Биоэнергетический прогресс - раздел История, А.И.Зотин, А.А.Зотин НАПРАВЛЕНИЕ, СКОРОСТЬ И МЕХАНИЗМЫ ПРОГРЕССИВНОЙ ЭВОЛЮЦИИ Как Отмечалось, Согласно А.н.северцову (1925, 1934, 1939) Морфо-Физиологическ...
Как отмечалось, согласно А.Н.Северцову (1925, 1934, 1939) морфо-физиологический прогресс осуществляется через ароморфозы приспособительные изменения организмов, в результате которых общая энергия жизнедеятельности взрослых потомков повышается. Хотя определение ароморфозов страдает некоторой неопределенностью, оно содержит в себе основу,
на которой можно построить количественные критерии эволюционного прогресса, пригодные для сравнительно-эволюционных исследований. В частности, Ивлев (1959, 1963) и Дольник (1968,1878) конкретизировали представления Северцова о повышении общей энергии жизнедеятельности организмов, идентифицировав ее с повышением в ходе прогрессивной эволюции стандартного или основного обмена. Стандартный обмен (потребление кислорода в состоянии покоя при 20°С у пойкилотермных животных и в термонейтральной зоне - у гомойотермных) как и другие виды энергетического обмена, связан с массой тела, так называемыми, аллометрическими соотношениями вида

|
| потребления кислорода; M - масса организма (в г); a, k и b - коэффициенты, причем b = - k. При сопоставлении стандартного обмена у разных видов нельзя сравнивать животных, имеющих разную массу, так как из формулы (47) следует, что интенсивность потребления кислорода уменьшается по мере увеличения массы животного (коэффициент b y всех организмов больше нуля и меньше единицы). Следовательно, у более крупных животных интенсивность дыхания всегда ниже, чем у мелких, даже если эти животные относятся к одному семейству или классу. Сравнивать энергетический метаболизм мыши и слона по интенсивности дыхания бессмысленно, так как масса тела у них отличается на 5 порядков, не говоря уж о сравнении энергетического обмена слона и инфузории, когда разница в массе достигает 15 порядков.
|
Исходя из этого, Ивлев (1959) в качестве основного показателя энергии жизнедеятельности животных предложил использовать коэффициент а из аллометрических соотношений (46) и (47), который соответствует интенсивности дыхания организма массой в 1 г. Эта величина, конечно, условная, так как слоны, так же как и инфузории, не бывают массой в 1 г, но иного выхода для сравнения стандартного обмена разных видов животных или растений нет.
Попытка Ивлева использовать биоэнергетический показатель для характеристики прогрессивной эволюции животных не удалась (Ивлев, 1963). Более удачливым в этом отношении оказался Дольник (1968). Используя данные о стандартном обмене самых разнообразных групп животных, он показал, что коэффициент а заметно возрастает по мере увеличения сложности организации животных от простейших до млекопитающих и птиц (табл. 1).
Таблица 1.
Диапазон массы тела и стандартный обмен [константы а и k из аллометрического соотношения (46)1 в разных классах и типах животных (Дольник, 1968).
| Типы и классы
| Масса, г
| A
| k
| Авторы
|
| Protozoa
| 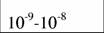
| 0,098
| 0,75
| (1)*
|
| Anthozoa
| 
| 0, 19
| 0,86
| (2)
|
| Oligochaeta
| 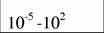
| 0,54
| 0,61
| (3)
|
| Xiphosura
| 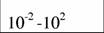
| 0,47
| 0,81
| (2)
|
| Crustacea
| 
| 0,93
| 0,81
| (4)
|
| Mollusca
| 
| 1,15
| 0,75
| (2)
|
| Insecta
| 
| 3,15
| 0,67
| (5)
|
| Acrania
|
| 2,13
| 0,91
| (6)
|
| Cyclostomata
| 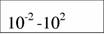
| 1,70
| 0,80
| (7)
|
| Pisces
| 
| 1,71
| 0,81
| (7)
|
| Amphibia
| 
| 1,45
| 0,67
| (8)
|
| Reptilia
| 
| 4,60
| 0,67
| (8)
|
| Mammalia
| 
| 21,4
| 0,73
| (9)
|
| Mammalia
|
| 18,0
| 0,76
| (10)
|
| Aves
| 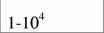
| 51,9
| 0,64
| (И)
|
| Aves
| 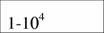
| 21,1
| 0,74
| (12)
|
| Aves
| 
| 40,9
| 0,66
| (12)
|
*(1) Hemmingsen, 1960; (2) Дольник, 1968; (3) Ивлев, 1963; (4) Винберг, 1950; (5) Wigglsworth, 1953; (6) Ивлев, 1961; (7) Винберг, 1956; (8) Brown, 1964; (9) Brody, 1945; (10) Kleiber, 1947; (11) Brody, Proctor, 1932; (12) King, Farner, 1961.
Позднее данные, приводимые Дольником, были уточнены (Коноплев, Зотин, 1975; Коноплев и др., 1975; Зотин, Криволуцкий, 1982; Зотин, 1979, 1984; Zotin, 1990), но основной вывод, сделанный этим автором, что коэффициент а для крупных таксонов порядка класса и типа возрастает по мере усложнения организации животных, полностью подтвердился. Больше того, оказалось, что при сопоставлении стандартного обмена (коэффициента а) той или иной группы животных (табл.1) и времени обнаружения этой группы в палеонтологической летописи (Зотин, 1979; 1984; Зотин, Криволуцкий, 1982), выявляется явная тенденция усиления энергетического обмена по мере протекания биологической эволюции (рис.6). Как видно из этого рисунка, в процессе эволюции происходило последовательное появление животных со все более высоким уровнем стандартного обмена, т.е. увеличивались энергетические возможности организмов. Таким образом, хотя критерий эволюционного прогресса, использованный Ивлевым и Дольником, не получил широкого признания (особенно среди
профессиональных эволюционистов), нет сомнений, что он может быть положен в основу количественной характеристики такого направления прогрессивной эволюции, как биоэнергетический прогресс.
По предположению Дольника (1968) прогрессивная эволюция равнозначна биоэнергетическому прогрессу. На этом положении мы подробнее остановимся во второй части данной книги, здесь же рассмотрим представления двух других авторов, рассматривающих прогрессивную эволюцию с позиций биоэнергетики. Кратко они сводятся к следующему.
И.И.Свентицкий (1977, 1980, 1981) предлагает ввести новый постулат: принцип биоэнергетической целенаправленности, согласно которому
"каждый элемент живой природы в своем развитии и функционировании стремится к состоянию, обеспечивающему в существующих условиях внешней среды наиболее полное использование свободной энергии системой данного трофического уровня, в которую этот элемент входит. " Автор приходит к выводу, "что в процессе эволюции использование свободной энергии каждой нишей непрерывно возрастает как за счет увеличения размеров популяции, использующих данный вид свободной энергии, так и путем повышения их биоэнергетического коэффициента полезного действия" (Свентицкий, 1980, С.7).
К сходным выводам пришел и Н.С.Печуркин (1982). Он формулирует энергетический принцип экстенсивного развития биологических систем в виде двух соотношений
(где H е - поток энергии среды, который может быть использован живыми организмами) и энергетический принцип интенсивного развития, согласно которому в процессе эволюции возрастает удельная используемость свободной энергии. "Согласно обсуждавшимся энергетическим принципам, магистральное направление эволюции существует, и оно заключается в увеличении способности захватывать и использовать потоки свободной энергии..." (Печуркин, 1982, С. 106). Реализовать эти принципы живые системы могут при освоении новых ареалов или при резких изменениях условий существования вида за счет быстрого увеличения численности популяции. Подтверждение этим принципам автор находит в экспериментах с микробными популяциями в проточных культурах и в теории ароморфозов (Печуркин, 1982).
Очевидно, что данные, приводимые на рис.6 подтверждают принципы, высказанные указанными выше авторами, как, и идею Шноля (1973, 1979) о возрастании кинетического совершенства в процессе биологической эволюции. Мы же, как будет показано дальше, даем этому явлению иное объяснение, основанное на постулатах термодинамики неравновесных процессов.
Следует также отметить, что принципы Свентицкого и Печуркина призваны объяснить направленность эволюционных процессов и особенно направленность эволюционного прогресса. При этом, хотя в приведенных формулировках они практически не отличаются друг от друга, в методологическом плане они резко различны. Печуркин считает, что эволюция организмов идет не сама по себе, а направляется потоком внешней свободной энергии, создаваемым в конечном счете Солнцем. "Именно поток энергии организует систему (круговорот) и заставляет ее эволюционировать" (Печуркин, 1982, С.46). Наоборот, Свентицкий (1977) включает в свой принцип биоэнергетическую целенаправленность, что подразумевает наличие внутренней причины эволюционного прогресса. Таким
образом, можно проводить аналогию, с одной стороны, между взглядами Печуркина и теорией ортогенеза Эймера (Eimer, 1898; Завадский, 1973), согласно которой направленность эволюционного прогресса зависит от причин, лежащих во внешней среде (климат и пища), с другой - между высказываниями Свентицкого и принципом градаций Ламарка (1937), принципом усовершенствования Нэгели (Nageli,1884), батмизма Копа (Соре, 1896), номогенеза Л.С.Берга (1922, 1977) и другими ламаркистскими теориям, в которых направленность эволюционных процессов и особенно прогрессивной эволюции объясняют целестремительными принципами или законами (Завадский, 1973; Филипченко, 1977; Назаров, 1984).
Между тем, нет необходимости возводить особые эволюционные принципы в виде стремления организмов как можно полнее использовать солнечную энергию или свободную энергию пищи. В этом отношении, как нам кажется, прав Викен (Wicken, 1980), который считает, что использование свободной энергии, идущей от Солнца, осуществляется через селекционизм, а не как прямой ответ на градиент свободной энергии. Градиент солнечной энергии создает как бы особую нишу, заполнение которой чрезвычайно выгодно организмам в борьбе за существование. Действительно, повышение интенсивности энергетического обмена за счет использования свободной энергии, имеющейся в среде, является большим преимуществом организмов, осуществивших это повышение, так как это увеличивает мощность и быстроту реакций организмов, возможность освоения новых географических районов и экологических ниш, увеличивает устойчивость и надежность физиологических процессов, протекающих в организмах или осуществляемых организмами. И здесь не требуется введение особых принципов, как это предложили Свентицкий и Печуркин - естественный отбор сам направляет эволюцию, когда это возможно, по пути все большего использования свободной энергии для повышения уровня энергетического обмена и создает направленность
эволюционного процесса, который А.Н.Северцов называл морфо-физиологическим прогрессом.
Это не означает того, что механизмы такого макроэволюционного процесса, как морфо-физиологический прогресс, полностью определяются механизмами микроэволюции. Арогенез, в соответствии с определением ароморфозов, из которых складывается арогенез, состоит в увеличении общей энергии жизнедеятельности организмов и, следовательно, его осуществление возможно только путем изменения интенсивности энергетического обмена. Последнее происходит в организмах только при определенных условиях, о которых будет подробно сказано дальше. Стоит только отметить, что в конечном счете механизмы эволюционных изменений интенсивности энергетического обмена организмов не осуществимы без участия микроэволюционных процессов. А это означает, что понимание механизмов прогрессивной эволюции базируется на теории естественного отбора.
И все же не следует забывать, что согласно термодинамике неравновесных процессов при эволюции сложных систем к равновесию действует принцип наискорейшего спуска, который предполагает появление в системе подсистем, двигающихся в сторону увеличения неравновесности, что и происходит в процессе прогрессивной эволюции в отдельных группах животных и растений (Зотин, Зотин, 1995).
Все темы данного раздела:
Часть 1. ТЕРМОДИНАМИЧЕСКИЕ ОСНОВЫ ЭВОЛЮЦИИ
Главный вопрос, который встает при рассмотрении биологических явлений, даже такого масштаба как процесс эволюции организмов, сводится в самом общем виде к следующему: является ли это явление естест
Вероятностная интерпретация критериев эволюции.
Критерии эволюции термодинамики неравновесных процессов (12,13,18), как и критерий классической термодинамики (6), показывают не только направление изменений термодинамической системы, но и определ
Кинетика приближения термодинамической системы к конечному состоянию.
Сначала ограничимся случаем, когда выполняется уравнение (23), т.е. система находится вблизи от стационарного состояния. Решая это уравнение, получаем
Микроэволюция
Микроэволюцией называют процессы, протекающие в популяциях и заканчивающиеся образованием нового вида (Тимофеев-Ресовский и др., 1969; Северцов, 1987; Яблоков, Юсуфов, 1998;). При образовании новог
Аксиоматика
Аксиоматике эволюционного учения посвящено довольно много работ (Воронцов, 1980, 1984; Медников, 1982, 1984, 1989; Jongeling, 1985). Так как в данном разделе книги речь идет только о доказательства
Синтетическая теория эволюции
Во времена Уоллеса и Дарвина еще не было четких представлений о механизмах наследования. Поэтому одним из самых коварных возражений против теории естественного отбора был, так называемый, парадокс
Макроэволюция
Макроэволюцией называют процессы, приводящие к возникновению таксонов надвидового ранга: родов, семейств, классов и т.д. (Тимофеев-Ресовский и др., 1969; Северцов, 1987, 1990). Макроэволюция охваты
Биологический и морфо-физиологический прогресс
Еще со времен Ламарка (1935, 1937) возникли представления о направленности процесса эволюции. Последующее развитие биологии и особенно многочисленные палеонтологические находки показали, что с ходо
Прогрессивная эволюция
Наличие эволюционного прогресса в живой природе не вызывает сомнений прежде всего из-за наличия палеонтологических данных, показывающих что в реальной эволюции происходило появление все более совер
Совершенствование регуляции и управления
Как отмечают многие авторы, эволюционный прогресс сопровождается не только увеличением интенсивности энергетического обмена, но и совершенствованием процессов регуляции и управления. Грант (1980, 1
Мегаэволюция
Выше мы рассматривали такие этапы эволюции, как микроэволюция и макроэволюция. Теперь остановимся на вопросах, связанных с проблемами мегаэволюции.
Мегаэволюцией Симпсон (1948), как и неко
Механизмы мегаэволюции.
Свойства биосферы определяются входящими в ее состав биоценозами (Тимофеев-Ресовский и др., 1969), поэтому ее эволюционные изменения должны начинаться с изменения структуры биоценозов и особенно би
Регрессивная эволюция.
Биосфера в целом и отдельные ее биологические составляющие (популяции, биоценозы, экосистемы) представляют собой термодинамические, открытые, организованные системы. Они должны, поэтому, подчинятьс
Энтропия и негэнтропия
По поводу применимости термодинамики для описания биологических систем и процессов в научной периодике время от времени возникали (Беркинблит, 1965; Быховский, 1965; Сырников, 1965; Popper, 1965, 1
Энтропия.
Понятие энтропии - одно из основных понятий термодинамики, без использования которого невозможно термодинамическое рассмотрение ни равновесных, ни неравновесных процессов (Зарафянц, 1935; Шамбадаль
Негэнтропия.
Термин негэнтропия введен в термодинамику Бриллюэном (Brilloun, 1949, 1951; Бриллюэн, 1960, 1962) как меры порядка системы. С помощью этого понятия он попытался связать термодинамику, статистическу
Организованные системы
Бюргард (Beauregard Costa, 1960) отмечает, что термин информация в утверждении (48) употребляется в двух смыслах: в первом случае: негэнтропия —> информация - речь идет о собирании сведений, а в
Живые системы.
В живых системах, которые можно определить, как открытые, далекие от равновесия, организованные системы, способные к размножению, всегда имеют место негэнтропийные эффекты. Они заключаются в том, ч
Термодинамические подходы к проблемам биологической эволюции
Термодинамический подход к проблеме эволюции и подход с точки зрения естественного отбора резко отличаются друг от друга и поэтому долгое время эти две теории эволюции практически не соприкасались.
Термодинамика и направленность эволюции
По вопросу о направленности эволюции существует не только большая литература, но и длительная, почти 200-летняя, дискуссия между ламаркистами и дарвинистами, конца которой не видно (Назаров, 1984;
Общие соотношения биологической термодинамики
В обычных условиях покоя организмы находятся в устойчивом состоянии с минимальной интенсивностью потребления кислорода, соответствующему основному или стандартному обмену. Под воздействием внешних
Часть 2. ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ: УСИЛЕНИЕ ЭНЕРГЕТИЧЕСКОГО ОБМЕНА
Наличие в живой природе более совершенных и менее совершенных организмов никогда не вызывало сомнений и было очевидным задолго до появления эволюционного учения (Лункевич, 1936, 1940, 1943; Воронцо
Стандартный обмен
Перейдем теперь к характеристике основных процессов, входящих в понятие энергетического обмена. Это прежде всего основной и стандартный обмен. Под основным обменом имеют в виду интенсивность
Значение массы тела
Прежде всего, следует учитывать то, что стандартный обмен, как и другие виды энергетического обмена, связан с массой тела животных аллометрическими соотношениями (46) и (47). Из этих соотношений сл
Влияние температуры
Стандартный обмен, как и другие биохимические процессы, протекающие в организмах, сильно зависит от температуры среды, поэтому принято измерять потребление кислорода у пойкилотермных животных при 2
Акклимация и акклиматизация
Упомянутые неточности определения зависимости стандартного обмена от температуры, помимо естественного разброса данных о потреблении кислорода животными (зависящего от состояния животного, времени
Критерий упорядоченности
Как уже говорилось, согласно предположению Ивлева (1959, 1963) и Дольника (1968, 1978) арогенез состоит в усилении энергетического обмена организмов: усилении основного или стандартного обмена. Это
Максимальный обмен
Как отмечалось, понятие стандартный обмен (обмен покоя) недостаточно четко определено, так как в состоянии покоя активность разных животных различна и, измеряя потребление кислорода в таком состоян
Константа Рубнера
Еще один критерий эволюционного прогресса, который может быть получен в цифровом выражении, предложен Э.С.Бауэром (Bauer, 1931; Бауэр, 1935) и назван им константой Рубнера. Этот критерий осн
Простейшие
Хотя к настоящему времени накоплено достаточно много материалов по дыханию простейших, работ, обобщающих и систематизирующих эти данные, сравнительно немного (John, 1941; Винберг, 1949; Zeuthen, 19
ApicomplexaSarcomastigophoraCiliophora
Расположение классов в типе Sarcomastigophora:
Eumycetozoea
Низшие беспозвоночные
Для большинства типов и классов беспозвоночных показана аллометрическа зависимость (46). Однако для многих типов низших беспозвоночных из-за ограниченного числа измеренных видов животных точность о
Черви - Plathelminthes (Плоские черви)
Из группы червей мы остановимся только на типе Plathelminthes, но при вычислении зависимости дыхания от температуры учитывались данные, полученные для аннелид. Хотя данных по влиянию температуры на
Высшие беспозвоночные
К высшим беспозвоночным животным относят типы членистоногих (Arthropoda), моллюсков (Mollusca), иглокожих (Echinodermata), щетинкочелюстных (Chaetognatha), оболочников (Tunicata) и полухордовых (He
Acariformes —>Amblypygi —> Scorpiones —> Parasitiformes —> —> Aranei —> Solifugae —> Opiliones
Таблица 11
Коэффициент а (мВт/г) и критерий упорядоченности Сr в разных таксонах класса Arachnida (Паукообразные). N - число видов, n - число измерений (Зотин и др., 1998а).
Низшие насекомые - Apterygota
Данных о дыхании низших насекомых, к которым относят 4 отряда (Плавильщиков, 1957), немного, кроме, может быть, представителей отряда ногохвосток (Бызова, 1972). Согласно ее обзору два вида из отря
Высшие насекомые - Pterygota
К высшим насекомым относят около 25-30 отрядов класса Insecta (Плавильщиков, 1957; Бей-Биенко, 1980). В известных нам обзорах об интенсивности дыхания взрослых насекомых приводится сравнительно нем
Насекомые с неполным превращением
Отряд Blattodea(Таракановые).
Данных о дыхании тараканов сравнительно немного. В работе Донцовой и др.(1985) приведены собственные и литературные материалы о массе тела и
Насекомые с полным превращением
Отряд Coleoptera (Жесткокрылые).
Для этого отряда имеются данные о массе тела и дыхании (117 видов насекомых, 133 измерений) из 19 семейств (Алексеева, Зотин, 1995). Авторы приводят следую
Суперорганизмы.
Совсем иные значения коэффициента а и критерия упорядоченности Сr получаются, если рассматривать суперорганизмы из отряда перепончатокрылых. Известно, что многие пчелы (Moritz, Southw
Макросистематика насекомых
Класс насекомых включает от 24 до 30 отрядов (классификация класса находится еще в процессе становления и точное число отрядов не установлено). Из них мы будем говорить только о 12 наиболее распрос
Taxodonta —> Eulamellibranchia —> Dysodonta
При определении критерия упорядоченности принимали, что моллюски умеренного пояса дышат в 1.66 раз сильнее, чем тропического. В этом случае получаем для класса двустворчатых моллюсков:
Таб
Gymnosomata —> Sacoglossa —> Nudibranchia—> —> Cephalaspidae
В целом для класса брюхоногих моллюсков имеем: Класс Gastropodaa=0.685±0.032; Сr=13.86±0.70 (112)
Класс Cephalopoda(Головоногие)
Головоногие
Bivalvia—> Gastropoda—> Loricata—> Cephalopoda
Тип Echinodermata (Иглокожие)
Работ, в которых изучалась скорость потребления кислорода взрослыми формами иглокожих сравнительно немного. Больше других в э
Echinoidea —>Asteroidea —> Holothuroidea —> —> Ophiuroidea—> Crinoidea
Таблица 31.
Коэффициент а и критерий упорядоченности Сr для разных классов типа Echinodermata.По данным, представленным в табл.30. N - число изученных видов.
&nbs
Caudata -> Gymnophiona -> Anura
Так же располагают отряды амфибий некоторые другие авторы (Наумов, 1950; Матвеев, 1956), но обычное расположение этих отрядов следующее: Gymnophiona - Caudata - Anura (Брэм, 1931; Кашкаров, Станчин
Testudintes —> Rhynchocephalia —> Squamata —> Crocodylia
В настоящее время еще преждевременно обсуждать вопрос о том правильно ли расположены семейства пресмыкающиеся в каждом подотряде согласно табл.36, так как экспериментальных данных о стандартном обм
Gt; Chiroptera —> Primates —> Cetacea —> Perissodactyla
Последовательность расположения отрядов по этой схеме отличается не только от предложенной другими авторами (табл.38), но и макросистемы отрядов млекопитающих, описанной в работе Зотина и Владимиро
Изменение энергетического обмена в ходе эволюции
Как отмечалось выше, вывод Дольника (1968), согласно которому по мере усложнения организации животных стандартный обмен заметно возрастает, подтверждается всеми известными нам данными. Чтобы сделат
Биологический и физический смысл эволюционного прогресса
Как видно из сказанного выше, в эволюционном прогрессе организмов мы различаем биоэнергетический прогресс, измеряемый с помощью коэффициента а [(46), (47)], и собственно прогрессивную эволюц
Направление и скорость прогрессивной эволюции и биоэнергетического прогресса
Хотя критерий эволюционного прогресса, использованный Ивлевым и Дольником, пока не получил широкого признания (особенно среди профессиональных эволюционистов), нет сомнений, что он может быть полож
Направление и скорость эволюционного прогресса в соответствии с константой Рубнера
Как отмечалось, константа Рубнера менее удобна для характеристики эволюционного прогресса, чем критерий упорядоченности или коэффициент а. В ее состав входят не только стандартный обмен, но
Аллогенез и катагенез
В данной работе мы в основном придерживаемся теории А.Н.Северцова относительно биологического прогресса. Необходимо, поэтому, разобраться в том, как изложенные выше представления соотносятся с этой
Часть 3. ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ: СОВЕРШЕНСТВОВАНИЕ РЕГУЛЯЦИИ И УПРАВЛЕНИЯ
"Да сохранит меня небо от глупого Ламарковского стремления к прогрессу"
"Знаю, что едва ли возможно определить
Ясно, что разумеется под более высокой
Или б
Критерии организованности
Негэнтропийные эффекты. Прежде всего, можно говорить, в принципе, об использовании для эволюционных проблем понятия негэнтропийного эффекта
Совершенствование терморегуляции в процессе прогрессивной эволюции
Как отметил Слоним (1982а), в филогенезе млекопитающих происходит совершенствование температурного гомеостаза "по мере восхождения по
эволюционной лестнице". Эта идея была п
Адаптация к условиям существования.
Очевидно, что в северных широтах необходима более совершенная терморегуляция, так как здесь имеются четкие сезонные изменения температуры и увеличивается разность между температурой тела животных и
Скорость совершенствования терморегуляции у млекопитающих в процессе эволюции.
Теперь вернемся к обсуждению вопроса о совершенствовании температурного гомеостаза "по мере восхождения по эволюционной лестнице". Данные, приводимые в табл.46, позволяют расположить отря
Gt; Lagomorpha -> Perissodactyla
Указанное расположение отрядов не очень сильно отличается от макросистематики отрядов, основанной на использовании критерия упорядоченности в классе млекопитающих (рис.26). Выделяются, пожалуй, тол
Суперорганизмы.
Помимо гомойотермных животных, к терморегуляции способны и, так называемые, суперорганизмы. К ним относятся сообщества некоторых общественных насекомых: муравейники, термитники, рои пчел, а
Эволюция максимальной продолжительности жизни животных.
Некоторые авторы указали на то, что в процессе биологической эволюции максимальная продолжительность жизни животных заметно возрастает (Sacher, 1975; Аринчин, 1982; Culter, 1984). Особенно наглядно
Зависимость массы мозга от массы тела.
Многие авторы изучали зависимость массы головного мозга от массы тела позвоночных животных (Friedenthal, 1910; Bonin, 1937; Crile, Quiring, 1940; Stephan, Andy, 1964; Jerison, 1970, 1973, 1988; Ste
Зависимость массы мозга от потребления кислорода
Как сказано, масса мозга аллометрически зависит от массы тела животных (171). С другой стороны, хорошо известна в сравнительной физиологии аллометрическая зависимость (46) между стандартным обменом
Изменение организованности животных в процессе эволюции
Как видно из приведенных данных, по некоторым показателям мозга (коэффициент энцефализации, энергетика мозга и коры головного мозга), а также по максимальной продолжительности жизни и совершенству
Часть 4. МАКРОСКОПИЧЕСКИЕ МЕХАНИЗМЫ БИОЭНЕРГЕТИЧЕСКОГО ПРОГРЕССА
Большинство отечественных эволюционистов не видят необходимости в постановке вопроса о специальных механизмах макроэволюции, отличных от микроэволюционных (Завадский, Колчинский, 1977; Иорданский,
Адаптация к низким температурам среды
Крог (Krogh, 1916) высказал парадоксальную идею, согласно которой стандартный обмен арктических животных выше, чем тропических. Эта идея породила большое число исследований энергетического обмена у
Адаптация к высокогорью
Адаптация к горным условиям жизни, куда постепенно могут быть вытеснены виды, не выдерживающие конкуренции за жизнь, связана с акклимацией и акклиматизацией не только к низкому содержанию кислорода
Адаптация к физическим нагрузкам
В условиях борьбы за существование в системе хищник-жертва или при недостаточной обеспеченности пищей выживание животных связано с интенсивной мышечной работой: в этих условиях многие виды животных
Часть 5. БИОЭНЕРГЕТИЧЕСКИЙ ПРОГРЕСС И ТЕПЛОВЫЕ БАРЬЕРЫ
В процессе прогрессивной эволюции происходит непрерывное увеличение стандартного обмена животных от простейших до млекопитающих и птиц (табл.41; рис.28). Встает, однако, вопрос, возможно ли дальней
Способны ли крокодилы и динозавры к терморегуляции?
Представленная гипотеза возникновения терморегуляции открывает новые возможности для установления времени появления гомойотермии в процессе эволюции позвоночных животных, определения уровня
Увеличение относительных размеров мозга
Многие авторы справедливо указывают, что прогрессивная эволюция характеризуется не только усилением энергетики организмов, усложнением их строения и функционирования, но и совершенствованием информ
Виртуальные цивилизации.
Следует отметить, что человеческая цивилизация могла и не возникнуть, так как, по всей видимости, первыми среди позвоночных животных встали на путь увеличения относительных размеров мозга представи
Социальная деятельность животных
Выше говорилось о предпосылках возникновения цивилизаций на базе увеличения относительных размеров мозга и связанной с этим разумной деятельностью. Но возможно, что это не единственный биологически
Ноосфера как следствие принципа наискорейшего спуска
Земная цивилизация является порождением человеческого разума и рук. Вопрос состоит в том, имела ли она биологические предпосылки? Не в смысле истории возникновения человека или отряда приматов, а в
Принцип наискорейшего спуска, как основа эволюции Вселенной
Согласно теории тепловой смерти Клаузиуса, которая вытекает из второго начала термодинамики, Вселенная движется к состоянию равновесия. Логично было бы предположить, что это движение осуществляется
Третий тепловой барьер
Как отмечалось, помимо первого теплового барьера, который возникает в процессе прогрессивной эволюции и приводит к появлению гомойотермных животных, и второго теплового барьера, преодоление которог
ЗАКЛЮЧЕНИЕ
Настоящая монография завершает серию работ, в которых сделана попытка построить термодинамические основы теоретической биологии. Эта попытка длится более 30 лет и начиналась работой «Thermodynamic








Новости и инфо для студентов