Биоэнергетический прогресс
Как отмечалось, согласно А.Н.Северцову (1925, 1934, 1939) морфо-физиологический прогресс осуществляется через ароморфозы приспособительные изменения организмов, в результате которых общая энергия жизнедеятельности взрослых потомков повышается. Хотя определение ароморфозов страдает некоторой неопределенностью, оно содержит в себе основу,
на которой можно построить количественные критерии эволюционного прогресса, пригодные для сравнительно-эволюционных исследований. В частности, Ивлев (1959, 1963) и Дольник (1968,1878) конкретизировали представления Северцова о повышении общей энергии жизнедеятельности организмов, идентифицировав ее с повышением в ходе прогрессивной эволюции стандартного или основного обмена. Стандартный обмен (потребление кислорода в состоянии покоя при 20°С у пойкилотермных животных и в термонейтральной зоне - у гомойотермных) как и другие виды энергетического обмена, связан с массой тела, так называемыми, аллометрическими соотношениями вида

|
| потребления кислорода; M - масса организма (в г); a, k и b - коэффициенты, причем b = - k. При сопоставлении стандартного обмена у разных видов нельзя сравнивать животных, имеющих разную массу, так как из формулы (47) следует, что интенсивность потребления кислорода уменьшается по мере увеличения массы животного (коэффициент b y всех организмов больше нуля и меньше единицы). Следовательно, у более крупных животных интенсивность дыхания всегда ниже, чем у мелких, даже если эти животные относятся к одному семейству или классу. Сравнивать энергетический метаболизм мыши и слона по интенсивности дыхания бессмысленно, так как масса тела у них отличается на 5 порядков, не говоря уж о сравнении энергетического обмена слона и инфузории, когда разница в массе достигает 15 порядков. |
Исходя из этого, Ивлев (1959) в качестве основного показателя энергии жизнедеятельности животных предложил использовать коэффициент а из аллометрических соотношений (46) и (47), который соответствует интенсивности дыхания организма массой в 1 г. Эта величина, конечно, условная, так как слоны, так же как и инфузории, не бывают массой в 1 г, но иного выхода для сравнения стандартного обмена разных видов животных или растений нет.
Попытка Ивлева использовать биоэнергетический показатель для характеристики прогрессивной эволюции животных не удалась (Ивлев, 1963). Более удачливым в этом отношении оказался Дольник (1968). Используя данные о стандартном обмене самых разнообразных групп животных, он показал, что коэффициент а заметно возрастает по мере увеличения сложности организации животных от простейших до млекопитающих и птиц (табл. 1).
Таблица 1.
Диапазон массы тела и стандартный обмен [константы а и k из аллометрического соотношения (46)1 в разных классах и типах животных (Дольник, 1968).
| Типы и классы | Масса, г | A | k | Авторы |
| Protozoa | 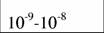
| 0,098 | 0,75 | (1)* |
| Anthozoa | 
| 0, 19 | 0,86 | (2) |
| Oligochaeta | 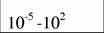
| 0,54 | 0,61 | (3) |
| Xiphosura | 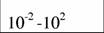
| 0,47 | 0,81 | (2) |
| Crustacea | 
| 0,93 | 0,81 | (4) |
| Mollusca | 
| 1,15 | 0,75 | (2) |
| Insecta | 
| 3,15 | 0,67 | (5) |
| Acrania |
| 2,13 | 0,91 | (6) |
| Cyclostomata | 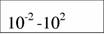
| 1,70 | 0,80 | (7) |
| Pisces | 
| 1,71 | 0,81 | (7) |
| Amphibia | 
| 1,45 | 0,67 | (8) |
| Reptilia | 
| 4,60 | 0,67 | (8) |
| Mammalia | 
| 21,4 | 0,73 | (9) |
| Mammalia |
| 18,0 | 0,76 | (10) |
| Aves | 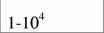
| 51,9 | 0,64 | (И) |
| Aves | 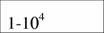
| 21,1 | 0,74 | (12) |
| Aves | 
| 40,9 | 0,66 | (12) |
*(1) Hemmingsen, 1960; (2) Дольник, 1968; (3) Ивлев, 1963; (4) Винберг, 1950; (5) Wigglsworth, 1953; (6) Ивлев, 1961; (7) Винберг, 1956; (8) Brown, 1964; (9) Brody, 1945; (10) Kleiber, 1947; (11) Brody, Proctor, 1932; (12) King, Farner, 1961.
Позднее данные, приводимые Дольником, были уточнены (Коноплев, Зотин, 1975; Коноплев и др., 1975; Зотин, Криволуцкий, 1982; Зотин, 1979, 1984; Zotin, 1990), но основной вывод, сделанный этим автором, что коэффициент а для крупных таксонов порядка класса и типа возрастает по мере усложнения организации животных, полностью подтвердился. Больше того, оказалось, что при сопоставлении стандартного обмена (коэффициента а) той или иной группы животных (табл.1) и времени обнаружения этой группы в палеонтологической летописи (Зотин, 1979; 1984; Зотин, Криволуцкий, 1982), выявляется явная тенденция усиления энергетического обмена по мере протекания биологической эволюции (рис.6). Как видно из этого рисунка, в процессе эволюции происходило последовательное появление животных со все более высоким уровнем стандартного обмена, т.е. увеличивались энергетические возможности организмов. Таким образом, хотя критерий эволюционного прогресса, использованный Ивлевым и Дольником, не получил широкого признания (особенно среди
профессиональных эволюционистов), нет сомнений, что он может быть положен в основу количественной характеристики такого направления прогрессивной эволюции, как биоэнергетический прогресс.
По предположению Дольника (1968) прогрессивная эволюция равнозначна биоэнергетическому прогрессу. На этом положении мы подробнее остановимся во второй части данной книги, здесь же рассмотрим представления двух других авторов, рассматривающих прогрессивную эволюцию с позиций биоэнергетики. Кратко они сводятся к следующему.
И.И.Свентицкий (1977, 1980, 1981) предлагает ввести новый постулат: принцип биоэнергетической целенаправленности, согласно которому
"каждый элемент живой природы в своем развитии и функционировании стремится к состоянию, обеспечивающему в существующих условиях внешней среды наиболее полное использование свободной энергии системой данного трофического уровня, в которую этот элемент входит. " Автор приходит к выводу, "что в процессе эволюции использование свободной энергии каждой нишей непрерывно возрастает как за счет увеличения размеров популяции, использующих данный вид свободной энергии, так и путем повышения их биоэнергетического коэффициента полезного действия" (Свентицкий, 1980, С.7).
К сходным выводам пришел и Н.С.Печуркин (1982). Он формулирует энергетический принцип экстенсивного развития биологических систем в виде двух соотношений

|
(где H е - поток энергии среды, который может быть использован живыми организмами) и энергетический принцип интенсивного развития, согласно которому в процессе эволюции возрастает удельная используемость свободной энергии. "Согласно обсуждавшимся энергетическим принципам, магистральное направление эволюции существует, и оно заключается в увеличении способности захватывать и использовать потоки свободной энергии..." (Печуркин, 1982, С. 106). Реализовать эти принципы живые системы могут при освоении новых ареалов или при резких изменениях условий существования вида за счет быстрого увеличения численности популяции. Подтверждение этим принципам автор находит в экспериментах с микробными популяциями в проточных культурах и в теории ароморфозов (Печуркин, 1982).
Очевидно, что данные, приводимые на рис.6 подтверждают принципы, высказанные указанными выше авторами, как, и идею Шноля (1973, 1979) о возрастании кинетического совершенства в процессе биологической эволюции. Мы же, как будет показано дальше, даем этому явлению иное объяснение, основанное на постулатах термодинамики неравновесных процессов.
Следует также отметить, что принципы Свентицкого и Печуркина призваны объяснить направленность эволюционных процессов и особенно направленность эволюционного прогресса. При этом, хотя в приведенных формулировках они практически не отличаются друг от друга, в методологическом плане они резко различны. Печуркин считает, что эволюция организмов идет не сама по себе, а направляется потоком внешней свободной энергии, создаваемым в конечном счете Солнцем. "Именно поток энергии организует систему (круговорот) и заставляет ее эволюционировать" (Печуркин, 1982, С.46). Наоборот, Свентицкий (1977) включает в свой принцип биоэнергетическую целенаправленность, что подразумевает наличие внутренней причины эволюционного прогресса. Таким
образом, можно проводить аналогию, с одной стороны, между взглядами Печуркина и теорией ортогенеза Эймера (Eimer, 1898; Завадский, 1973), согласно которой направленность эволюционного прогресса зависит от причин, лежащих во внешней среде (климат и пища), с другой - между высказываниями Свентицкого и принципом градаций Ламарка (1937), принципом усовершенствования Нэгели (Nageli,1884), батмизма Копа (Соре, 1896), номогенеза Л.С.Берга (1922, 1977) и другими ламаркистскими теориям, в которых направленность эволюционных процессов и особенно прогрессивной эволюции объясняют целестремительными принципами или законами (Завадский, 1973; Филипченко, 1977; Назаров, 1984).
Между тем, нет необходимости возводить особые эволюционные принципы в виде стремления организмов как можно полнее использовать солнечную энергию или свободную энергию пищи. В этом отношении, как нам кажется, прав Викен (Wicken, 1980), который считает, что использование свободной энергии, идущей от Солнца, осуществляется через селекционизм, а не как прямой ответ на градиент свободной энергии. Градиент солнечной энергии создает как бы особую нишу, заполнение которой чрезвычайно выгодно организмам в борьбе за существование. Действительно, повышение интенсивности энергетического обмена за счет использования свободной энергии, имеющейся в среде, является большим преимуществом организмов, осуществивших это повышение, так как это увеличивает мощность и быстроту реакций организмов, возможность освоения новых географических районов и экологических ниш, увеличивает устойчивость и надежность физиологических процессов, протекающих в организмах или осуществляемых организмами. И здесь не требуется введение особых принципов, как это предложили Свентицкий и Печуркин - естественный отбор сам направляет эволюцию, когда это возможно, по пути все большего использования свободной энергии для повышения уровня энергетического обмена и создает направленность
эволюционного процесса, который А.Н.Северцов называл морфо-физиологическим прогрессом.
Это не означает того, что механизмы такого макроэволюционного процесса, как морфо-физиологический прогресс, полностью определяются механизмами микроэволюции. Арогенез, в соответствии с определением ароморфозов, из которых складывается арогенез, состоит в увеличении общей энергии жизнедеятельности организмов и, следовательно, его осуществление возможно только путем изменения интенсивности энергетического обмена. Последнее происходит в организмах только при определенных условиях, о которых будет подробно сказано дальше. Стоит только отметить, что в конечном счете механизмы эволюционных изменений интенсивности энергетического обмена организмов не осуществимы без участия микроэволюционных процессов. А это означает, что понимание механизмов прогрессивной эволюции базируется на теории естественного отбора.
И все же не следует забывать, что согласно термодинамике неравновесных процессов при эволюции сложных систем к равновесию действует принцип наискорейшего спуска, который предполагает появление в системе подсистем, двигающихся в сторону увеличения неравновесности, что и происходит в процессе прогрессивной эволюции в отдельных группах животных и растений (Зотин, Зотин, 1995).