Термодинамика и направленность эволюции
По вопросу о направленности эволюции существует не только большая литература, но и длительная, почти 200-летняя, дискуссия между ламаркистами и дарвинистами, конца которой не видно (Назаров, 1984; Северцов, 1990). Краткая суть этой дискуссии состоит в том, лежат ли в основе биологической эволюции особые законы (эволюция на основе закономерностей - номогенез) или для понимания всех ее особенностей достаточно теории естественного отбора (эволюция на основе случайностей - селекционизм). Выше говорилось, что термодинамика, как чисто дедуктивная наука, решает наиболее общие вопросы, связанные с изучением направления и движущих сил любого реального процесса. Это позволяет надеяться, что использование ее понятий и методологии создаст новые возможности в понимании общих закономерностей биологической эволюции и эволюции биосферы. Подобную возможность осознают не только физики и механики (Prigogine et al., 1972; Николис, Пригожин, 1977; Эбелинг, 1979; Пригожин, 1983; Моисеев, 1987; Хазен, 1993а.1996), но и биологи (Wicken,
1979-1987; Wiley, Brooks, 1983; Brooks, 1984; Brooks, Wiley, 1984, 1988; Swensen, 1989, 1992, 1997).
Первая проблема, которая встает при попытке использования термодинамики при построении закономерностей биологической эволюции состоит в том, как применить термодинамические критерии (12), (13), (18), показывающие, что в процессе эволюции физической системы к равновесию или стационарному состоянию происходит непрерывное уменьшение диссипативных функций, с увеличением сложности строения и организованности живых организмов в процессе прогрессивной эволюции? Эта проблема, несмотря на ее кажущуюся трудность, решается довольно просто: в большой системе, такой как биосфера, всегда возможно существование подсистем, которые в процессе изменений не приближаются, а удаляются от равновесного состояния за счет сопряженных процессов, происходящих в основной системе [см.(10)]. К этому выводу, естественно, пришли многие авторы, рассматривающие возможность использования термодинамики для понимания проблем эволюции (Wicken, 1979; Гладышев, 1988, 1996; Голубев, 1991).
К сожалению, многие из них опираются при этом на методологию классической термодинамики, хотя и используют иногда такие понятия, как поток свободной энергии, диссипация энергии. Например, Голубев (1990, 1991) за основу критериев прогрессивной эволюции биосистем принимает увеличение удельной свободной энергии. Нет, однако, доказательств, что удельная свободная энергия более продвинутых по пути прогрессивной эволюции организмов выше, чем у организмов, стоящих на более низкой ступени организации. Более вероятно, что она одинакова, например, в классе млекопитающих у приматов и однопроходных или в типе моллюсков у головоногих и панцирных. Все дело в том, как отмечалось выше, что открытые системы, далекие от равновесия, какими являются все без исключения живые организмы, не следует характеризовать такими термодинамическими величинами, как свободная энергия, энтропия, энтальпия и др., но только скоростями изменения этих величин: продукцией и потоком
энтропии (5), диссипативными функциями (8), (9), (14), (15), критериями эволюции (12), (13), (17), (18), (52), (53).
Сказанное относится не только к работам Голубева (1990, 1991) или Гладышева (1977, 1988, 1996), но и ко многим другим современным исследованиям, в которых разрабатываются проблемы эволюции с позиции термодинамики (Wicken, 1979-1987; Wiley, Brooks, 1982; Brooks, 1984, 1986; Brooks, Wiley, 1984, 1988; Swensen, 1989, 1992, 1997; Хазен, 1993а,б, 1996). Хотя последние хорошо осознают необходимость использования для понимания проблем эволюционного учения не классической термодинамики, а термодинамики неравновесных процессов, в решении многих проблем, связанных с рассмотрением вопросов увеличения сложности и организованности животных и растений в процессе прогрессивной эволюции, они остаются на позициях термодинамики равновесных процессов, особенно когда пользуются понятиями теории информации.
И все же в этих работах высказано много идей, которые полезны для понимания прогрессивной эволюции и проблемы направленности эволюционного процесса. Прежде всего это касается, как отмечалось, сопряженных процессов, за счет которых может осуществляться прогрессивная эволюция в биосфере. Викен (Wicken, 1980), например, речь ведет о градиенте свободной энергии, создаваемой Солнцем, в пределах которого эволюция биосферы осуществляется в направлении стационарного состояния с минимальной диссипацией энергии и максимальной структурированностью.
В то же время при обсуждении проблем возрастания сложности строения организмов в процессе прогрессивной эволюции и увеличения молекулярной информации в последовательностях ДНК за счет естественного отбора Викен (Wicken, 1980, 1987), так же как Брукс и др. (Brooks et al., 1984; Brooks, Wiley, 1988) опирается на понятия информации и негэнтропии. Позднее Хазен (1993а,б, 1996), не используя понятие негэнтропии, пришел к сходным представлениям. К сожалению и в подобного рода исследованиях авторы остаются на позициях
классической термодинамики, которая способна описать только конечные состояния системы, так как в классических построениях процессы протекают бесконечно медленно и понятия скоростей, столь характерные при неравновесных процессах, протекающих в открытых системах, не входят в методологию равновесной термодинамики.
Вторая проблема касается кинетики перехода биосферы к конечному стационарному состоянию и сил, вызывающих этот процесс. Классическая термодинамика (включая и второе начало) ничего не говорит о подобного рода процессах, так как она может судить только о конечных состояниях системы. Это можно сделать, только опираясь на термодинамику неравновесных процессов, для которой скорости процессов и время не являются чужеродными понятиями. Николис и Пригожин (1979), в частности, приходят к выводу, что в сильно неравновесной системе, поддерживаемой непрерывным потоком энергии, как это имеет место в системе Солнце-Земля, могут возникнуть структурные флуктуации, которые по принципу положительной обратной связи способны ускорить возникновение новых структурных флуктуации согласно схеме:
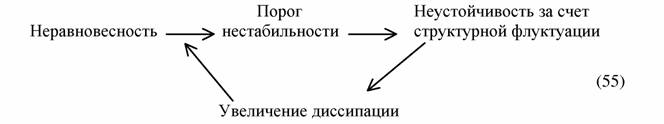
|
| Авторы приходят к выводу, "... что в качестве движущей силы эволюции следует рассматривать энергетическую диссипацию. Здесь уместно отметить, что, несмотря на повышение уровня организации и сложности живых систем, с течением времени происходит ускорение биологической эволюции. Этот процесс можно представить так, что каждая новая ступень функциональной организации |
несет в себе зародыш дальнейшей эволюции" (Николис, Пригожин, 1979, с.456). На наш взгляд (Зотин, Зотин, 1995) схема (55) показывает возможный механизм прогрессивной эволюции, но не ее движущие силы. Последние связаны с принципом наискорейшего спуска термодинамики неравновесных процессов и существованием градиента солнечной энергии.
В системе Солнце-Земля, где градиент подводимой к Земле энергии приблизительно постоянен на протяжении миллиардов лет, возникает гигантский переходной процесс в виде возникновения жизни и образования биосферы. Он заканчивается, когда гидросфера и литосфера полностью заполняются жизнью и она достигает максимального совершенства в отношении улавливания солнечной энергии, т.е. когда эффективность фотосинтеза растений близка современному значению. К этому моменту биосфера достигает стационарного уровня и медленно (так как величина попадающей на Землю солнечной энергии практически не меняется) начинает эволюционировать в сторону равновесия. На всех этапах эволюции биосферы действует механизм естественного отбора, но для прогрессивной эволюции материал ему поставляют возникающие по схеме (55) живые системы с более сложной и совершенной структурой. Движущей силой на всех этапах эволюции биосферы является градиент солнечной энергии и термодинамические силы, определяемые принципом наискорейшего спуска.
В первой главе было показано, что в термодинамике неравновесных процессов разработан принцип, который может быть равнозначен началу термодинамики и который описывает макрокинетику необратимых процессов - принцип наискорейшего спуска. Если исходить из этого принципа, то оказывается, что неравновесная термодинамическая система (в данном случае биосфера) тем быстрее движется к состоянию равновесия, чем больше в ней образуется подсистем с возрастающей неравновесностью. Другими словами, приближение общей термодинамической системы к состоянию равновесия сопровождается появлением подсистем, двигающихся в противоположном направлении, т.е. по пути прогрессивной эволюции (Зотин, Зотин, 1995; Zotin, Zotin, 1996). Более того,
появление таких подсистем является обязательным следствием принципа наискорейшего спуска.
Сходные идеи раньше нас развил Свенсен (Swensen, 1992) и Хазен (1993а). Свенсен еще в 1982 г., опираясь на закон максимальной продукции энтропии, показал, что при возникновении порядка энтропия продуцируется быстрее, чем при возникновении беспорядка (Swensen, 1992). Если исходить из принципа наискорейшего спуска, то это означает, что возникновение порядка в процессе эволюции является обязательным условием его осуществления.
Хазен (1993 а) исходил из статистической интерпретации понятия энтропии и информационног смысла этого понятия. Даже название его статьи, «Принцип максимума производства энтропии и движущая сила прогрессивной биологической эволюции», опубликованной в журнале «Биофизика» за 1993 год, показывает, насколько близки наши взгляды. Однако мы шли к использованию понятий неравновесной термодинамики, к пониманию движущей силы прогрессивной эволюции разными путями. В то время как Хазен использовал информационное толкование понятия энтропии и изменение нормировки энтропии, мы опирались на чисто феноменологический принцип - принцип наискорейшего спуска термодинамики неравновесных процессов (Зотин, Зотин, 1995). По нашему мнению этот путь более прост и, главное, позволяет проверить сделанные выводы на многочисленных экспериментальных материалах, полученных при изучении энергетического обмена разных видов животных. Теория Хазена, основанная на понятии энтропии в статистической и информационной интерпретации, относится к более сложной и мало разработанной области статистической термодинамики и теории информации, и не совсем ясно можно ли и как можно ее использовать в термодинамике биологических процессов. Примеры, приводимые автором, носят качественный характер и не позволяют количественно характеризовать прогрессивную эволюцию животных. Вместе с тем путь, намеченный Хазеном (1991, 1992, 1993 а), кажется перспективным, особенно для понимания возникновения и использования
генетической информации, работы мозга и др. Несмотря на это в дальнейшем при рассмотрении проблем прогрессивной эволюции мы будем опираться на чисто феноменологический подход, так как он имеет более зримую биологическую основу.
Следует остановиться еще на одном вопросе. При возникновении жизни (предбиологическая эволюция по Эйгену), при распространении жизни по поверхности Земли и в процессе прогрессивной эволюции решающее значение имеет естественный отбор. Из теории естественного отбора вытекают различные принципы оптимальности (Розен, 1969; Ханин и др., 1978; Озернюк, 1992), которые по внешней форме и по критериям иногда напоминают термодинамические принципы и критерии эволюции. Например, принцип минимума энергии в онтогенезе, предложенный Озернюком (Озернюк, 1988, 1989, 1992), вытекает из естественного отбора, так как минимальные траты на развитие явно выгодны для организмов. В то же время этот принцип вытекает также из чисто термодинамических принципов: интегрального принципа наименьшей диссипации энергии и принципа наискорейшего спуска. Наоборот, попытку использовать принцип наименьшей диссипации энергии для понимания проблем эволюции (Моисеев, 1987) можно легко свести к принципу оптимальности, т.е. к довольно тривиальному утверждению, что естественный отбор действует в направлении наиболее экономного расходования энергии организмами. Вообще, в тех случаях, когда принцип оптимальности, вытекающий из теории естественного отбора, относится к энергетическим процессам он, как правило, соприкасается с термодинамическим принципом минимальной диссипации энергии. Естественно, эти принципы не противоречат один другому.
Но есть области теории эволюции, где термодинамические подходы не стыкуются с принципами оптимальности или играют более существенную роль для понимания эволюционных проблем. Это прежде всего критерии эволюции (18), (52) и (53), которые указывают направление изменений биологических процессов при индивидуальном и историческом развитии организмов и биосферы
в целом. Это касается и принципа наискорейшего спуска - основного двигателя прогрессивной эволюции. Естественный отбор (но не принципы оптимальности), как и процессы типа (55), играют, конечно, роль в механизме осуществления прогрессивной эволюции, но не они являются главным двигателем происходящих изменений.