когда теория истинна?" [...] Я поставил перед собой другую проблему. Я хотел провести различие между наукой и псевдонаукой, прекрасно зная, что наука часто ошибается, а псевдонаука может случайно натолкнуться на истину". Издавна существует стандартный ответ: наука отличается от псевдонауки (или от "метафизики") своим эмпирическим методом, т.е. исходит из наблюдений и экспериментов. Однако такой ответ вряд ли можно счесть исчерпывающим: например, астрология (которая, очевидным образом, наукой в строгом смысле не является) оперирует громадной массой эмпирического материала, опирающегося на наблюдения - гороскопами и биографиями.
Поппер вспоминает, что в 1919 году, когда он начинал учиться в Венском университете, все были увлечены новыми, поистине революционными, концепциями: теорией относительности Эйнштейна, а также историческим материализмом Маркса и новейшими психологическими теориями - психоанализом Фрейда и так называемой "индивидуальной психологией" Адлера. Быстро ощутив - сперва на каком-то подсознательном уровне - некое принципиальное различие между двумя этими группами теорий, Поппер попытался сформулировать для себя: чем марксизм, психоанализ и индивидуальная психология так отличаются от физических теорий - например, от теории относительности? Ясно, что дело тут было не в математическом аппарате (или отсутствии такового), а в чем-то ином, более серьезном.
"Я обнаружил, что те из моих друзей, которые были поклонниками Маркса, Фрейда и Адлера, находились под впечатлением некоторых моментов, общих для этих теорий, в частности под впечатлением их явной объяснительной силы. Казалось, эти теории способны объяснить буквально все, что происходило в той области, которую они описывали. Изучение любой из них как бы приводило к полному духовному перерождению или к откровению, раскрывающему наши глаза на новые истины, скрытые от непосвященных. Раз ваши глаза однажды были раскрыты, вы будете видеть подтверждающие примеры всюду: мир полон верификациями теории. Все, что происходит, подтверждает ее".
Итак, главная черта этой группы теорий - непрерывный поиск верифицирующих их эмпирических результатов (наблюдений): чем больше, тем лучше. Более того: невозможно представить себе, например, такую форму человеческого поведения, которая не укладывалась бы в рамки соответствующей психологической теории. В примере, рассматриваемом Поппером, один человек толкает ребенка в воду с намерением утопить его, а другой жертвует жизнью в попытке спасти этого ребенка: "Каждый из этих случаев легко объясним и в терминах Фрейда, и в терминах Адлера. Согласно Фрейду, первый человек страдает от подавления некоего комплекса (скажем, Эдипова), а второй достиг сублимации. Согласно Адлеру, первый человек страдает от чувства неполноценности (которое вызывает у него необходимость доказать самому себе, что он способен отважиться на преступление); то же самое происходит и со вторым (у которого возникает потребность доказать самому себе, что он способен спасти ребенка)." С такой же легкостью обе эти теории переинтерпретируют и любые другие человеческие поступки.
С теорией относительности дело обстоит совершенно иначе. Как раз во время, описываемое Поппером, А.Эддингтону впервые удалось подтвердить одно из предсказаний, сделанных Эйнштейном. Согласно его теории гравитации, большие массы (такие, как Солнце) должны притягивать свет точно так же, как они притягивают материальные тела. Поэтому свет далекой фиксированной звезды, видимой вблизи Солнца, достигает Земли по такому направлению, что звезда кажется смещенной по сравнению с ее реальным положением. В обычных условиях этот эффект наблюдать невозможно, поскольку близкие к Солнцу звезды совершенно теряются в его ослепительных лучах. Однако эти звезды можно сфотографировать во время полного солнечного затмения, а затем сравнить их положение с тем, что наблюдается ночью, когда масса Солнца не влияет на распространение их лучей. Именно это и проделал Эддингтон, получив в итоге тот самый эффект, что был ранее предсказан Эйнштейном.
"В рассматриваемом примере, - пишет Поппер, - производит впечатление тот риск, с которым связано подобное предсказание. Если наблюдение показывает, что предсказанный эффект определенно отсутствует, то теория просто-напросто отвергается. Данная теория несовместима с определенными возможными результатами наблюдения - с теми результатами, которых до Эйнштейна ожидал бы каждый. Такая ситуация совершенно отлична от описанной мною ранее, когда соответствующие [психологические] теории оказывались совместимыми с любым человеческим поведением, и было практически невозможно описать какую-либо форму человеческого поведения, которая не была бы подтверждением этих теорий."
Все это и привело Поппера к заключению о том, что подтверждения (верификации) теории недорого стоят - их при желании можно набрать сколько угодно, почти для любой теории. Собственно говоря, принимать во внимание подтверждающее свидетельство следует лишь в тех случаях, когда оно является результатом реальной "проверки теории на прочность" - попытки ее опровергнуть, которая оказалась безуспешной. Теория же, которая не опровергаема никаким мыслимым событием, является ненаучной; принципиальная неопровергаемость представляет собой не достоинство теории (как часто думают), а ее порок. Итак, критерием научного статуса теории является ее проверяемость и принципиальная опровергаемость (фальсифицируемость) [05]. Иными словами, наука (в отличие от псевдонауки) должна делать проверяемые предсказания ("Будет так-то и так-то, в противном случае я съем свою шляпу"), причем предсказания эти должны быть рискованными, не очевидными априори (не типа - "Солнце завтра по-прежнему взойдет на востоке").
Из рассмотренных выше теорий критерию фальсифицируемости отвечает лишь теория относительности: даже если в период ее выдвижения существующие измерительные инструменты не позволяли осуществить проверку, принципиальная возможность опровержения этой теории существовала уже тогда. Случай с астрологией - обратный; астрологи попросту игнорируют неблагоприятные для них свидетельства, а в своих прогнозах прибегают к обычному трюку всех прорицателей: предсказывают события столь неопределенно, чтобы предсказания всегда сбывались, то есть чтобы они были неопровергаемыми. (Помните, в "Ходже Насреддине": "Буду ли я счастлива в своем новом браке?" - трепетно спрашивала какая-нибудь почтенных лет вдова и замирала в ожидании ответа. "Да, будешь счастлива, если на рассвете не влетит в твое окно черный орел, - гласил ответ гадальщика. - Остерегайся также посуды, оскверненной мышами, никогда не пей и не ешь из нее." И вдова удалялась, полная смутного страха перед черным орлом, тягостно поразившим ее воображение, и вовсе не думая о каких-то презренных мышах; между тем, в них-то именно и крылась угроза ее семейному благополучию, что с готовностью растолковал бы ей гадальщик, если бы она пришла к нему с жалобами на неправильность его предсказания.")
Сложнее ситуация с марксистской социологией. В ранних своих формулировках она действительно давала проверяемые предсказания (например, Марксов анализ движущих сил и сроков грядущей "социальной революции"), которые все оказались опровергнутыми (революции происходили не в промышленно развитых, а в самых отсталых странах, и т.п.). Однако последователи Маркса, вместо того, чтобы признать это опровержение, переинтерпретировали и теорию, и свидетельства так, чтобы привести их в соответствие. Таким путем они "спасли" свою теорию, но при этом сделали ее неопровергаемой - и тем самым лишили ее научного статуса (в Советском Союзе марксизм превратился уже в чистое богословие - т.е. в комментирование священных текстов). Что же касается двух упомянутых психоаналитических теорий, то они являются изначально непроверяемыми и неопровергаемыми. Как подчеркивает Поппер, "это не означает, что Фрейд и Адлер вообще не сказали ничего правильного [...] Но это означает, что те "клинические наблюдения", которые, как наивно полагают психоаналитики, подтверждают их теорию, делают это не в большей степени, чем ежедневные подтверждения, обнаруживаемые в своей практике астрологами". Итак, по Попперу: теория относительности - научная и правильная, т.е. не опровергнутая, несмотря на все усилия; марксизм (ранний) - научная, но неправильная; психоанализ - правильная (в смысле - дающая позитивные практические результаты), но ненаучная.
Разумеется, Поппер нарисовал умышленно упрощенную картину. Ведь согласно его методологическим правилам, если теории противоречит некий факт, то она становится фальсифицированной и должна быть немедленно отброшена. Однако в реальности научное сообщество сплошь и рядом вынуждено сохранять заведомо "фальсифицированные" теории до тех пор, пока не появятся новые, более совершенные - "За неимением гербовой..."; с этим был вынужден согласиться и сам Поппер. Попперовский фальсификационализм пережил пик своей популярности в шестидесятые-семидесятые годы, а ныне уступил место более утонченным методологическим концепциям. Тем не менее, главные попперовские положения (что цена непроверяемой гипотезе, сколь бы красива она ни была - пятак в базарный день, и что суть научного исследования не в том, чтобы подбирать примеры, подтверждающие теорию, а чтоб искать всё новые способы для ее критической проверки) остаются в силе. Тем из вас, кто собирается в дальнейшем заниматься наукой, следует иметь это в виду.
Образование нашей планеты: "холодная" и "горячая" гипотезы. Гравитационная дифференциация недр. Происхождение атмосферы и гидросферы.
Рассказ о происхождении Земли и Солнечной системы нам придется начать издалека. В 1687 г. И. Ньютон вывел закон всемирного тяготения - "каждое тело во Вселенной притягивает остальные с силой, прямо пропорциональной произведению их масс, и обратно пропорциональной квадрату расстояния между ними". Теоретически закон всемирного тяготения позволяет рассчитать движения любого тела во вселенной под влиянием тяготения других тел. Но - увы! - только теоретически: уравнения, необходимые для описания движения всего трех изолированных тел под влиянием тяготения друг друга столь сложны, что их решение не удавалось получить почти три столетия, до 60-х годов нашего века. Понятно, что о полном решении для такой системы тел, как Солнечная система, и говорить не приходится. Что же до приближенных расчетов (которыми занимались многие выдающиеся математики и астрономы - Ж. Лагранж, П. Лаплас и другие), то они показывают, что возмущения в орбитах планет носят периодический характер: параметры орбиты меняются в одном направлении, затем в противоположном, и так до бесконечности. Итак, в самой по себе определяемой тяготением структуре Солнечной системы вроде бы нет ничего, что мешало бы ей существовать вечно; недаром сам Ньютон вопрос о происхождении Солнечной системы вообще не ставил.
Давайте, однако, задумаемся: если бы причиной движения планет было одно лишь тяготение - что с ними произошло бы? Правильно - они "упали" бы на Солнце. Планеты, тем не менее, благополучно двигаются себе по своим орбитам перпендикулярно действующей на них силе тяжести и при этом еще вращаются вокруг собственной оси. Это движение не могло возникнуть - и не возникло! - под влиянием тяготения Солнца; откуда же оно взялось?
Дело в том, что всякое вращающееся тело обладает определенным качеством, которое называется "моментом количества движения" (МКД). Величина МКД зависит от трех параметров: массы тела, его круговой скорости и расстояния до центра вращения. К XVIII веку было установлено, что МКД не возникает из ничего и не исчезает бесследно, а может лишь передаваться от тела к телу. Это - закон сохранения момента количества движения, принадлежащий к ряду законов сохранения (таких, как законы сохранения вещества, энергии и пр.). А коли так, то любая теория возникновения Вселенной (или Солнечной системы) как минимум не должна ему противоречить.
Итак, все тела, составляющие Солнечную систему, обладают собственным МКД; создать МКД невозможно - откуда же он взялся? Возможен следующий выход из этого тупика. Дело в том, что МКД могут различаться в зависимости от направления вращения: по и против часовой стрелки - положительный и отрицательный МКД. Если телу (или системе тел) сообщить два МКД - равной величины, но разного знака - то оба момента взаимно уничтожатся, и возникнет система, лишенная МКД. Но в таком случае верно и обратное: система, изначально не обладавшая МКД, может разделиться на две: одну с положительным, другую - с равным ему отрицательным МКД. Таким образом, МКД как бы появляется и исчезает без нарушения закона сохранения. Исходя из этого, можно предположить, что Вселенная вначале не обладала МКД, но затем одни ее части получили положительный момент, а другие - одновременно - отрицательный.
Так вот, если посмотреть на Солнечную систему "с высоты" - из некой точки над Северным полюсом Земли (и, соответственно, над плоскостью ее орбиты), то окажется, что Земля, Солнце и большинство иных тел вращаются вокруг своей оси против часовой стрелки; планеты вокруг Солнца и спутники вокруг планет - тоже. То есть - положительные и отрицательные МКД всех тел, составляющих Солнечную систему, отнюдь не уравновешиваются между собой; суммарный МКД этой системы очень велик, и необходимо выяснить его происхождение.
В 1796 году П.Лаплас сформулировал небулярную теорию, согласно которой последовательность событий при образовании Солнечной системы такова. Имеется первичное газо-пылевое облако (туманность - по латыни "небула"), возникшее в результате концентрации рассеянного межзвездного вещества под действием взаимного притяжения его частиц (просто в соответствии с законом всемирного тяготения). Небула не является идеальным шаром , и ее края - просто по теории вероятности - находятся на неодинаковом расстоянии от ближайшей небулы (или звезды), а потому притягиваются той с неодинаковой силой (которая, как мы помним, обратно пропорциональна квадрату расстояния). Этой неравновесности достаточно для того, чтобы наша небула получила первичный толчок, который и придаст ей вращательное движение, пусть и чрезвычайно слабое.
Как только небула начинает поворачиваться вокруг своей оси, в ней возникает сила тяжести (как в космическом корабле, который специально "раскручивают" для противодействия невесомости). Под воздействием силы тяжести небула должна начать сжиматься - т.е. ее радиус уменьшается. А мы с вами помним, что МКД (который есть величина постоянная) зависит от трех параметров: массы тела, радиуса и скорости его вращения; масса - тоже величина неизменная, поэтому уменьшение радиуса может быть компенсировано только увеличением скорости вращения. В результате огромный газовый шар будет вращаться все быстрее и быстрее, работая как центрифуга: под действием центробежной силы его экватор вспухает, придавая шару форму все более сплющенного эллипсоида. Затем наступает момент, когда все возрастающая центробежная сила на экваторе уравновешивает силу притяжения, и от него (экватора) начинает отслаиваться кольцо, а затем, по мере дальнейшего сжатия небулы, еще и еще. Вещество этих вращающихся колец начинает под действием взаимного притяжения его частиц конденсироваться в планеты, от которых, в свою очередь, отрываются их спутники.
Теория Лапласа, согласно которой Земля была изначально холодной, сохраняла популярность на протяжении почти столетия, хотя ей и противоречили некоторые астрономические данные (например - вращение Венеры и Урана в сторону, обратную всем остальным планетам и Солнцу). Однако ближе к концу XIX века, когда было твердо установлено, что температура в недрах нашей планеты чрезвычайно высока (по современным данным - свыше тысячи градусов), большинство ученых стало разделять мнение об изначально горячей Земле - огненном шаре, постепенно остывающем с поверхности. Поиски источника этого раскаленного вещества вполне естественно было начать с Солнца. В начале нашего века астрономы Т. Чемберлен и Ф. Мультон выдвинули, а Дж. Джинс математически обосновал планетезимальную теорию происхождения планет Солнечной системы. Суть ее состоит в том, что некогда поблизости от Солнца ("поблизости" - это по космическом масштабам) прошла другая звезда. При этом взаимное притяжение вырвало из каждой из них по гигантскому протуберанцу звездного вещества, которые, соединившись, составили "межзвездный мост", распавшийся затем на отдельные "капли" - планетезимали. Остывающие планетезимали и дали начало планетам и их спутникам.
Вторая половина нашего века, однако, стала временем возвращения к концепции изначально холодной Земли. Во-первых, нашлись серьезные, чисто астрономические, возражения против планетезимальной теории. Г. Рессел, например, обратил внимание на то простое обстоятельство, что если между Солнцем и проходящей звездой протянется лента из звездного вещества, то ее средняя часть (где притяжение двух светил взаимно уравновешивается) должна будет пребывать в полной неподвижности. И напротив, выяснилось, что некоторые оказавшиеся ошибочными положения Лапласа вполне могут быть откорректированы в рамках дальнейшего развития небулярной теории. (В качестве примера могут быть приведены гипотеза О.Ю. Шмидта - в ней газо-пылевое облако захватывается уже существующим на тот момент Солнцем, или более популярная ныне модель К. фон Вайцзекера; в последней вращающаяся небула представляет собой уже не гомогенный шар, как у Лапласа, а систему разноскоростных вихрей, несколько напоминающую шарикоподшипник. Ныне полагают также, что газ и пыль во вращающейся газо-пылевой туманности ведут себя по разному: пыль собирается в плоский экваториальный диск, а газ образует почти шарообразное облако, густеющее по направлению к центру туманности. Впоследствии пыль экваториального диска слипается в планеты, а газ под собственной тяжестью разогревается так, что "вспыхивает" в виде Солнца).
Более существенным для победы "холодной" концепции, однако, оказалось другое: был найден убедительный и при этом достаточно простой ответ на вопрос - откуда же берется тепло, разогревшее недра изначально холодной Земли до столь высоких температур? Этих источников тепла, как сейчас полагают, два: энергия распада радиоактивных элементов и гравитационная дифференциация недр. С радиоактивностью все достаточно ясно, да и источник это второстепенный - на него приходится, согласно современным оценкам, не более 15% энергии разогрева. Идея же гравитационной дифференциация недр (ее детальную разработку связывают с именем О.Г. Сорохтина) заключается в следующем.
Зная массу и объем Земли (они были рассчитаны еще в XVIII веке), легко определить усредненную плотность земного вещества - 5,5 г/см3. Между тем, плотность доступных нам для прямого изучения горных пород вдвое меньше: средняя плотность вещества земной коры составляет 2,8 г/см3. Отсюда ясно, что вещество в глубоких недрах Земли должно иметь плотность много выше средней.
Известно, что почти девать десятых массы Земли приходится на долю всего четырех химических элементов - кислорода (входящего в состав окислов), кремния, алюминия и железа. Поэтому можно с достаточной уверенностью утверждать, что более "легкие" наружные слои планеты состоят преимущественно из соединений кремния (алюмосиликатов), а "тяжелые" внутренние - железа.
В момент образования Земли ("горячим" или "холодным" способом - для нас сейчас неважно) "тяжелые" и "легкие" элементы и их соединения не могли не быть полностью перемешаны. Дальше, однако, начинается их гравитационная дифференциация: под действием силы тяжести "тяжелые" соединения (железо) "тонут" - опускаются к центру планеты, а "легкие" (кремний) - "всплывают" к ее поверхности. Давайте теперь рассмотрим этот процесс в мысленно вырезанном вертикальном столбе земного вещества, основание которого - центр планеты, а вершина - ее поверхность. "Тонущее" железо постоянно смещает центр тяжести этого столба к его основанию. При этом потенциальная энергия столба (пропорциональная произведению массы тела на высоту его подъема, что в нашем случае составляет расстояние между центром Земли и центром тяжести столба) постоянно уменьшается. Суммарная же энергия Земли, в соответствии с законами сохранения, неизменна; следовательно, теряющаяся в процессе гравитационной дифференциации потенциальная энергия может преобразовываться лишь в кинетическую энергию молекул - то есть выделяться в виде тепла.
Расчеты геофизиков показывают, что эта энергия составляет чудовищную величину 4*1030 кал (что эквивалентно триллиону суммарных ядерных боезапасов всех стран мира). Этого вполне достаточно для того, чтобы - даже не прибегая к помощи энергии радиоактивного распада - разогреть недра изначально холодной Земли до расплавленного состояния. При этом, однако, рассчитывая тепловой баланс Земли за всю ее историю, геофизики пришли к выводу, что температура ее недр лишь местами могла доходить до 1600оC, в основном составляя около 1200оC; а это означает, что наша планета, вопреки бытовавшим ранее представлениям, никогда не была полностью расплавленной. Разумеется, планета постоянно теряет тепловую энергию, остывая с поверхности, однако этот расход в значительной степени (если не полностью) компенсируется излучением Солнца.
Итак, Земля на протяжении всей своей истории представляет собой твердое тело (более того: в глубинах, при высоких давлениях, очень твердое тело), которое, однако, парадоксальным образом ведет себя при очень больших постоянных нагрузках как чрезвычайно вязкая жидкость. Сама форма планеты - эллипсоид с чуть выпяченным Северным полюсом и чуть вдавленным Южным - идеально соответствует той, что должна принимать жидкость в состоянии равновесия. В толще этой "жидкости" постоянно происходят чрезвычайно медленные, но немыслимо мощные движения колоссальных масс вещества, с которыми связаны вулканизм, горообразование, горизонтальные перемещения континентов и т.д. - их закономерности мы будем обсуждать в следующей главе. Здесь важно запомнить, что источником энергии для всех этих процессов является в конечном счете все та же самая гравитационная дифференциация вещества в недрах планеты. Соответственно, когда этот процесс завершится полностью, наша планета станет геологически неактивной, "мертвой" - подобно Луне. Согласно расчетам геофизиков, к настоящему моменту уже 85% имеющегося на Земле железа опустилось в ее ядро, а на "оседание" оставшихся 15% потребуется еще около 1,5 млрд. лет.
 
|
В результате гравитационной дифференциации недра планеты оказывается разделенными (как молоко в сепараторе) на три основных слоя - "тяжелый", "промежуточный" и "легкий". Внутренний, "тяжелый" слой (с плотностью вещества около 8 г/см3) - центральное ядро, состоящее из соединений железа и иных металлов; из 6400 км, составляющих радиус планеты, на ядро приходится 2900 км. Поверхностный, "легкий" слой (плотность его вещества около 2,5 г/см3) называется корой. Средняя толщина коры всего-навсего 33 км; она отделена от нижележащих слоев поверхностью Мохоровичича, при переходе через которую скачкообразно увеличивается скорость распространения упругих волн. Между корой и ядром располагается "промежуточный" слой - мантия; ее породы имеют плотность около 3,5 г/см3 и находятся в частично расплавленном состоянии. Верхняя мантия отделена от нижней мантии лежащим в 60-250 км от поверхности расплавленным слоем базальтов - астеносферой; верхняя мантия вместе с корой образует твердую оболочку планеты - литосферу (рисунок 4). Именно в астеносфере находятся магматические очаги, питающие вулканы, деятельности которых Земля обязана своей подвижной оболочкой - гидросферой и атмосферой.
Согласно современным представлениям, атмосфера и гидросфера возникли в результате дегазации магмы, выплавляющейся при вулканических процессах из верхней мантии и создающей земную кору. Атмосфера и гидросфера состоят из легких летучих веществ (соединений водорода, углерода и азота), содержание которых на Земле в целом очень мало - примерно в миллион раз меньше, чем в космосе. Причина такого дефицита состоит в том, что эти летучие вещества были "вымыты" еще из протопланетного облака солнечным ветром (т.е. потоками солнечной плазмы) и давлением света. В момент образования Земли из протопланетного облака все элементы ее будущей атмосферы и гидросферы находились в связанном виде, в составе твердых веществ: вода - в гидроокислах, азот - в нитридах (и, возможно, в нитратах), кислород - в окислах металлов, углерод - в графите, карбидах и карбонатах.
Современные вулканические газы примерно на 75% состоят из воды и на 15% - из углекислоты, а остаток приходится на метан, аммиак, соединения серы (H2S и SO2) и "кислые дымы" (HCl, HF, HBr, HJ), а также инертные газы; свободный кислород полностью отсутствует. Изучение содержимого газовых пузырьков в древнейших (катархейских) кварцитах Алданского щита показало, что качественный состав этих газов полностью соответствует тому, что перечислено выше. Поскольку эта первичная атмосфера была еще очень тонкой, температура на поверхности Земли была равна температуре лучистого равновесия, получающейся при выравнивании потока солнечного тепла, поглощаемого поверхностью, с потоком тепла, излучаемым ею; для планеты с параметрами Земли температура лучистого равновесия равна примерно 15оC.
В итоге почти весь водяной пар из состава вулканических газов должен был конденсироваться, формируя гидросферу. В этот первичный океан переходили, растворяясь в воде, и другие составные части вулканических газов - большая часть углекислого газа, "кислые дымы", окиси серы и часть аммиака. В результате первичная атмосфера (содержащая - в равновесии с океаном - водяные пары, CO2, CO, CH4, NH3, H2S и инертные газы, и являющаяся восстановительной) оставалась тонкой, и температура на поверхности планеты не отклонялась сколь-нибудь заметно от точки лучистого равновесия, оставаясь в пределах существования жидкой воды. Это и предопределило одно из главных отличий Земли от других планет Солнечной системы - постоянное наличие на ней гидросферы.
Как же изменялся объем гидросферы на протяжении ее истории? В расплавленном базальте (в астеносфере) при температуте 1000оС и давлении 5-10 тыс. атмосфер растворено до 7-8% H2O: именно столько воды, как установлено вулканологами, дегазируется при излиянии лав. Большая часть этой воды (имеющей, таким образом, мантийное происхождение) пополняла собою гидросферу, но часть ее поглощалась обратно породами океанической коры (этот процесс называется серпентинизацией). Расчеты геофизиков показывают, что в катархее и архее воды в океанских впадинах было мало - она еще не прикрывала срединно-океанические хребты; в океаническую кору она поступала не из океанов, а снизу - непосредственно из мантии. В начале протерозоя уровень океанов достиг вершин срединно-океанических хребтов, но на протяжении всего раннего протерозоя практически весь объем поступавшей в океаны воды поглощался породами океанической коры. К началу среднего протерозоя процессы серпентизации закончились, и океаническая кора обрела современный состав. С этого времени объем океанов вновь начал нарастать, и процесс этот будет продолжаться, постепенно замедляясь, и дальше - пока на Земле не прекратятся вулканические процесы.
Если спросить человека: "Отчего море соленое?", он почти наверняка ответит: "Оттого же, отчего солоны бессточные озера (вроде озера Эльтон, снабжающего нас пищевой поваренной солью): впадающие в море реки несут некоторое количество солей, потом вода испаряется, а соль остается". Ответ этот неверен: соленость океана имеет совершенно иную природу, чем соленость внутриконтинентальных конечных водоемов стока. Дело в том, что вода первичного океана имела различные примеси. Одним источником этих примесей были водорастворимые атмосферные газы, другим - горные породы, из которых в результате эрозии (как на суше, так и на морском дне) вымываются различные вещества. "Кислые дымы", растворяясь в воде, давали галогеновые кислоты, которые тут же реагировали с силикатами - основным компонентом горных пород, и извлекали из них эквивалентное количество металлов (прежде всего - щелочных и щелочноземельных - Na, Mg, Ca, Sr, K, Li). При этом, во-первых, вода из кислой становилась практически нейтральной, а во-вторых, соли извлеченных из силикатов элементов переходили в раствор; таким образом, вода океана с самого начала была соленой. Концентрация катионов в морской воде совпадает с распространенностью этих металлов в породах земной коры, а вот содержание основных анионов (Cl-, Br-, SO4-, HCO3-) в морской воде намного выше того их количества, которое может быть извлечено из горных пород. Поэтому геохимики полагают, что все анионы морской воды возникли из продуктов дегазации мантии, а все катионы - из разрушенных горных пород.
Главный фактор, определяющий кислотность морской воды - содержание в ней углекислоты (CO2 - водорастворимый газ, и в океанах его сейчас растворено 140 трл. т - против 2,6 трл.т, содержащихся в атмосфере). В океанах существует динамическое равновесие между нерастворимым карбонатом кальция CaCO3 и растворимым бикарбонатом Ca(HCO3)2: при недостатке CO2 "лишний" бикарбонат превращается в карбонат и выпадает в осадок, а при избытке CO2 карбонат превращается в бикарбонат и переходит в раствор. Карбонатно-бикарбонатный буфер возник в океане на самом начальном этапе его существования, и с тех пор он поддерживает кислотность океанской воды на стабильном уровне.
Что касается атмосферы, то ее состав стал меняться в протерозое, когда фотосинтезирующие организмы начали вырабатывать (в качестве побочного продукта своей жизнедеятельности) свободный кислород; сейчас считается твердо установленным, что весь свободный кислород планеты имеет биогенное происхождение. Кислород, в отличие от углекислоты, плохо растворим в воде (соотношение между атмосферным и растворенным в воде CO2 составляет, как мы видели, 1:60, а для O2 оно составляет 130:1), и потому почти весь прирост кислорода идет в атмосферу. Там он окисляет CO и CH4 до CO2, H2S - до S и SO2, а NH3 - до N2; самородная сера, естественно, выпадает на поверхность, углекислота и сернистый ангидрид растворяются в океане, и в итоге в атмосфере остаются только химически инертный азот (78%) и кислород (21%). Атмосфера из восстановительной становится современной, окислительной; впрочем, подробнее историю кислорода на Земле мы обсудим позднее, там, где речь пойдет о ранней эволюции живых существ (глава 5).
Помимо кислорода и азота, в атмосфере содержится небольшое количество так называемых парниковых газов - углекислый газ, водяной пар и метан. Составляя ничтожную долю атмосферы (менее процента), они, тем не менее, оказывают важное влияние на глобальный климат. Все дело в особых свойствах этих газов: будучи сравнительно прозрачными для коротковолнового излучения, поступающего от Солнца, они в то же время непрозрачны для длинноволнового - излучаемого Землею в космос. По этой причине вариации в количестве атмосферного CO2 могут вызывать существенные изменения теплового баланса планеты: с ростом коцентрации этого газа атмосфера по своим свойствам все более приближается к стеклянной крыше парника, которая обеспечивает нагрев оранжерейного воздуха путем "улавливания" лучистой энергии - "парниковый эффект".
Эволюция земной коры. Дрейф континентов и спрединг океанического дна. Мантийная конвекция.
В 1852 году Э. де Бомон предложил для объяснения процесса горообразования (орогенеза) контракционную теорию (от латинского "контракцио" -… Контракционная теория имела множество уязвимых мест. Подсчеты показывали, что… Отправным пунктом в построениях Вегенера было удивительное сходство береговых линий континентов по разные стороны…Происхождение жизни: абиогенез и панспермия. Гиперцикл. Геохимический подход к проблеме.
Что же касается происхождения жизни на Земле, то обычно проблему эту, еще со времен Э. Геккеля (1866), сводят к чисто химической задаче: как… Дело в том, что многие органические соединения представляют собой смесь двух… Сейчас известно, что все белки на нашей планете построены только из левовращающих аминокислот, а нуклеиновые кислоты -…А (дополнительная). Термодинамические подходы к сущности жизни. Второе начало термодинамики, энтропия и диссипативные структуры.
Первой созданной человеком машиной для превращения тепла в механическую энергию был паровой двигатель. Это устройство производит работу путем… Ну, ладно: нельзя так нельзя. Но уж по крайней мере всю работу, что содержится… Идеальная отдача K = (Т2-Т1)/Т2, где Т1 и Т2 - температура холодного и горячего резервуаров (в градусах абсолютной…Ранний докембрий: древнейшие следы жизни на Земле. Маты и строматолиты. Прокариотный мир и возникновение эукариотности
Один из ведущих специалистов по древнейшим формам жизни У. Шопф (1992), приводя соответствующую цитату, отмечает, что на протяжении почти ста лет… Как известно, осадочные породы можно разделить по их происхождению на… Как мы помним из главы 1, древнейшие из известных минералов имеют возраст 4,2 млрд. лет (оценка возраста Земли в…Поздний докембрий: возникновение многоклеточности. Гипотеза кислородного контроля. Эдиакарский эксперимент.
Малоразмерные фитопланктонные организмы (прежде всего - прокариотные) сталкиваются с одной проблемой, на которую впервые обратил внимание… Неприятностей этих можно избежать, лишь заставив организм перемещаться… Многоклеточность привела, среди прочего, к резкому повышению способности организмов создавать в своем теле запас…А (дополнительная). Взаимоотношения хищника и жертвы в экологическом и эволюционном масштабах времени.
Хотя злые языки из числа антиэволюционистов и утверждают, будто филетические ряды мало чем отличаются от известной карикатуры XIX века, изображающей… В случае таксона память эта обеспечивается передаваемым от предка к потомку… Итак, создавать теорию эволюции экосистем должны, на паритетных началах, экологи и палеонтологи, причем первым, по…Кембрий: "скелетная революция" и пеллетный транспорт. Эволюция морской экосистемы: кембрий, палеозой и современность.
Самое же интересное заключается в том, что составляющие эту современную биоту таксоны (а их состав на уровне типов с той поры уже принципиально не… Пытаясь разобраться в причинах этого события, называемого обычно… Начать тут придется издалека. Одной из самых важных задач современной океанологии является детальное изучение морских…Ранний палеозой: "выход жизни на сушу". Появление почв и почвообразователей. Высшие растения и их средообразующая роль. Тетраподизация кистеперых рыб.
 
|
Вплоть до самого недавнего времени человек выносил из школьного учебника биологии и популярных книг по теории эволюции такую примерно картину события, именуемого обычно "Выходом жизни на сушу". В начале девонского периода (или в конце силурийского) на берегах морей (точнее - морских лагун) появились заросли первых наземных растений - псилофитов (рисунок 29, а), положение коих в системе растительного царства остается не вполне ясным. Растительность сделала возможным появление на суше беспозвоночных животных - многоножек, паукообразных и насекомых; беспозвоночные, в свою очередь, создали пищевую базу для наземных позвоночных - первых амфибий (ведущих свое происхождение от кистеперых рыб) - таких, как ихтиостега (рисунок 29, б). Наземная жизнь в те времена занимала лишь чрезвычайно узкую прибрежную полоску, за которой простирались необозримые пространства абсолютно безжизненных первичных пустынь.
Так вот, согласно современным представлениям, в означенной картине неверным (или, по меньшей мере, неточным) является практически всё - начиная с того, что достаточно развитая наземная жизнь достоверно существовала много раньше (уже в следующем за кембрием ордовикском периоде), и кончая тем, что упомянутые "первые амфибии" наверняка были существами чисто водными, не имеющими связи с сушей. Дело, однако, даже не в этих частностях (о них мы поговорим в свой черед). Важнее другое: скорее всего, принципиально неверна сама формулировка - "Выход живых организмов на сушу". Есть серьезные основания полагать, что сухопутные ландшафты современного облика в те времена вообще отсутствовали, и живые организмы не просто вышли на сушу, а в некотором смысле создали ее как таковую. Впрочем, давайте по порядку.
Итак, первый вопрос - когда; когда же все-таки на Земле появились первые несомненно наземные организмы и экосистемы? Однако тут сразу возникает встречный вопрос: а как определить, что некий вымерший организм, с которым мы столкнулись - именно наземный? Это вовсе не так просто, как кажется на первый взгляд, ибо принцип актуализма здесь будет работать с серьезными сбоями. Типичный пример: начиная с середины силурийского периода в палеонтологической летописи появляются скорпионы - животные по нынешним временам вроде бы сугубо сухопутные. Однако сейчас уже достаточно твердо установлено, что палеозойские скорпионы [29] дышали жабрами и вели водный (или, по крайней мере, амфибиотический) образ жизни; наземные же представители отряда, у которых жабры превращены в характерные для паукообразных легкие "книжечного" типа (book-lungs), появились лишь в начале мезозоя. Следовательно, находки в силурийских отложениях скорпионов сами по себе ничего (в интересующем нас плане) не доказывают.
Более продуктивно здесь, как кажется, отслеживать появление в летописи не наземных (по нынешним временам) групп животных и растений, а определенных анатомических признаков "сухопутности". Так, например, растительная кутикула с устьицами и остатки проводящих тканей - трахеид наверняка должны принадлежать наземным растениям: под водой, как легко догадаться, и устьица, и проводящие сосуды ни к чему... Однако существует и иной - воистину замечательный! - интегральный показатель существования в данное время наземной жизни. Подобно тому, как показателем существования на планете фотосинтезирующих организмов является свободный кислород, показателем существования наземных экосистем может служить почва: процесс почвообразования идет только на суше, а ископаемые почвы (палеопочвы) хорошо отличимы по структуре от любых типов донных отложений.
Надо заметить, что почва сохраняется в ископаемом состоянии не слишком часто; лишь в последние десятилетия на палеопочвы перестали смотреть как на некую экзотическую диковинку и начали их систематическое изучение. В итоге в изучении древних кор выветривания (а почва есть не что иное, как биогенная кора выветривания) произошла подлинная революция, буквально перевернувшая прежние представления о жизни на суше. Самые древние палеопочвы найдены в глубоком докембрии - в раннем протерозое; в одной из них, имеющей возраст 2,4 млрд лет, С. Кемпбелл (1985) обнаружил несомненные следы жизнедеятельности фотосинтезирующих организмов - углерод со смещенным изотопным отношением 12С/13С. В этой связи можно упомянуть и обнаруженные недавно остатки цианобактериальных построек в протерозойских карстовых полостях: процессы карстования - образование котловин и пещер в водорастворимых осадочных породах (известняки, гипсы) - могут идти только на суше.
Другим фундаментальным открытием в этой области следует считать обнаружение Г. Реталляком (1985) в ордовикских палеопочвах вертикальных норок, прорытых какими-то достаточно крупными животными - видимо, членистоногими или олигохетами (дождевыми червями) [30]; в этих почвах нет никаких корней (которые обычно сохраняются очень хорошо), но есть своеобразные трубчатые тельца - Реталляк интерпретирует их как остатки несосудистых растений и/или наземных зеленых водорослей. В несколько более поздних, силурийских, палеопочвах найдены копролиты (окаменевшие экскременты) каких-то почвообитающих животных; пищей им, судя по всему, служили гифы грибов, составляющие заметную долю вещества копролитов (впрочем, не исключено, что грибы могли и вторично развиться на органике, содержащейся в копролитах).
Итак, к настоящему моменту два факта могут считаться установленными достаточно твердо:
- Жизнь появилась на суше очень давно, в среднем докембрии. Она была представлена, судя по всему, различными вариантами водорослевых корок (в том числе - амфибиотическими матами) и, возможно, лишайниками [31]; все они могли осуществлять процессы архаичного почвообразования.
- Животные (беспозвоночные) существовали на суше по меньшей мере с ордовика, т.е. задолго до появления высшей растительности (чьи достоверные следы по-прежнему остаются неизвестными до позднего силура). Средой обитания и пищей этим беспозвоночным могли служить упомянутые выше водорослевые корки; при этом сами животные неизбежно становились мощным почвообразующим фактором.
Последнее обстоятельство заставляет вспомнить одну старую дискуссию - о двух возможных путях заселения суши беспозвоночными. Дело в том, что неморские ископаемые этого возраста были очень редки, и все гипотезы на сей предмет казались лишь более или менее убедительными спекуляциями, не подлежащими реальной проверке. Одни исследователи предполагали, что животные вышли из моря напрямую - через литораль с водорослевыми выбросами и иными укрытиями; другие настаивали на том, что сперва были заселены пресноводные водоемы, и лишь с этого "плацдарма" началось впоследствии "наступление" на сушу. Среди сторонников первой точки зрения выделялись своей убедительностью построения М.С. Гилярова (1947), который, основываясь на сравнительном анализе адаптаций современных почвообитающих животных, доказывал, что именно почва должна была служить первичной средой обитания самых ранних жителей суши [32]. При этом надо учитывать, что почвенная фауна действительно крайне плохо попадает в палеонтологическую летопись и отсутствие ископаемых "документов" здесь вполне объяснимо. У этих построений, однако, был один по-настоящему уязвимый пункт: а откуда же взялась сама эта почва, если в те времена еще не было наземной растительности? Всем ведь известно, что почвообразование идет при участии высших растений - сам Гиляров называл настоящими почвами лишь те, что связаны с ризосферой, а все прочее - корами выветривания... Однако теперь - когда стало известно, что примитивное почвообразование возможно и с участием одних лишь низших растений - концепция Гилярова обрела "второе дыхание", а недавно была впрямую подтверждена данными Реталляка по ордовикским палеопочвам.
С другой стороны, несомненные пресноводные фауны (которые содержат, помимо всего прочего, дорожки следов на поверхности осадка) появляются намного позже - в девоне. В их состав входят скорпионы, мелкие (примерно в ладонь величиной) ракоскорпионы, рыбы и первые неморские моллюски; среди моллюсков есть и двустворчатые - длительно живущие организмы, неспособные переносить заморы и пересыхание водоемов. Фауны же с такими бесспорно-почвенными животными, как тригонотарбы ("панцирные пауки") и растительноядные двупарноногие многоножки существуют уже в силуре (лудловский век). А поскольку водная фауна всегда попадает в захоронения на порядок лучше, чем наземная, то все это позволяет сделать еще один вывод:
- Почвенная фауна появилась существенно раньше пресноводной. То есть - по крайней мере для животных пресные воды не могли играть роль "плацдарма" при завоевании суши.
Этот вывод, однако, заставляет нас вернуться к тому самому вопросу, с которого мы начинали свои рассуждения, а именно: вышли живые организмы на сушу или фактически создали ее как таковую? А.Г. Пономаренко (1993) полагает, что все сообщества, о которых шла речь выше, на самом деле трудно с определенностью назвать "наземными" или "сообществами внутриконтинентальных водоемов" (хотя по крайней мере маты должны были существенную часть времени находиться в воде). Он считает, что "существование настоящих континентальных водоемов, как текучих, так и стоячих, представляется весьма проблематичным до того, как в девоне сосудистая растительность несколько снизила скорость эрозии и стабилизировала береговую линию". Основные события должны были происходить в уже знакомых нам выположенных прибрежных амфибиотических ландшафтах без стабильной береговой линии - "не суша, не море" (см. главу 5 ).
Не менее необычная (с точки зрения сегодняшнего дня) обстановка должна была складываться и на водоразделах, занятых "первичными пустынями". В наши дни пустыни существуют в условиях недостатка влаги (при превышении испарения над выпадением осадков) - что и препятствует развитию растительности. А вот в отсутствие растений ландшафт парадоксальным образом становился тем более пустынным (по облику), чем больше выпадало осадков: вода активно размывала горные склоны, прорезая глубокие каньоны, при выходе на равнину давала конгломераты, а дальше по равнине распространялись разрозненные по поверхности псефиты [33], которые называют равнинным пролювием; ныне такие отложения слагают лишь конусы выноса временных водотоков.
Эта картина позволяет по-новому взглянуть на одно странное обстоятельство. Почти все известные силурийско-девонские наземные флоры и фауны найдены в различных точках древнего Континента красного песчаника (Old Red Sandstone), названного так по характерным для него породам - красноцветам; все местонахождения при этом связаны с отложениями, считающимися дельтовыми. Иными словами, выходит, что весь этот континент (объединявший Европу и восток Северной Америки) представляет собой как бы одну сплошную гигантскую дельту. Резонный вопрос: а где же располагались соответствующие реки - ведь для них на континенте таких размеров просто нет площадей водосбора! Остается предположить что все эти "дельтовые" отложения, по всей видимости, возникали именно в результате процессов эрозии в описанных выше "мокрых пустынях".
Итак, жизнь на суше (которая, впрочем, не вполне еще суша) вроде бы существует с незапамятных времен, а в конце силура просто-напросто появляется еще одна группа растений - сосудистые (Tracheophyta)... Однако на самом деле появление сосудистых растений - одно из ключевых событий в истории биосферы, ибо по своей средообразующей роли эта группа живых организмов не имеет себе равных, по крайней мере, среди эукариот. Именно сосудистая растительность и внесла, как мы увидим далее, решающий вклад в становление наземных ландшафтов современного облика.
Общепринятой считается та точка зрения, что некие водоросли, жившие вблизи берега, сначала "высунули голову на воздух", затем заселили приливно-отливную зону, а потом, постепенно превращаясь в высшие растения, целиком вышли на берег. За этим последовало постепенное завоевание ими суши. Предками высших растений большинство ботаников считает одну из групп зеленых водорослей - харовых (Charophyta); они образуют ныне сплошные заросли на дне континентальных водоемов - как пресных, так и соленых, тогда как в море (да и то лишь в опресненных заливах) найдены всего несколько видов. Харовые обладают дифференцированным талломом и сложно устроенными органами размножения; с высшими растениями их сближают несколько уникальных анатомических и цитологических признаков - симметричные спермии, присутствие фрагмопласта (структуры, участвующей в построении клеточной стенки в процессе деления) и наличие одинакового набора фотосинтетических пигментов и запасных питательных веществ.
Однако против этой точки зрения выдвигалось серьезное - чисто палеонтологическое - возражение. Если процесс преобразования водорослей в высшие растения действительно происходил в прибрежных водах (где условия для попадания в палеонтологическую летопись наиболее благоприятны), то почему мы не видим никаких его промежуточных стадий? Тем более, что и сами харовые появляются в позднем силуре - одновременно с сосудистыми растениями, а особенности биологии этой группы не дают оснований оснований предполагать для нее длительный период "скрытого существования"... Поэтому появилась парадоксальная, на первый взгляд, гипотеза: а почему, собственно говоря, появление макроостатков высших растений в конце силура должно однозначно трактоваться как следы их выхода на сушу? Может быть, совсем наоборот - это следы переселения высших растений в воду? Во всяком случае, многие палеоботаники (С.В. Мейен, Г. Стеббинс, Г. Хилл) активно поддерживали гипотезу о происхождении высших растений не от водных макрофитов (типа харовых), а от наземных зеленых водорослей. Именно этим сухопутным (а потому не имеющим реальных шансов попасть в захоронения) "первичным высшим растениям" могли принадлежать загадочные споры с трехлучевой щелью, которые весьма многочисленны в раннем силуре и даже в позднем ордовике (начиная с карадокского века).
Впрочем, недавно выяснилось, что, судя по всему, правы сторонники обеих точек зрения - каждые по-своему. Дело в том, что некоторые из микроскопических наземных зеленых водорослей имеют тот же самый комплекс тонких цитологических признаков, что харовые и сосудистые (см. выше); эти микроводоросли теперь стали включать в состав Charophyta. Таким образом, возникает вполне логичная и непротиворечивая картина. Первоначально существовала - на суше - группа зеленых водорослей ("микроскопические харовые"), от которой в силуре произошли две близкородственные группы: "настоящие" харовые, заселившие континентальные водоемы, и высшие растения, начавшие осваивать сушу, и лишь спустя некоторое время (в полном соответствии со схемой Мейена) появившиеся в прибрежных местообитаниях.
Из курса ботаники вам должно быть известно, что высшие растения (Embryophyta) делятся на сосудистые (Tracheophyta) и мохообразные (Bryophyta) - мхи и печеночники. Многие ботаники (например, Дж. Ричардсон, 1992) считают, что именно печеночники (если исходить из их современных жизненных стратегий) - главные претенденты на роль "первопроходцев суши": они живут ныне на наземных водорослевых пленках, в мелководных эфемерных водоемах, в почве - совместно с синезелеными водорослями. Интересно, что азотфиксирующая сине-зеленая водоросль Nostoc способна жить внутри тканей некоторых печеночников и антоцеротовых, обеспечивая своих хозяев азотом; это наверняка было очень важно для первых обитателей примитивных почв, где этот элемент не мог не быть в жестком дефиците [34]. Упоминавшиеся выше споры из позднеордовикских и раннесилурийских отложений наиболее схожи именно со спорами печеночников (достоверные макроостатки этих растений появляются позже, в раннем девоне).
 
|
Однако в любом случае мохообразные (если даже они действительно появились еще в ордовике) облик континентальных ландшафтов вряд ли изменили. Первые же сосудистые растения - риниофиты [35] - появились в позднем силуре (лудловский век); вплоть до раннего девона (жединский век) они были представлены крайне однообразными остатками единственного рода Cooksonia, простейшего и архаичнейшего из сосудистых. А вот в отложениях следующего века девона (зигенского) мы находим уже множество разнообразных риниофитов (рисунок 30). С этого времени среди них обособляются две эволюционные линии. Одна из них пойдет от рода Zosterophylum к плауновидным (в их число входят и древовидные лепидодендроны - одни из основных углеобразователей в следующем, карбоновом, периоде). Вторая линия (в ее основание обычно помещают род Psilophyton) ведет к хвощеобразным, папоротникам и семенным - голосеменным и покрытосеменным (рисунок 30). Даже девонские риниофиты еще очень примитивны и, честно говоря, неясно - можно ли назвать их "высшими растениями" в строгом смысле: у них есть проводящий пучок (правда, сложенный не трахеидами, а особыми вытянутыми клетками со своеобразным рельефом стенок), но отсутствуют устьица. Такое сочетание признаков должно свидетельствовать о том, что растения эти никогда не сталкивались с дефицитом воды (можно сказать, что вся их поверхность являет собою одно большое открытое устьице), и, по всей видимости, являлись гелофитами (то есть росли "по колено в воде", вроде нынешнего камыша).
Появление сосудистых растений с их жесткими вертикальными осями вызвало целый каскад экосистемных новаций, изменивших облик всей биосферы:
- Фотосинтезирующие структуры стали располагаться в трехмерном пространстве, а не на плоскости (как это было до сих пор - в период господства водорослевых корок и лишайников). Это резко увеличило интенсивность образования органического вещества и, тем самым, суммарную продуктивность биосферы.
- Вертикальное расположение стволов сделало растения более устойчивыми к занесению смываемым мелкоземом (по сравнению, например, с водорослевыми корками). Это уменьшило безвозвратные потери экосистемой неокисленного углерода (в виде органики) - совершенствование углеродного цикла.
- Вертикальные стволы наземных растений должны быть достаточно жесткими (по сравнению с водными макрофитами). Для обеспечения этой жесткости возникла новая ткань - древесина, которая после гибели растения разлагается относительно медленно. Таким образом, углеродный цикл экосистемы обретает дополнительное резервное депо и, соответственно, стабилизируется.
- Появление постоянно существующего запаса трудноразложимой органики (сконцентрированного в основном в почве) ведет к радикальной перестройке пищевых цепей. С этого времени большая часть вещества и энергии оборачивается через детритные, а не через пастбищные цепи (как это было в водных экосистемах).
- Для разложения трудноусваиваемых веществ, из которых состоит древесина - целлюлозы и лигнина - потребовались новые типы разрушителей мертвой органики. С этого времени на суше роль основных деструкторов переходит от бактерий к грибам.
- Для поддержания ствола в вертикальном положении (в условиях действия силы тяжести и ветров) возникла развитая корневая система: ризоидами - как у водорослей и мохообразных - тут уже не обойдешься. Это привело к заметному снижению эрозии и появлению закрепленных (ризосферных) почв.
С.В. Мейен полагает, что суша должна была покрыться растительностью к концу девона (зигенский век) [36], поскольку с начала следующего, карбонового, периода на Земле образуются уже практически все типы осадков, отлагающихся ныне на континентах. В дозигенские же времена континентальные осадки практически отсутствуют - видимо, в связи с их постоянным вторичным размывом в результате нерегулируемого стока. В самом начале карбона на континентах начинается угленакопление - а это свидетельствует о том, что на пути стока вод стояли мощные растительные фильтры. Не будь их, остатки растений непрерывно смешивались бы с песком и глиной, так что получались бы обломочные породы, обогащенные растительными остатками - углистые сланцы и углистые песчаники, а не настоящие угли.
Итак, возникшая в прибрежных амфибиотических ландшафтах густая "щетка" из гелофитов (можно назвать ее "риниофитный камыш") начинает действовать как фильтр, регулирующий плащевой сток: она интенсивно отцеживает (и осаживает) сносимый с суши обломочный материал и формирует тем самым стабильную береговую линию. Некоторым аналогом этого процесса может служить формирование крокодилами "аллигаторовых прудов": животные постоянно углубляют и расширяют населяемые ими болотные водоемы, выбрасывая грунт на берег. В результате их многолетней "ирригационной деятельности" болото превращается в систему чистых глубоких прудов, разделенных широкими облесенными "дамбами". Так и сосудистая растительность в девоне разделила пресловутые амфибиотические ландшафты на "настоящую сушу" и "настоящие пресноводные водоемы". Не будет ошибкой сказать, что именно сосудистая растительность стала истинным исполнителем заклинания: "Да будет твердь!" - отделивши оную твердь от хляби...
|
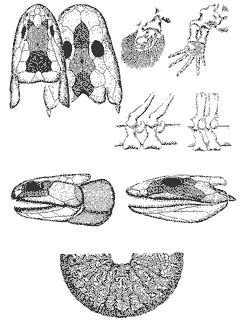
Именно с вновь возникшими пресноводными водоемами связано и появление в позднем девоне (фаменский век) первых тетрапод (четвероногих) - группы позвоночных, имеющих две пары конечностей; она объединяет в своем составе амфибий, рептилий, млекопитающих и птиц (попросту говоря, тетраподы - это все позвоночные, кроме рыб и рыбообразных). В настоящее время общепринято, что тетраподы ведут свое происхождение от кистеперых рыб (Rhipidistia) (рисунок 31); эта реликтовая группа имеет ныне единственного живого представителя, латимерию [37]. Достаточно популярная некогда гипотеза происхождения четвероногих от другой реликтовой группы рыб - двоякодышащих (Dipnoi), ныне практически не имеет сторонников.
Необходимо отметить, что в прежние годы возникновение ключевого признака тетрапод - двух пар пятипалых конечностей - считали их однозначной адаптацией к наземному (или по меньшей мере амфибиотическому) образу жизни. Ныне, однако, большинство исследователей склоняется к тому, что "проблема появления четвероногих" и "проблема их выхода на сушу" - суть вещи разные и даже не связанные между собою прямой причинной связью. Предки тетрапод жили в мелководных, часто пересыхающих, обильно заросших растительностью водоемах непостоянной конфигурации. Судя по всему, конечности появились для того, чтобы передвигаться по дну водоемов (это особенно важно, когда водоем обмелел настолько, что у тебя уже спина начинает вытарчивать наружу) и продираться сквозь плотные заросли гелофитов; особенно же полезны конечности оказались для того, чтобы при пересыхании водоема переползти посуху в другой, соседний.
Первые, девонские, тетраподы - примитивные амфибии лабиринтодонты [38] (название это происходит от их зубов с лабиринтоподобными складками эмали - структура, прямо унаследованная от кистеперых: см. рисунок 31), такие как ихтиостега и акантостега, в захоронениях всегда встречаются вместе с рыбами, которыми, судя по всему, и питались. Они были покрыты чешуей, как рыбы, имели хвостовой плавник (похожий на тот, что мы видим у сома или налима), органы боковой линии и - в некоторых случаях - развитый жаберный аппарат; конечность их еще не пятипалая (число пальцев достигает 8), и по типу сочленения с осевым скелетом - типично плавательная, а не опорная. Все это не оставляет сомнения в том, что существа эти были чисто водными (рисунок 32); если они и появлялись на суше при неких "пожарных" обстоятельствах (пересыхание водоема), то компонентом наземных экосистем наверняка не являлись [39]. Лишь много позже, в карбоновом периоде, появились мелкие наземные амфибии - антракозавры, которые, судя по всему, питались членистоногими, но об этом речь впереди (см. главу 10). |


Особого внимания заслуживает тот факт, что в девоне появляется целый ряд неродственных параллельных групп стегоцефалоподобных кистеперых рыб - причем как до, так и после возникновения "настоящих" тетрапод (лабиринтодонтов). Одной из таких групп были пандерихтиды - кистеперые, лишенные спинного и анального плавников, чего не бывает ни у каких других рыб. По строению черепа (уже не "рыбьему", а "крокодильему"), плечевого пояса, гистологии зубов и положению хоан (внутренних ноздрей) пандерихтиды очень сходны с ихтиостегой, однако приобрели эти признаки явно независимо. Таким образом, перед нами процесс, который можно назвать параллельной тетраподизацией кистеперых (он был детально изучен Э.И. Воробьевой). Как обычно, "заказ" на создание четвероногого позвоночного, способного жить (или, по крайней мере, выживать) на суше был дан биосферой не одному, а нескольким "конструкторским бюро"; "выиграла конкурс" в конечном итоге та группа кистеперых, которая "создала" известных нам тетрапод современного типа. Однако наряду с "настоящими" тетраподами еще долго существовал целый спектр экологически сходных полуводных животных (типа пандерихтид), сочетающих признаки рыб и амфибий - если так можно выразиться, "отходы" процесса тетраподизации кистеперых.
Поздний палеозой - ранний мезозой: криоэры и термоэры. Палеозойские леса и континентальные водоемы - растения и насекомые.
РИСУНОК 33. Расположение главных палеофлористических областей в позднем палеозое при современном положении материков (а) и на мобилистской… Причины климатических различий, существовавших в позднем палеозое между… Картина эволюции растительности от девона до наших дней выяснена (в общих чертах) достаточно давно. Для наиболее…Эволюция наземных позвоночных (1): поздний палеозой - ранний мезозой. Анамнии и амниоты. Две линии амниот - тероморфная и завроморфная.
Ситуация принципиально изменилась с начала карбона, когда в уже знакомых нам "лесах-водоемах" возникла богатая фауна из амфибиотических и… Чем объяснить итоговый эволюционный неуспех амфибий? Ведь этот класс процветал… Из курса зоологии вы должны помнить, что лягушка не имеет ни ребер, ни диафрагмы. Поэтому она - в отличие от нас -…Эволюция наземных позвоночных (2): поздний мезозой. Завроморфный мир. Маммализация териодонтов. Динозавры и их вымирание.
Эволюция архозавров выглядит так (рисунок 44). От пермо-триасовых текодонтов обособились к концу триаса все остальные отряды: крокодилы, птерозавры… От мелких хищных теропод позднее произошли птицы. Следует отметить, что… Когда говорят о структуре мезозойского сообщества наземных позвоночных, сразу замечают, что крупный размерный класс…Мезозойские биоценотические кризисы. "Ангиоспермизация мира" (средний мел) и "Великое вымирание" (конец мела). Импактные и биотические гипотезы.
Как уже было сказано, история наземных экосистем - это фактически история трех групп живых организмов: сосудистых растений, насекомых и позвоночных-тетрапод, а также их взаимоотношений. Основу окружающего нас кайнозойского мира составляют цветковые растения и экологически связанные с ними группы насекомых (таких, как, например, опылители - пчелы, мухи, бабочки), а также общественные насекомые (муравьи и термиты); наиболее заметными членами сообщества позвоночных являются плацентарные млекопитающие и птицы. Если говорить о водоемах, то тут доминируют настоящие костистые рыбы; верхние трофические этажи в морях формируют морские млекопитающие (китообразные и ластоногие) и акулы современного типа.
Мезозойский мир выглядел совершенно иначе. Растительную основу его составляли голосеменные (помимо хвойных - очень разнообразные гинкговые, саговники и родственые им беннетиты), в сообществе позвоночных доминировали архозавры - динозавры на земле, птерозавры в воздухе. Рыбы представлены в основном ганоидами [67], в морях главными хищниками являлись эвриапсидные рептилии (ихтиозавры и плезиозавры), а также головоногие (аммониты и белемниты). Различия среди насекомых не менее существенны: в те времена доминировали занимающие ныне подчиненное положение тараканы, цикады, сетчатокрылые и скорпионницы. Когда же именно возник этот наш кайнозойский мир? Произошла ли эта смена более или менее единовременно во всех группах, либо была растянутой во времени? Было ли это плавным замещением "ветеранов" "новобранцами", или, напротив, кризисом, сопряженным с массовыми вымираниями мезозойских групп?
О резких изменениях планетарной биоты в конце мелового периода ученые знали уже в XVIII веке; происходившие тогда события часто называют "Великим вымиранием". Наибольшее внимание обычно привлекает исчезновение гигантских рептилий (динозавров и птерозавров на суше, плезиозавров и мозозавров в море), однако помимо них в это время вымирают аммониты и белемниты, иноцерамы и рудисты (крупные - размером до метра - двустворки) и еще множество морских групп. Особено сильно пострадал планктон: раковинные простейшие - радиолярии и форамениферы, одноклеточные водоросли с известковым (кокколитофориды) и кремнеземовым (диатомеи) скелетом. При этом меловые виды обычны и разнообразны вплоть до последнего миллиметра отложений перед мезозойско-кайнозойской границей. Из данных магнитостратиграфии следует, что наибольшие изменения фауны соответствуют периоду обращенной полярности магнитного поля Земли (так называемый интервал 39), длившемуся не более 500 тыс лет. Иными словами, смена мезозойской биоты на кайнозойскую произошла внезапно и очень быстро, что наводит на мысль о какой-то глобальной катастрофе; а поскольку изменения эти охватили как морские, так и сухопутные группы, экологически не связанные между собою, логично связать ее с какой-то внешней по отношению к биосфере причиной - земной (катастрофический вулканизм и т.п.) или космической (вспышка сверхновой, многократное усиление излучения из космоса из-за переполюсовки магнитного поля планеты, падение гигантского астероида и т.п.).
Здесь следует сделать одно отступление, касающееся методологии. Мы с вами уже говорили о том, что основу европейского научного мышления составляет "Бритва Оккама" (см. главу 1-а) - из нескольких конкурирующих гипотез выбирается наиболее простая, не вводящая "избыточных сущностей". Всё так, но только вот представления о "простоте" и о том, какие из сущностей являются "избыточными" могут довольно сильно различаться у представителей разных наук.
Применительно к нашему конкретному случаю это означает следующее. Для физика более простыми, а потому более привлекательными, будут "однофакторные", импактные, гипотезы (извергнулись разом все вулканы - и всё живое отравилось серой и тяжелыми металлами; упал астероид - и поднятая им пыль настолько замутнила атмосферу, что растения перестали фотосинтезировать); представления же о системных свойствах биосферы являются "избыточными".
Иное дело - биолог; для него вполне очевидно, что экосистемы и биосфера являют собою не механические наборы живых организмов (вроде террариумов на Птичьем рынке), а высокоцелостные гомеостаты, обладающие своим собственным поведением, способные кондиционировать среду своего обитания и эффективно противостоять внешним возмущениям. Поэтому с точки зрения биолога более простыми будут гипотезы, предполагающие внутренние причины экосистемных смен - даже наиболее резких, кризисных (превращения червеобразной гусеницы в твердую неподвижную куколку, а куколки - в бабочку выглядят со стороны явными "кризисами" в процессе индивидуального развития насекомого), а внешние воздействия - "избыточной сущностью", которую можно привлекать лишь после того, как исчерпаны все иные, "внутрибиосферные" варианты объяснения. Это различие между "физиками" и "биологами" (названия эти, разумеется, условны) носит в некотором смысле мировоззренческий характер, и оттого лучше загодя обозначить свою позицию; я, как легко догадаться, занимаю "биологическую" позицию - положение обязывает...
Итак, возвращаемся к "Великому вымиранию". В середине 60-х годов при исследовании разреза пограничных мел-кайнозойских отложений в Губбио (Италия) в тонком слое глины, соответствующем фаунистической границе (его возраст - 67 млн лет), была обнаружена необычайно высокая концентрация редкого металла иридия (Ir), в 20 раз превышающая его среднее содержание в земной коре. Впоследствии такие иридиевые аномалии на мел-кайнозойской границе были найдены во множестве мест по всему миру, а содержание металла в некоторых из них превышает фоновое в 120 раз; протяженность периода, когда накапливались эти осадки, была (судя по толщине их слоя) очень невелика - не более 10 тысяч лет. В земной коре иридий редок потому, что он хорошо растворим в железе: почти весь земной иридий сконцентрирован сейчас в ядре планеты. Соответственно, его довольно много в веществе железных метеоритов, представляющих собой фрагменты планетных ядер.
На этом основании Л. Алварес (1980) предположил, что иридиевая аномалия - следствие удара о Землю крупного астероида, вещество которого рассеялось по всей ее поверхности (астероидная гипотеза). Чтобы дать наблюдаемое количество иридия, требуется тело внеземной природы диаметром приблизительно 10 км и массой 1010 т; при его падении на сушу возникла бы воронка диметром около 100 км (главным претендентом на эту роль считают кратер Чикксулуб на Юкатанском полуострове). Расчеты показывают, что при таком ударе в атмосферу было выброшено огромное количество пылевидного материала (в 60 раз больше массы самого астероида). Альварес и его сторонники полагают, что эта пыль обращалась вокруг Земли несколько лет, прежде чем выпала обратно на поверхность. Это плотное пылевое облако, сквозь которое слабо проходит солнечный свет, должно было сильно ослабить фотосинтез, что привело к гибели растений (прежде всего -планктонных водорослей, имеющих очень короткий жизненный цикл), а затем и питающихся ими животных. Кроме того, это должно было вызвать резкое охлаждение поверхности планеты ("астероидная зима"). В воде океанов из-за ослабления фотосинтеза накапливается CO2, что ведет к повышению ее кислотности, а это, в свою очередь, - к растворению раковин карбонатсекретирующих организмов (от фораменифер до иноцерамов).
Р. Кэрролл (1993) в своем учебнике палеонтологии пишет: "Ученые-физики, признавая основные выводы Альвареса, оспаривают некоторые детали. Специалисты по ископаемым остаткам обычно настроены очень критически"; А.Г. Пономаренко весьма точно отразил отношение палеозоологов к "импактным" гипотезам, завершив соответствующую лекцию словами: "К несчастью, воззрения эти в последние годы распространились, как чума". Палеонтологи, конечно, не могут профессионально оценить математические модели, из коих следует принципиальная, теоретическая возможность наступления "астероидной зимы" (хотя по некоторым расчетам пыль должна была осесть не через несколько лет, а уже через несколько недель, а гидрохимики напоминают о существовании в морской воде карбонатно-бикарбонатного буфера, изменить кислотность которого чрезвычайно трудно), но вот о конкретных событиях конца мела им очень даже есть чего сказать. Суть дела в том, что наличие причинной связи между иридиевой аномалией (которая факт) и позднемеловым вымиранием (которое тоже факт), вообще-то говоря, решительно ни из чего не следует.
Начнем с того, что иридиевых аномалий в отложениях самого различного возраста найдено (с той поры, как их стали целенаправленно искать) уже несколько десятков - но только они никак не связаны с крупными фаунистическими сменами. И наоборот - все попытки обнаружить следы астероидных импактов в горизонтах, соответствующих другим крупным вымираниям, таким, например, как пермо-триасовое (по справедливости "Великим" следовало бы назвать именно его - оно было куда более масштабным, чем мел-палеогеновое) ни к чему не привели. Существует целый ряд точно датированных метеоритных кратеров даже более крупного размера, чем предполагаемый Альваресом (до 300 км в диаметре), и при этом достоверно известно, что ничего серьезного с биотой Земли в те моменты не происходило. Скурпулезное, "по миллиметрам", изучение пограничных разрезов показало, что синхронность иридиевой аномалии и "Великого вымирания" сильно преувеличена. Массовое вымирание морских организмов было "мгновенным" лишь по геологическим меркам и продолжалось, по разным оценкам, от 10 до 100 тысячелетий (а вовсе не годы - как это должно было быть по импактным моделям). Последовательность исчезновения планктонных организмов в разных местах неодинакова, а пики вымирания могут расходиться со временем иридиевой аномалии на десятки тысяч лет, причем многие группы (например, белемниты) вымирают до аномалии, а не после нее.
В этом смысле весьма показательна ситуация с Эльтанинским астероидом (около 4 км в поперечнике), упавшим в позднем плиоцене, около 2.5 млн лет назад, на шельф между Южной Америкой и Антарктидой; остатки астероида были недавно подняты из образовавшегося в морском дне кратера. Последствия этого падения выглядят вполне катастрофическими - километровые цунами забрасывали морскую фауну вглубь суши; именно тогда на андийском побережье возникли очень странные захоронения фауны со смесью морских и сухопутных форм, а в антарктических озерах вдруг появляются чисто морские диатомовые водоросли. Что же касается отдаленных, эволюционно значимых последствий, то их просто не было: следы этого импакта заключены внутри одной стратиграфической зоны. То есть - абсолютно никаких вымираний за всеми этими ужасными пертурбациями не последовало.
Обращает на себя внимание тот факт, что из всех групп, ставших жертвами "Великого вымирания", наземными были лишь динозавры (которые к тому же, как мы помним, вымерли несколько позже иридиевой аномалии); не морской группой были и вымершие тогда же птерозавры, но те, судя по всему, были трофически связаны с морем. (Кстати, упоминавшееся выше грандиозное пермо-триасовое вымирание было чисто морским - на суше тогда тоже не произошло ничего примечательного.) Если же обратиться к меловой континентальной биоте, то окажется, что радикальные изменения в составе ее ключевых групп (растений и насекомых) произошли примерно на 30 млн лет раньше - в конце раннего мела; А.Г. Пономаренко удачно назвал эти события "ангиоспермизацией мира", выделив важнейшую их черту - массовое распространение ангиоспермов (цветковых растений). Здесь можно вспомнить, что ботаники иногда стремятся использовать свою собственную временную шкалу и делить историю растительности не на "палеозой, мезозой и кайнозой", а на "палеофит, мезофит и кайнофит". Так вот, граница между "мезофитом" и "кайнофитом" проходит внутри мелового периода: с точки зрения ботаника (и - как мы увидим дальше - энтомолога) поздний мел - это фактически уже кайнозой.
Появление и распространение покрытосеменных - одно из ключевых событий в истории биосферы, а вызванные им изменения сопоставимы по своим масштабам с появлением наземных (сосудистых) растений или животных с минеральным скелетом. Нынешний мир - ангиоспермный; голосеменным и споровым пришел в мелу такой же конец, как в позднем палеозое - водорослевым матам (при появлении сосудистой растительности). Ныне голосеменные растут лишь там, где им "позволяют": сосна растет на песках и болоте, а лиственница - на вечной мерзлоте совсем не оттого, что им там хорошо, а оттого, что на эти неудобья "не позарился" никто из цветковых. В некоторых ландшафтах (в тропических лесах и в пустынях) голосеменных нет вовсе [68].
Если задать ботанику вопрос - какие черты покрытосеменных привели их к эволюционному успеху, он наверняка начнет говорить о двойном оплодотворении, о триплоидном эндосперме и т.п.; это всё, конечно, верно - но немножко не о том. "С точки зрения биосферы" наиболее существенны, пожалуй, три свойства цветковых, позволившие им радикально изменить самую структуру мезозойских экосистем.
- Склонность к неотении (неотения - обретение организмом половозрелости на ранних (личиночных) стадиях своего индивидуального развития), то есть - в данном конкретном случае - к образованию травянистых форм, которые по сути дела есть древесные проростки, ставшие способными к размножению; напомним, что голосеменные "траву" не образуют вовсе - ни сейчас, ни в прошлом. Именно травянистые двудольные являются самыми агрессивными эксплерентами ("сорными растениями"), мгновенно захватывающими места нарушений растительного покрова, а травянистые однодольные, склонные к образованию многолетней дернины, наиболее эффективно препятствуют эрозии.
- Появление "мясистых" листьев с большим объемом фотосинтезирующей паренхимы, вызвавшее появление листогрызущих насекомых. Дело в том, что до того насекомые потребляли лишь генеративные органы, содержимое сосудов, даже древесину - но только не листья: в листе голосеменного (даже такого, как гинкго), попросту говоря, нечего есть. Ситуация менятся лишь с появлением цветковых - объеденные листья и мины (ходы, проедаемые личинками насекомых в толще листа) известны с самого начала позднего мела (сеноманский век). На покрытосеменных сформировалась общирная фауна листогрызущих насекомых, и уже после этого они перешли на голосеменные, папоротники и хвощи.
- Энтомофилия цветков, открывающая покрытосеменным путь к сложной коэволюции с насекомыми. Собственно говоря, сам цветок покрытосеменных, судя по всему, сформировался в процессе адаптаций к насекомоопылению для защиты завязи от повреждений ее опылителями.
 
|
Ангиоспермы впервые появляются в палеонтологической летописи в начале раннего мела, в неокоме; на протяжении барремского и аптского веков они представлены единичными находками, в альбе идет стремительное нарастание их числа, а уже в самом начале позднего мела, в сеномане, цветковые составляют большую часть ископаемых флор - то есть наступает "кайнофит" (рисунок 50). Среднемеловая экспансия цветковых была столь стремительной (она фактически уложилась в ничтожный по геологический меркам отрезок от 115 до 108 млн лет), что Дарвин в свое время в сердцах назвал этот эпизод палеонтологической истории "отвратительной тайной - abominable mistery". Делались попытки объяснить это тем, что цветковые на самом-то деле появились на Земле давным-давно, минимум в юре - просто они обитали лишь в горах (не попадая по этой причине в палеонтологическую летопись), а в середине мела заселили, наконец, приводные местообитания, благоприятствующие попаданию в захоронения. Вообще такой "период скрытого существования" сбрасывать со счетов никак нельзя - но в данном случае эта гипотеза не проходит: дело в том, что пыльца покрытосеменных (которая, ясное дело, летает везде, и ее не "упрячешь" в горы) появляется в летописи одновременно с их макроостатками. По всем признакам, цветковые действительно возникли в начале мела, причем именно в приводных местообитаниях.
В 70-х годах советские палеоэнтомологи исследовали множество местонахождений меловых насекомых в Сибири и Монголии, "закрасив" тем самым самое крупное "белое пятно" в геологической истории класса. Оказалось, что у насекомых график, который отражает долю вымерших таксонов в фаунах разных времен, ведет себя довольно неожиданным образом (рисунок 51). Доля вымерших семейств мало меняется на протяжении большей части мезозоя, и на границе юры и мела составляет около 60% ; затем, в течении раннего мела, доля эта стремительно падает от 60 до 20% , в позднем мелу это падение резко замедляется (от 20 до 5%), а к началу кайнозоя кривая вновь "выходит на плато", только уже в районе 5%. Иными словами, у насекомых самая резкая фаунистическая смена, связанная с массовым вымиранием мезозойских групп, приходится не на конец мела, а на его середину - в точности как у растений. На этом основании В.В. Жерихин (1978) предложил следующую модель среднемелового биоценотического кризиса.
 
|
Покрытосеменные возникли в начале мелового периода, первоначально как ценофобы - растения, не входящие в закономерные, сложившиеся в длительной коэволюции сукцессионные ряды [69]. Они росли в качестве "сорной" растительности по свободным от других растений участкам (на речных отмелях, береговых оползнях, гарях), которые соседствовали с гораздо более обширными участками, занятыми зрелыми сообществами мезофитной растительности. Покрытосеменные были исходно энтомофильными и смогли сформироваться лишь в окружении энтомофильных же голосеменных (таких, как беннетиты, саговники, кейтониевые), используя уже сложившийся к тому времени комплекс их опылителей. Но если покрытосеменные оказались среди пионерных растений мезофита единственными энтомофилами (а, судя по всему, так оно и было), то это должно было дать им гиганские преимущества: именно для локально распределенных, не образующих сплошных зарослей пионеров насекомоопыление особенно выгодно; впрочем, возможно, имело место и прямое конкурентное вытеснение покрытосеменными энтомофильных голосеменных из пионерных сообществ - за счет более эффективного привлечения насекомых, лучшей защиты от повреждения завязи опылителями и т.д.
Основой стремительной среднемеловой экспансии цветковых стало то, что им удалось закрепиться в качестве нормальной пионерной растительности - для чего решающими факторами стали их исходная энтомофильность и наличие среди них травянистых форм (что во много раз убыстряет зарастание поврежденных участков). Покрытосеменные не пытались потеснить всю мезофитную растительность, что было абсолютно нереально; они "всего-навсего" конкурентно вытеснили прежних пионеров - и тем самым блокировали все последующие стадии мезофитной экогенетической сукцессии. Разрушение существовавших в то время закономерных сукцессионных рядов (попросту говоря - мезофитная растительность продолжала существовать там, где она существует, но потеряла способность восстанавливаться после экзогенных нарушений), вызвало полный развал мезозойских наземных экосистем и массовое вымирание входивших в них животных; наиболее интенсивно эта деструкция шла в альбе (захватывая конец апта и начало сеномана).
В.В. Жерихин (1978) обратил внимание на любопытнейшую черту энтомофаун этого времени: многие семейства, характерные для мезозоя, обнаруживаются в них в последний раз, тогда как сколько-нибудь заметного появления новых групп не отмечается, и при этом появляется целый ряд представителей реликтовых для того времени (раннемезозойских и даже позднепалеозойских) групп. Это "всплывание реликтов" считают одним из наиболее ярких симптомов кризиса, охватившего среднемеловые сообщества: вымирание доминирующих раннемеловых групп (наиболее тесно связанных с существующей сукцессионной системой) создает экологический вакуум, который ненадолго заполняют древние формы, оттесненные к тому времени "на обочину жизни".
Складывающиеся на совершенно новой основе раннекайнофитные сообщества первоначально были несбалансированными и неустойчивыми. Слабость биоценотической регуляции и обилие незанятых экологических ниш вели к очень быстрой, взрывной, эволюции различных групп организмов (из насекомых здесь можно отметить высших бабочек и мух, паразитических перепончатокрылых, а также общественных насекомых - муравьев и термитов), по мере же насыщения биоценозов темпы эволюции снижались. Сложные трофические цепи в новых кайнофитных сообществах сформировались очень быстро (листья, поврежденные минирующими бабочками, сами эти бабочки и их высокоспециализированные паразиты из числа наездников Braconidae появляются в летописи почти одновременно, в сеномане) и в дальнейшем менялись мало. "По экологическому облику и систематическому составу , - заключает Жерихин (1980), - позднемеловые фауны значительно сильнее отличаются от раннемеловых, чем от кайнозойских, и могут рассматриваться как ранний, первоначально сильно обедненный, вариант этих последних. В сущности, по набору жизненных форм они имеют не так уж много отличий от современных".
Если вернуться к истокам среднемелового кризиса - самому возникновению покрытосеменных, - то здесь обнаруживается еще один интересный аспект, на который обратил внимание А.Г. Пономаренко (1997). Дело в том, что, по данным последних лет, серьезные эволюционные изменения среди насекомых начинаются несколько раньше широкого распространения покрытосеменных - с конца юры. Это, казалось бы, противоречит гипотезе о причинной связи между этими явлениями - но только на первый взгляд. Именно в это время начинается очередная "...зация" - ангиоспермизация голосеменных, и широко распространяются проангиоспермы (термин В.А. Красилова, 1989) - голосеменные с отдельными чертами цветковых (таковы, например, гнетовое Eoantha с крупными, явно энтомофильными "цветами", или беннетит Manlaia, имитирующий осоку - см. рисунок 52). "С точки зрения экосистемы" точное систематическое положение всех этих проангиоспермов не столь уж важно - все они могут в некоторой степени играть экологическую роль настоящих покрытосеменных.
 
|
Неудивительно, что параллельно с энтомофильными голосеменными появляются антофильные насекомые; имитаторам цветковых в растительном мире сопутствуют столь же многочисленные имитаторы бабочек среди насекомых (таковы, например, Calligrammatidae из сетчатокрылых, или Aneuretopsychidae из скорпионниц - см. рисунок 52). Важнее же всего то, что здесь складывается система с положительной обратной связью: наличие у некого беннетита комплекса опыляющих его насекомых стимулирует появление энтомофилии у его соседей-голосеменных, что, в свою очередь, расширяет экологическую нишу этих насекомых-опылителей, и т.д. Каждый шаг на этом пути увеличивает вероятность следующего и делает появление покрытосеменных (т.е. растений с полным синдромом ангиоспермных признаков) совершенно неизбежным; какая же конкретно группа голосеменных даст начало "настоящим цветковым" (равно как - какие именно из мекоптероидных насекомых породят "настоящих бабочек"), вообще-то говоря, не столь уж важно. Именно эти взаимно индуцируемые процессы Пономаренко и назвал "ангиоспермизацией мира".
"Ангиоспермизация мира" захватывает не только насекомых, но и все прочие компоненты континентальных экосистем. С начала мела на суше стремительно распространяются птицы (это особенно хорошо видно по укрупнению птерозавров - те полностью уходят в нишу крупных парителей) и териевые млекопитающие; в пресных водоемах настоящие костистые рыбы вытесняют ганоидов. После апт-альбского кризиса мир обретает вполне знакомый нам облик; правда, в нем наличествуют динозавры (и не просто "наличествуют" - они вполне процветает на протяжении большей части позднего мела), однако судьба этой группы была рассмотрена нами выше (глава 11), и нет оснований напрямую связывать ее вымирание с драматическими изменениями в морской биоте на границе мела и кайнозоя. Таким образом, мезозойско-кайнозойская смена на суше произошла много раньше, чем в море.
Ну, а каковы же всё-таки причины позднемелового "Великого вымирания"? Если не падение астероида и прочие "импакты" - то что же? Давайте прежде всего попытаемся найти какие-нибудь общие черты для этого события и для другого морского вымирания - пермско-триасового. Оно было самым крупным в истории Земли - тогда в морских группах вымерло 50% семейств, 70% родов и более 90% видов (для сравнения: при мел-кайнозойском вымирании исчезло менее четверти семейств). Помимо прямого исчезновения множества палеозойских групп (трилобиты, палеозойские кораллы - ругозы и табуляты) произошла радикальная смена структуры морских экосистем (роль основных рифостроителей перешла от мшанок к кораллам, донных фильтраторов - от брахиопод к двустворчатым моллюскам, и т.п.). Причины этой грандиозной фаунистической смены остаются загадкой; стандартные объяснения вроде "глобального катастрофического вулканизма" или морской регрессии, резко сократившей площадь шельфов (основной зоны обитания морских организмов) производят впечатление явно придуманных "за неимением лучшего". Мне лично наиболее остроумной и логичной (хотя и не бесспорной) представляется гипотеза Х. Таппан - специалиста по фитопланктону.
В 1986 году Таппан опубликовала работу под названием "Фитопланктон: за солью на планетарном столе (Phytoplancton: below the salt at the global table)"; название это требует пояснения. В средневековом английском замке представители всех социальных слоев ели за общим длинным столом. Дворяне, рыцари и почетные гости сидели во главе, и большая солонка, стоящая примерно посередине стола, как бы отделяла их места от мест слуг, солдат и "гостей второй свежести", занимающих другой его конец; места эти называли, соответственно, "перед солью" и "за солью" ("above and below the salt"). Кушанья, как легко догадаться, подавали сперва на тот конец, что "перед солью", а "за соль" доходило - уж что останется...
Так вот, по мысли Таппан, наземная биота планеты всегда занимает место "во главе стола", а океанская - оказывается "за солью". Необходимые для жизнедеятельности фитопланктона биогены (прежде всего азот и фосфор) поступают с суши, за счет эрозии; наземная растительность, между тем, стремится подавить эрозию и оставить как можно больше биогенов для собственного потребления (помните - замыкание геохимических циклов?). Таким образом, все заметные эволюционные успехи наземной растительности замедляют поступление биогенов в океан, что со временем вызывает вымирание соответствующих групп фитопланктона; фитопланктон же - основа первого трофического уровня любых морских экосистем, так что дальше разваливается вся цепь. Следует помнить, однако, что инерция такой системы огромна, и между "прогрессивным изменением" в наземной растительности и вызванным им "фитопланктонным шоком" в океане проходит ощутимый срок.
Подойдя с этих позиций к пермо-триасовому и мел-палеогеновому морским вымираниям, мы увидим, что в поздней перми и в позднем мелу действительно произошли два серьезных изменения в структуре наземной растительности, прямо влияющие на регуляцию потока биогенов с суши. В перми высшая растительность впервые начинает заселять водоразделы, тормозя их прежде беспрепятственный размыв; этому способствует возникновение засухоустойчивых голосеменных (типа вольциевых) с пикноксилической древесиной и мелкими, сильно склеротизованными листьями (а также, как мы помним, появление насекомых с водными личинками - см. главу 9). В позднем мелу появляется трава, образующая дернину - наиболее эффективный из известных подавителей эрозии.
Что же касается импактов, то они, по всей видимсти, не представляют угрозы для сбалансированных, "здоровых" экосистем, но вот если те уже находятся в состоянии кризиса (вроде планктона, столкнувшегося с серьезными глобальными изменениями в порядке поступления биогенов), то кратковременная "астероидная зима" вполне способна сыграть роль "последней соломинки, ломающей спину верблюда". Тут возможна аналогия с выпадением соли из пересыщенного раствора: никому ведь не придет в голову считать причиной этого процесса упавший в сосуд кристаллик, непосредственно инициировавший его начало.
Кайнозой: наступление криоэры. Новые типы сообществ - тропические леса и травяные биомы. Эволюция млекопитающих и появление человека.
Согласно моделям климатологов, наиболее интенсивный теплообмен будет достигнут при таком взаиморасположении материков и полюсов, когда планета имеет… В кайнозое картина становится близка к обратной, и причиною тому - перемещения… Надо заметить, что отсутствие в мезозое холодных климатических зон давно не вызывает сомнения у палеоклиматологов.…Четвертичный период (антропоген): Великое оледенение. Ледниковая теория. Перигляциальные сообщества и мамонтовая фауна.
Вероятно всем вам доводилось встречать где-нибудь на краях полей и лесных опушках окатанные гранитные валуны, иногда с характерной… Здесь необходимо сделать одно замечание. Читатель современных учебников и… Так, в 1821 году Бакленд исследовал найденные в одной из йоркширских пещер скелеты гигантских гиен, а также…А (дополнительная). Историческая биогеография. Викариантная модель и концепция "оттесненных реликтов". Фитоспрединг.
Возможных вариантов тут два: то ли верблюды этих континентов по каким-то причинам не достигли, то ли они здесь раньше жили, но вымерли. Но можно ли… Давайте попытаемся реконструировать историю сумчатых, исходя лишь из их… Рассмотренная ситуация наглядно демонстрирует два момента. Во-первых, насколько опасно в историко-биогеографических…Рекомендуемая литература
- Художественная
- А. Конан-Дойл "Затерянный мир"
- В.А.Обручев "Плутония"; "Земля Санникова"
- Ж. Рони Старший "Борьба за огонь"; "Пещерный лев"
- М. Крайтон "Парк юрского периода"
- Научно-популярная
- А. Азимов .....................
- С. Уэда "Новый взгляд на Землю"
- А.С. Монин "Популярная история Земли"
- Дж. Имбри, К.П. Имбри "Тайны ледниковых эпох"
- Юл. Медведев "Судеб посланник" (в книге "Бросая вызов")
- И.А. Ефремов "Дорога ветров"
- Дж. Смит "Старина четвероног"
- А.К. Рождественский "На поиски динозавров в Гоби"
- И. Аугуста, З. Буриан "По путям развития жизни"; "Летающие ящеры и древние птицы"; "Ящеры древних морей"; "Гиганты суши"; "Книга о мамонтах"; "Жизнь древнего человека"
- К.Л. Фентон, М.А. Фентон "Каменная книга. Летопись доисторической жизни"
- Ю.А. Орлов "О мире древних животных"
- Д. Эттенборо "Жизнь на Земле"
- И. Яковлева, В. Яковлев "По следам минувшего"
- М.Ф. Ивахненко, В.А. Корабельников "Живое прошлое Земли"
- А. Гангнус "Через горы времени"
- Г.А. Заварзин "Бактерии и состав атмосферы Земли"
- И.Н. Крылов "На заре жизни"
- А.Ю. Розанов "Что произошло 600 миллионов лет назад"
- С.В. Мейен "Из истории растительных династий"; "Следы трав индейских"
- Н. Эйдельман "Ищу предка"
- Р. Фоули "Еще один великолепный вид"
- В.А. Красилов "Нерешенные вопросы теории эволюции"
- Научная и учебная
- А.С. Монин, ...... Шишков "История климата"
- Р. Кэрролл "Палеонтология и эволюция позвоночных" (3 тома)
- В.В. Друщиц "Палеонтология беспозвоночных"
- С.В. Мейен "Основы палеоботаники"; "Историческое развитие класса насекомых", Труды ПИН АН СССР, т.175
- В.В. Жерихин "Развитие и смена меловых и кайнозойских фаунистических комплексов" (Общая часть)
- Ф. Дарлингтон "Зоогеография"
- Дж. Симпсон "Великолепная изоляция"
- С.М. Разумовский "Закономерности динамики фитоценозов"
- О.Г. Кусакин, А.Л. Дроздов "Филема органического мира"
- В.А. Красилов "Происхождение и ранняя эволюция цветковых растений"; "Эволюция и биостратиграфия"
- Л.П. Татаринов "Очерки по теории эволюции"
- Т. де Шарден "Феномен человека"
- Э. Шредингер "Что такое жизнь с точки зрения физика"
- И. Пригожин, И. Стенгерс "Порядок из хаоса"
Словарь терминов
Словарь названий живых и ископаемых организмов
Примечания
- Я буду употреблять здесь привычный для всех оборот - "отношения хищник-жертва", хотя в действительности речь пойдет об отношениях между любыми консументами и их добычей (в частности - фитопланктоном).
- Имеются в виду одноядерные одноклеточные; некоторые многоядерные простейшие (например, знакомые нам ксенофиофоры) и сифоновые водоросли могут достигать нескольких сантиметров.
- Во всех обсуждаемых случаях, говоря о размерах организмов, имеют в виду так называемый "размер, приведенный к шару" - РПШ. РПШ взрослого человека, например, составляет 60 см - именно таков будет диаметр шара, имеющего массу 80 кг при плотности человеческого тела.
- Здесь возможна и еще одна аналогия: военная техника ("хищник") всегда опережает в своем развитии гражданскую ("жертва"), но при этом созданные для военных целей технологии неизбежно приходят потом в мирные области (баллистические ракеты - космонавтика, СОИ - мобильная телефонная связь, и т.д.).
- В конце прошлого века из докембрийских отложений Франции, датируемых ныне 1 млрд. лет были описаны сферические остатки, отнесенные к радиоляриям. Хотя ошибочность этого отнесения показана уже давно, упоминания о "докембрийских радиоляриях" продолжают встречаться не только в научно-популярной литературе, но и в учебниках палеонтологии.
- Симбиотические микроорганизмы составляют до 80% объема тела современных губок.
- Закон Бернулли гласит, что чем выше скорость потока жидкости или газа, тем ниже создаваемое им поперечное давление.
- Здесь следует заметить, что первые трилобиты (Olenelidae) были не детритофагами, а хищниками; они появились в раннем кембрии, жили вместе с археоциатами и исчезли одновременно с ними.
- Описаны очень интересные нижнекемриские следовые дорожки: по поверхности осадка идет общий след многощетинкового червя и трилобита, а потом след червя обрывается - и дальше ползет один трилобит (успешно догнавший и съевший свою жертву).
- Вторично размер аммонитов на короткое время возрос перед самым их вымиранием, в конце мела (раковина Pachydiscus имеет диаметр более 2 м).
- В различных научно-популярных изданиях акул принято называть очень древней группой, чуть ли не "живыми ископаемыми". Это не совсем верно. Пластинчатожаберные хрящевые рыбы действительно известны начиная с карбона, однако те из них, кого называют "акулой" неспециалисты (крупный высокоскоростной хищник с телом гидродинамически совершенной формы - одним словом, персонаж фильма "Челюсти") вперыве появились в юре, а современные их семейства - лишь в мелу.
- Сочетание "хищная акула" не есть "масло масляное": самые крупные из современных акул - китовая и гигантская (15 и 12 м, соответственно) - питаются планктоном.
- Скорпионы представляют собою специализированную группу уже знакомых нам (по главе 7) морских ракоскорпионов-эвриптерид, представители которой перешли от плавания к хождению по дну и, приобретя мелкие размеры, освоили сначала морскую литораль, а потом и сушу.
- С находкой кембрийских морских многоножкоподобных членистоногих их существование и на раннепалеозойской суше кажется вполне вероятным, хотя достоверные находки многоножек в континентальных отложениях появляются лишь в позднем силуре.
- Не исключено, что уже в венде на суше существовали и макроскопические растения. В это время на талломах некоторых водорослей (Kanilovia) возникают загадочные сложноустроенные микроструктуры в виде зигзагообразно разрывающейся повдоль спиральной хитиноидной ленты. М.Б.Бурзин (1996) вполне логично предположил, что они служат для разбрасывания спор - а такой механизм необходим лишь в воздушной среде.
- Некоторые авторы (например, Л.С.Берг) считали почвенную жизнь вообще первичной.
- Псефиты - рыхлые осадки из обломочного материала, более грубые, чем "глина" (пелиты) и "песок" (псаммиты).
- Ни одно из высших растений не способно к азотфиксации, т.е. к преобразованию азота из атмосферного газа N2 в усваиваяемую форму (например, ионов NO3 - ); это - дополнительный аргумент в пользу того, что к моменту появления на суше высших растений там уже давно существовали прокариотические сообщества, которые обогатили почву азотом в доступной форме.
- Более привычное название - псилофиты - сейчас не употребляют по номенклатурным соображениям. В литературе последних лет вы можете столкнуться с еще одним названием - проптеридофиты.
- К концу девона уже появились представители почти всех основных отделов высших растений, не только споровых (плауновидные, папоротникообразные, хвощеобразные), но и голосеменных (гинкговые).
- Широко известна истинно романтическая история открытия этого "живого ископаемого", описанная в замечательной книге Дж.Смита "Старина четвероног". Следует, правда, заметить, что образ жизни латимерии не имеет ничего общего с тем, что вели девонские рипидистии: она живет в Индийском океане на глубинах в несколько сот метров.
- Старое название, которое вы можете встретить в книгах - стегоцефалы - сейчас не употребляют.
- Не называем же мы "наземным существом" угря, который способен ночью, по росистой траве, переползти из одного водоема в другой, преодолевая расстояние в несколько сот метров!
- Палеозойские растения, сочетающие в себе признаки голосеменных и папоротникообразных (семена и расчлененную "папоротниковую" листву) раньше выделяли в отдельный класс голосеменных - птеридоспермы, или семенные папоротники. Теперь, однако, их считают искусственной, сборной группой (не "кладой" - самостоятельной эволюционной ветвью, а "градой" - уровнем развития). То есть - самые различные таксоны голосеменных прошли в своем эволюционном развитии "птеридоспермовую стадию", а разные группы "семенных папоротников" теперь включены в соответствующие классы голосеменнных (Glossopteridales - в гинкговых, Lagenostomales - в цикадовых, и т.д.)
- Маноксилические (что означает "малодревесинные") стволы состоят преимущественно из коры - в противовес пикноксилическим (состоящим преимущественно из древесины). Такие стволы не выносят промораживания, и - если только не принадлежат однолетникам (что легко определить по их толщине) - считаются свидетельством безморозного климата.
- Сферический сегмент - часть шара, отделенная секущей плоскостью.
- Интересно, что подобную схему атмосферной циркуляции (с одной ячейкой) предполагал еще в начале XVIII века - правда, для современности - физик Дж.Гадлей, однако позже выяснилось, что атмосфера Земли организована иначе, с тремя ячейками, и о модели Гадлея забыли.
- Апвелинг - место подьема к поверхности океана глубинных вод. Последние всегда обогащены биогенами (прежде всего - фосфором), и потому в зонах апвелингов развиваются чрезвычайно высоко продуктивные морские сообщества, и море едва не кишит рыбой. Апвелинги, развивающиеся вдоль западных побережий Южной Америки и Южной Африки (там, где к этим материкам подходят рукава Циркумантарктического течения), служат главными районами морского рыболовства.
- Скорпионы представляют собою специализированную группу уже знакомых нам (по главе 7) морских ракоскорпионов-эвриптерид, представители которой перешли от плавания к хождению по дну и, приобретя мелкие размеры, освоили сначала морскую литораль, а потом и сушу.
- С находкой кембрийских морских многоножкоподобных членистоногих их существование и на раннепалеозойской суше кажется вполне вероятным, хотя достоверные находки многоножек в континентальных отложениях появляются лишь в позднем силуре.
- Не исключено, что уже в венде на суше существовали и макроскопические растения. В это время на талломах некоторых водорослей (Kanilovia) возникают загадочные сложноустроенные микроструктуры в виде зигзагообразно разрывающейся повдоль спиральной хитиноидной ленты. М.Б.Бурзин (1996) вполне логично предположил, что они служат для разбрасывания спор - а такой механизм необходим лишь в воздушной среде.
- Некоторые авторы (например, Л.С.Берг) считали почвенную жизнь вообще первичной.
- Псефиты - рыхлые осадки из обломочного материала, более грубые, чем "глина" (пелиты) и "песок" (псаммиты).
- Ни одно из высших растений не способно к азотфиксации, т.е. к преобразованию азота из атмосферного газа N2 в усваиваяемую форму (например, ионов NO3 ¯ ); это - дополнительный аргумент в пользу того, что к моменту появления на суше высших растений там уже давно существовали прокариотические сообщества, которые обогатили почву азотом в доступной форме.
- Более привычное название - псилофиты - сейчас не употребляют по номенклатурным соображениям. В литературе последних лет вы можете столкнуться с еще одним названием - проптеридофиты.
- К концу девона уже появились представители почти всех основных отделов высших растений, не только споровых (плауновидные, папоротникообразные, хвощеобразные), но и голосеменных (гинкговые).
- Широко известна истинно романтическая история открытия этого "живого ископаемого", описанная в замечательной книге Дж.Смита "Старина четвероног". Следует, правда, заметить, что образ жизни латимерии не имеет ничего общего с тем, что вели девонские рипидистии: она живет в Индийском океане на глубинах в несколько сот метров.
- Старое название, которое вы можете встретить в книгах - стегоцефалы - сейчас не употребляют.
- Не называем же мы "наземным существом" угря, который способен ночью, по росистой траве, переползти из одного водоема в другой, преодолевая расстояние в несколько сот метров!
- Палеозойские растения, сочетающие в себе признаки голосеменных и папоротникообразных (семена и расчлененную "папоротниковую" листву) раньше выделяли в отдельный класс голосеменных - птеридоспермы, или семенные папоротники. Теперь, однако, их считают искусственной, сборной группой (не "кладой" - самостоятельной эволюционной ветвью, а "градой" - уровнем развития). То есть - самые различные таксоны голосеменных прошли в своем эволюционном развитии "птеридоспермовую стадию", а разные группы "семенных папоротников" теперь включены в соответствующие классы голосеменнных (Glossopteridales - в гинкговых, Lagenostomales - в цикадовых, и т.д.)
- Маноксилические (что означает "малодревесинные") стволы состоят преимущественно из коры - в противовес пикноксилическим (состоящим преимущественно из древесины). Такие стволы не выносят промораживания, и - если только не принадлежат однолетникам (что легко определить по их толщине) - считаются свидетельством безморозного климата.
- Сферический сегмент - часть шара, отделенная секущей плоскостью.
- Интересно, что подобную схему атмосферной циркуляции (с одной ячейкой) предполагал еще в начале XVIII века - правда, для современности - физик Дж.Гадлей, однако позже выяснилось, что атмосфера Земли организована иначе, с тремя ячейками, и о модели Гадлея забыли.
- Апвелинг - место подьема к поверхности океана глубинных вод. Последние всегда обогащены биогенами (прежде всего - фосфором), и потому в зонах апвелингов развиваются чрезвычайно высоко продуктивные морские сообщества, и море едва не кишит рыбой. Апвелинги, развивающиеся вдоль западных побережий Южной Америки и Южной Африки (там, где к этим материкам подходят рукава Циркумантарктического течения), служат главными районами морского рыболовства.
- Глоссоптеридиевые - архаичные гинкговые, которые в старых классификациях относили к числу "семенных папоротников" (птеридоспермов) (см. сноску 34).
- Некоторые авторы, указывая на низкую механическую прочность огромных лепидодендроновых стволов (которые практически лишены древесины), предполагали, что они были почти полностью погружены в воду, высовывая наружу лишь верхушку - вроде современных рдестов.
- В известном фантастическом романе Обручева "Плутония" (являющем собою фактически научно-популярную книгу о вымерших животных) герои сталкиваются с огромными, размером с собаку, муравьями - "царями юрской природы". Надобно заметить, что разнообразных динозавров геолог Обручев описал вполне грамотно и реалистично, а вот исполинские муравьи - это уже чистейшая фантазия.
- Первоначально плейромейи считались гелофитами, однако потом обратили внимание на то, что эти растения почти всегда встречаются в ископаемом состоянии полностью, вместе с неповрежденной корневой системой: такой тип захоронения характерен именно для свободноплавающих форм.
- Раньше все эти формы, имеющие признаки как рептилий, так и амфибий, объединяли в особый подкласс батрахозавров, но ныне считают, что батрахозавры - сборная группа, и название это не употребляется.
- Это особенно интересно на фоне того, что ранее, в девоне, существуют находки амфибий и за пределами тропического пояса, например в Австралии. Дело в том, что девонские амфибии были еще существами чисто водными, а воде, как известно, температурные перепады между климатическими зонами сильно выровнены.
- Весьма своеобразная эндотермия возникает даже у некоторых рыб - высокоскоростных хищников вроде тунца или меч-рыбы, но как раз у рыб-то, с их двухкамерным сердцем и единственным кругом кровообращения, кровь разделена на венозную и артериальную изначально!
- У насекомых, тоже пошедших по пути изоляции покровов и экономии воды, конечным продуктом белкового обмена также является мочевая кислота.
- Ряд исследователей считает эвриапсидный тип черепа лишь модификацией диапсидного, которую разные группы водных рептилий приобретали независимо; при таком понимании "эвриапсиды" становятся искусственной, сборной группой (вроде знакомых нам "семенных папоротников").
- Синдром - сочетание признаков (симптомов), имеющих общий механизм возникновения.
- Недавно М.Ф.Ивахненко (1980) привел серьезные доводы в пользу того, что и третья, "анапсидная", эволюционная линия тоже имеет независимые амфибийные корни; на этом основании он предложил выделить черепах, котилозавров и связанные с ними формы в отдельный класс - парарептилии. Кроме того, существует ряд доводов в пользу того, что амфибии, стоящие в основании соответствующих рептилийных стволов, ведут происхождение от разных групп кистеперых; если эта гипотеза подтвердится, то окажется, что тероморфы и завроморфы разошлись даже не на "амфибийном", а еще на "рыбном" уровне!
- О теплокровности высших терапсид может свидетельствовать и ряд косвенных признаков. В это время впервые появляются и подземные (роющие) животные, и ночные (с очень крупными глазами и бинокулярным зрением, как у лемура или совы) - а такой образ жизни для пойкилотермного существа, нагревающегося "от солнышка", вряд ли возможен.
- Белые грудные мышцы куриных как раз и обеспечивают резкий энергичный взлет.
- Картина из "Парка юрского периода" - тиранозавр, преследующий героев вплавь - видимо вполне соответствует действительности.
- Окончательно эту проблему удалось решить лишь на уровне высших (териевых) млекопитающих; даже однопроходные млекопитающие (относимые к прототериям) сохраняют "рептильные" черты в строении плечевого пояса (неслитая с лопаткой коракоидная кость).
- Cейсмозавр (известный лишь по отпечаткам следов) должен был достигать в длину 48 метров.
- Недавно описанный птеранодон Quetzalcoatlus (от которого, правда, известны лишь несколько изолированных остатков) должен был достигать в размахе крыльев 16 м.
- У позднемелового птеродактиля Pterodaustro в нижней челюсти были сотни длинных щетинковидных зубов - полный аналог "китового уса", позволяющего отцеживать из воды мелких беспозвоночных.
- Высказывалась достаточно обоснованная гипотеза, что зауроподы обитали на морских мелководьях (чему есть прямые доказательства в виде их следов на известковом морском песке с остатками морских моллюсков). Они проводили время, стоя на дне, на глубине 3-4 метров, и питались водорослями; отсюда понятно, зачем им такие чудовищные размеры тела - просто чтобы не сбивало с ног волнами.
- Собирательное название элементов (медь, свинец и т.п.), соли которых высокотоксичны для высших животных.
- К фитофагии пытались перейти и некоторые прототерии (юрские мультитуберкуляты, несколько сходные с грызунами), однако эти попытки, как видно, не увенчались успехом.
- Ситуация, видимо, сходная с современными крокодилами: те активно охраняют кладку, но немедленно расстаются с вылупившимися детенышами.
- Ныне от этой группы уцелели лишь осетровые и некоторые экзотические формы вроде африканских многоперов и североамериканских панцирных щук.
- Единственные исключения - гнетум и вельвичия в первом случае и эфедра - во втором, но эти растения по своей жизненной форме являются имитаторами цветковых: у них есть широкий мягкий лист, ягоды и т.п. В некоторых новейших классификациях их даже стали включать в состав цветковых.
- Под сукцессией здесь подразумевается не всякая смена растительности, а то, что С.М.Разумовский (1982) называет экогенетической сукцессией: жестко детерменированная последовательность растительных ассоциаций (сериальный ряд), каждая из которых видоизменяет состав почвы так, что он становится более благоприятен для ассоциации-последователя, нежели для нее самой, и приводит в конечном итоге к ассоциации-климаксу, в почвах которой достигнут "нулевой баланс" по органике; климаксная ассоциация не имеет внутренних причин к смене и может существовать неопределенно долго - пока не будет разрушена извне (например, лесным пожаром). Пример экогенетической сукцессии для Подмосковья - зарастание гари иван-чаем (пионерная стадия), березняк, смена его ельником и, далее, дубравой (климаксом); цикл этот длится около 300 лет. Совокупность климакса с ведущими к нему сериальными рядами называют сукцессионной системой; ее можно рассматривать как гомеостат, который имеет равновесное состояние (климакс), а будучи же выведен из него внешними силами, возвращается в исходное положение при помощи ряда стандартизованных реакций (экогенетических смен).
- В миоцене был момент (мессинский кризис), когда Средиземное море почти полностью высохло, а дно его заросло саваннами африканского типа.
- Полидоминантный лес - слагающийся большим числом лесообразующих пород. Широко известно такое сопоставлениие: на одном-единственом гектаре амазонской гилеи можно насчитать больше видов деревьев, чем во всей Англии.
- Когда в Австралию завезли домашних копытных, оказалось, что местные навозники не справляются с утилизацией их помета и накопление неразложенных экскрементов на пастбищах останавливает возобновление трав; пришлось завозить туда европейских навозников - тогда все пришло в норму.
- "Южноамериканские копытные" - шесть вымерших отрядов, эндемичных для этого материка. Все они, как и наши парнокопытные и непарнокопытные, ведут происхождение от кондиляртр, однако с позднего мела развивались в изоляции, и сходство с различными "северными" группами приобрели независимо, то есть чисто конвергентно.
- Это были одни из самых своеобразных копытных: пропорции конечностей у них таковы, что локомоция их была "полудвуногой", как у гориллы, а на ногах у них вторично возникли крупные невтяжные когти. Считают, что они обгладывали листву деревьев, пригибая к себе ветви при помощи "рук".
- Именно это существо было описано И.А.Ефремовым в романе"Великая Дуга" под названием "гишу - ужас ночей, пожиратель слонов".
- Лошади (из непарнокопытных) и полорогие (из парнокопытных) являют собою две вершины в эволюции травоядности. У обоих имеется в пищеварительном тракте "бродильная камера", где симбиотические микроорганизмы дополнительно ферментируют пережеванную траву, однако у полорогих это рубец в передней части желудка, а у лошадей - слепая кишка позади него. Полорогие перерабатывают пищу медленно и неторопливо, извлекая из нее максимум того, что возможно, а лошади, напротив, прогоняют травяную массу через желудок с предельной быстротой. Наблюдения в африканских саваннах показали, что лошади (зебры), избирательно питаясь наиболее грубоволокнистыми травами, делают остальные типы корма более доступными для полорогих (антилоп).
- Мне не хотелось бы вступать здесь в полемику с авторами наводнивших Россию "религиозно-просветительских" брошюр. Отсылаю их всех "по принадлежности" - к Папе Римскому, точнее - к папской энциклике, написанной в 1967 году после Второго Ватиканского Собора и признающей, что теория Дарвина "верно трактует вопросы происхождения человеческого тела".
- Седиментогенез - процесс накопления и окончательного осаждения осадков в водной среде.
- История эта весьма назидательна в том смысле, что факт наличия у некоторой естественнонаучной гипотезы математической модели (например, как в знакомых нам "импактных" гипотезах мелового вымирания) не должен гипнотизировать естествоиспытателя; математика - всего лишь инструмент (как та астролябия Остапа Бендера, которая "сама меряет... Было б только чего мерить"), каковой сам по себе истинности не гарантирует.
- То, что ледник возникает не столько "от холода", сколько "от сырости", известно достаточно давно. Обратившись к карте 58, легко видеть, что покровное оледенение достигало максимума в теплых и влажных приатлантических областях, тогда как в сухой и холодной Восточной Сибири и на Аляске существовали лишь локальные горные ледники.
- Необходимо помнить, что появление на территории лесной зоны этих степных (т.е. вроде бы южных) видов маркирует не потепления, как иногда ошибочно полагают зооеографы, а именно ледниковые эпохи.
- Существующие на Земле пустыни делят по динамике выпадения осадков на три типа. Береговые пустыни развиваются там, где к жарким побережьям подходят холодные морские течения (Намиб, Атакама): здесь осадков, считай, вовсе нет; жизни, соответственно, тоже. Пустыни центральноазиатского типа (Гоби, Бетпак-Дала): в них осадки "равномерно размазаны" по всему году - и потому жизнь тут есть весь год, но едва теплится. Пустыни средиземноморского типа (Сахара, Кара-Кумы, Большая Песчаная пустыня в Австрали): здесь осадков столько же, что и в предыдущем типе, но только все они выливаются разом, за две-три недели; тут происходит краткий и бурный расцвет жизни (разнообразные эфемеры), которая затем переходит в латентное состояние - до следующего года.
- Дань, которую английские короли в X-XI веках были вынуждены выплачивать викингам, регулярно вторгавшимся на Британские острова.
– Конец работы –
Используемые теги: Еськов, История, земли, жизни0.065
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: К.Ю. Еськов История Земли и жизни на ней
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.182 сек.
Новости и инфо для студентов