Критерий упорядоченности
Как уже говорилось, согласно предположению Ивлева (1959, 1963) и Дольника (1968, 1978) арогенез состоит в усилении энергетического обмена организмов: усилении основного или стандартного обмена. Это очень важное заключение для проблем прогрессивной эволюции. Рассмотрим, поэтому, подробнее термодинамическую основу такого критерия биоэнергетического прогресса как стандартный обмен.
С термодинамической точки зрения состояние равновесия соответствует полному беспорядку - наиболее вероятному состоянию системы. Если система уклоняется от состояния равновесия, вероятность ее состояния уменьшается и степень упорядоченности возрастает. Отсюда следует, что упорядоченность - понятие, обратное беспорядку. Следовательно, упорядоченность в устойчивом состоянии любой термодинамической системы будет тем выше, чем дальше система отстоит от состояния равновесия.
В классической термодинамике мерой беспорядка является энтропия и ее возрастание в ходе движения изолированной системы к состоянию равновесия
сопровождается увеличением неупорядоченности системы.
Обратный процесс - уменьшение энтропии - соответствует увеличению упорядоченности системы. В термодинамике неравновесных процессов принимается, что движение открытой системы к равновесию характеризуется уменьшением диссипативной функции (12), а точнее функции внешней диссипации (13), (18). Следовательно, в открытых неравновесных системах возрастание неупорядоченности связано не с увеличением энтропии, а с уменьшением псидфункции. Соответственно, упорядоченность открытой системы в устойчивом состоянии тем больше, чем больше величина функции внешней диссипации. Эта функция обратно пропорциональна вероятности состояния системы (29), что еще раз подчеркивает ее связь с упорядоченностью. Таким образом, величина функции внешней диссипации в устойчивом состоянии открытой системы может служить мерой ее упорядоченности. Она показывает степень удаленности системы от равновесного состояния.
Исходя из сказанного, степень упорядоченности живых систем можно сопоставлять с величиной стандартного обмена. Действительно, диссипативная функция организмов равна интенсивности потребления кислорода (64) или, учитывая (65), интенсивности теплопродукции, а величина функции внешней диссипации или псидфункции организма, находящегося в состоянии покоя, показывает степень их удаленности от состояния равновесия, т.е. степень их упорядоченности.
Величина стандартного обмена связана с массой тела организмов аллометрической зависимостью (46), (47). Как говорилось, из этих зависимостей следует, что сравнивать стандартный обмен видов животных, имеющих разные размеры, затруднительно, так как масса организмов может отличаться на многие порядки. Поэтому, как отмечалось, для целей сравнения используют коэффициент а из аллометрической зависимости (46) или (47). Этот коэффициент может быть положен в основу количественной характеристики направления прогрессивной эволюции (Коноплев, Зотин, 1975; Коноплев и др., 1975; Зотин, Криволуцкий, 1982; Zotin, 1990). Чтобы показать это более наглядно, рассмотрим понятие критерия упорядоченности.
В термодинамике неравновесных процессов устойчивое состояние системы характеризуется величиной удельной диссипативной функции (8) или
величиной функции внешней диссипации (14). Удельная
диссипативная функция связана обратной зависимостью с плотностью вероятности состояния (24), т.е., как говорилось, может быть использована в качестве показателя степени упорядоченности системы. В еще большей степени это относится к функции внешней диссипации, которая просто обратно пропорциональна плотности вероятности состояния системы (29).
Псидфункция равна интенсивности теплопродукции (14) и, поэтому,
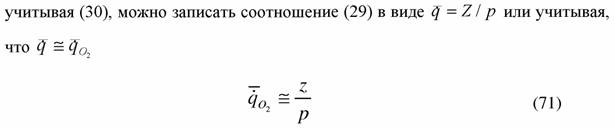
|
| Для того, чтобы можно было использовать формулу (71) в экспериментальной работе необходимо ввести понятие отношения вероятностей (Коноплев, Зотин, 1975; Коноплев и др., 1975). Пусть значение величин с индексом p относится к простейшим организмам, например, к амебам, а без индекса - к любым другим организмам. Используя (30) и (47) и учитывая, что для всех организмов b=0.25, напишем для отношения вероятностей живых организмов (ρο ): |
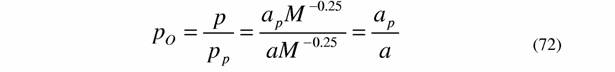
|
| Расчет по данным, приводимым Хеммингсеном (Hemmingsen, 1960) для простейших и микроорганизмов дает величину aр = 0,098 мВт/г (Зотин, Криволуцкий, 1982). Использование более многочисленных наблюдений о стандартном обмене простейших показало (см. табл. 6), что в качестве эталона из группы простейших лучше использовать тип Sarcomastigophora, для которого ар = 0,082 мВт/г (Владимирова, Зотин, 1985). Уточнение методики расчета коэффициента к в формуле (46) показало, что для отряда Amoebida, который мы выбираем в качестве эталона, как наиболее просто устроенных эукариот, ap =0.067 мВт/г (табл.6). Исходя из этого, для условного организма с массой в 1 |
г, согласно формуле (72) выражение для отношения вероятностей состояния любого организма равно:
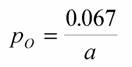 (73)
(73)
Таким образом, предложение Ивлева (1959, 1963) и Дольника (1968, 1978) использовать в качестве критерия эволюционного прогресса коэффициент а из аллометрических соотношений (46), (47) получает более надежную термодинамическую основу.
Для большей наглядности мы предложили использовать в сравнительных исследованиях величину обратно пропорциональную относительной вероятности состояния организма и назвали ее критерием упорядоченности (Коноплев, Зотин, 1975; Коноплев и др., 1975). Согласно (73), он равен
 (74)
(74)
В отличие от коэффициента а из формул (46), (47) коэффициент а в формуле для критерия (74) зависит от некоторых особенностей физиологии животных и от экологических условий их жизни. Чтобы иметь это в виду, мы обозначили его через а'. Когда физиологическими и экологическими особенностями можно пренебречь, то а'= а.
Коэффициент а в критерии упорядоченности Сr (74), так же как коэффициент а, зависит от температуры, при которой производится измерение дыхания животных. Мы приняли, что стандартный обмен пойкилотермных животных должен приводиться к температуре 20°С, а гомойотермных животных - к критической точке термонейтральной зоны. Это одно из условий определения указанных коэффициентов. При определении критерия упорядоченности гомойотермных животных следует учитывать также температуру тела животных. У млекопитающих она достигает 36-38 °С, а у птиц и некоторых насекомых - 40-42°С.
Критерий упорядоченности (74), как говорилось, характеризует не столько стандартный обмен данного животного, а степень его удаленности от состояния равновесия. Поэтому его величина не должна зависеть от факторов внешней среды и таких особенностей гомойотермных животных, как терморегуляция. Отсюда следует, что для определении критерия упорядоченности гомойотермных животных, сопоставимого с критерием упорядоченности пойкилотермных, необходимо приводить коэффициент а' в формуле (74) по кривой Крога к температуре 20°С. Здесь мы снова прибегаем к довольно искусственному приему, так как температура тела равная 20°С в норме не встречается у гомойотермных животных. Но при определении критерия упорядоченности гомойотермных животных такой прием обязателен.
Вторая проблема, которую следует учитывать при вычислении критерия упорядоченности как пойкилотермных, так и гомойотермных животных состоит в следующем. При сопоставлении стандартного обмена животных с помощью аллометрических соотношений (46) и (47) предполагается, что в дыхании участвует вся масса тела животного. Но хорошо известно, что это не так: многие животные имеют специализированные опорные и защитные образования, которые не принимают или принимают незначительное участие в процессах дыхания. У многих водных пелагических беспозвоночных животных тело настолько оводнено, что трудно сопоставлять общую массу таких животных с массой донных животных и, тем более, наземных. Встает вопрос: как в этом случае определять сопоставимый стандартный обмен (коэффициент а') у разных видов животных?
Мы не будем подробно обсуждать этот вопрос, хотя ему посвящена довольна обширная литература, а примем, что его можно определять по содержанию воды в теле животного. Содержание воды колеблется в значительных пределах, поэтому приходится выбрать некий эталон, к которому приводить массу тела всех остальных животных. Для наземных животных за такой эталон мы принимаем содержание воды равное 70%, к которому близко содержание воды в теле многих животных.
Например, в табл.3 приведены данные о содержании воды в теле некоторых млекопитающих, определенные прямым методом и методом разведения воды, меченной стабильными или радиоактивными изотопами
водорода (Culebras et al., 1977). Судя по данным, приводимым в этой таблице содержание воды в теле млекопитающих в среднем равно 68.7±0.9%. Это достоверно не отличается от эталонного содержания воды в 70%, тем более, что по измерению Калебраса и др. (Culebras et al., 1977) у крыс весом 227 г (п=21) содержание воды в теле, определенное прямым методом равнялось 70.2%, а методом разбавления тритиевой воды - 71.4%. Близкое к 70% содержание воды в теле получено и для других сухопутных животных, данные о дыхании которых приведены в этой книге. Оно равно в среднем 69.7+0.6 (п=64).
Таблица 3.
Содержание воды в теле млекопитающих, определенное разными авторами прямым методом и методом разведения меченой воды (Culebras et al., 1977).
| Вид | Содержание воды в теле (%) | |
| Прямой метод | Метод разбавления | |
| Крысы | 65.3 | - |
| 65-70 | - | |
| 57.6 - 68.7 | - | |
| 63.9 | 68.1 | |
| 66.5 | 70.8 | |
| 66.4 | 74.4 | |
| 70.2 | 71.38 | |
| Кролик | 64.2 | - |
| 68.8 | - | |
| 73.6 | 72.8 | |
| - | 65.5 | |
| Морская свинка | - | 75.2 |
| Собака | 65,7 | - |
| Критерий упорядоченности (74) опирается на коэффициент а' . Из формулы (46) следует, что коэффициент а равен |
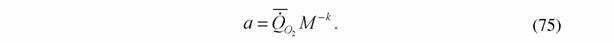
|
В тех случаях, когда степень оводненности меньше эталонной величины, что связано с наличием скелетных образований, расчет коэффициента а' согласно формуле (75) и данным о содержании воды в теле сухопутных животных следует проводить по формуле:
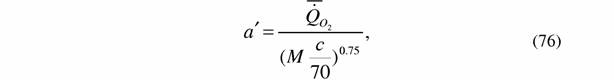
|
| или |

|
| где с - процентное содержание воды в теле животного. Что касается водных животных, то у них содержание воды в теле иное, чем у наземных. Мы приняли в качестве эталона для водных животных содержание воды в теле простейших, так как в этом случае определение содержания воды в большинстве случаев не осложнено скелетными образованьями. У простейших содержание воды в теле в среднем равно 86.5% (Владимирова, Зотин, 1983). Эту величину мы и примем за эталон для водных животных. Когда содержание воды в теле водных животных ниже этой величины следует использовать формулы аналогичные (76) и (77): |

|
| или |
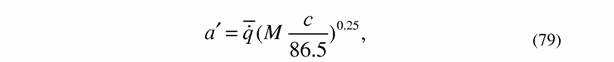
|
| так как разница скорее всего связана с наличием скелетных образований в теле животных. |
| Когда содержание воды в теле водного животного больше эталонного, что как правило связано с пелагическим образом жизни (например, у медуз или |
гребневиков), мы также должны увеличивать реальные данные о стандартном обмене на разность между эталонным и измеренным в опыте. Но в этом случае нужно использовать формулы:

|
| или |

|
Наконец еще одна проблема, которую, как считают многие специалисты, необходимо учитывать при сопоставлении стандартного обмена разных видов животных: экологические (точнее сказать, климатические) условия их жизни. Существует такое понятие, как температурная компенсация, которая означает, что у животных, живущих в более холодном климатическом поясе стандартный обмен выше, чем у животных из более теплых поясов Земли (Schölander et al., 1953; Ивлева, 1972, 1981; Слоним.1982а,в; Зотин, 1988; Озернюк, 1992). Долгое время наличие температурной компенсации у животных вызывало недоверие, пока Шолендер и др. (Schölander et al., 1953) точно не показали ее у ракообразных (рис. 11).
Наличие температурной компенсации показано не только для далеких видов, но и для близких видов и даже подвидов, например, змей и ящериц (Hailey, Davies, 1986; Patterson, Davies, 1989).
При определении сопоставимого стандартного обмена животных температурную компенсацию, как и влияние иных условий жизни (горы, пустыни и т.д.) можно не учитывать, так как величина коэффициента а закреплена генетически. Но в случае критерия упорядоченности учет этих обстоятельства желателен, так как в этом случае мы определяем степень удаленности данного животного или группы животных от состояния равновесия вне зависимости от того в каких условиях они живут. Исходя из этого, при расчетах коэффициента упорядоченности приходится получать данные о различиях коэффициента а' у представителей животных, живущих в разных
климатических поясах. В настоящее время мы можем это сделать достаточно грубо, относя одних животных к умеренному поясу, других - к тропическому.
При рассмотрении данных о стандартном обмене разных видов животных, живущих в разных климатических поясах, следует учитывать не только экологические условия жизни, но и видовую принадлежность. Чтобы уменьшить ошибку при определении температурной компенсации следует, поэтому брать средние данные о стандартном обмене данного вида, а не все имеющиеся наблюдения, которые для одних видов очень многочисленны, а для других - единичны. В табл. 4 приведены эти данные, взятые из статьи Зотина и Алексеевой (1997) о насекомых и рассчитанные по материалам статьи Зотина и Владимировой (1986а,б) о млекопитающих. Как видно из этой таблицы, имеются существенные различия в соотношении стандартного обмена умеренного и тропического поясов (ау/ат) у представителей разных отрядов насекомых и млекопитающих, но не исключено, что эти различия связаны с методическими неточностями при их определении. Поэтому было бы разумным принять, что для всех животных он одинаков. В среднем, судя по данным, приводимым в табл.4, ау/ат=1.66. Мы примем эту величину в качестве единого коэффициента при сопоставлении стандартного обмена животных, живущих в теплых странах с животными умеренного и арктического пояса. Грубость этого приема очевидна. Мы не учитываем также других условий жизни, с которыми, по-видимому, придется считаться при более точном определении критерия упорядоченности, чем мы это сделали в данной книге.
Таблица4
Сопоставимый стандартный обмен [коэффициент а (мВт/г) из аллометрического соотношения (46)] и соотношение стандартного обмена умеренного и тропического поясов αу/αT у разных животных. N - число изученных видов.
| Отряд | Климатические пояса | а | αу/αT | N |
| Насекомые | ||||
| Blattodea | Тропический | 0.83±0.15 | ||
| Умеренный | 1.05±0.19 | 1.28 |
| Odonata | Тропический | 2.14±0.13 | ||
| Умеренный | 2.47 | 1.15 | ||
| Orthoptera | Тропический | 1.10±0.12 | ||
| Умеренный | 2.49±0.13 | 2.26 | ||
| Lepidoptera | Тропический | 1.92±0.32 | ||
| Умеренный | 2.84±0.25 | 1.48 | ||
| Coleoptera | Тропический | 1.19±0.10 | ||
| Умеренный | 3.33±0.65 | 2.80 | ||
| Diptera | Тропический | 1.99±0.57 | ||
| Умеренный | 3.76±0.61 | 1.89 | ||
| Hymenoptera | Тропический | |||
| Умеренный | 1.13 | |||
| Млекопитающие | ||||
| Insectivora | ||||
| Сем. Soricidae | Тропический | 33.67±3.28 | ||
| Умеренный | 55.30±5.17 | 1.64 | ||
| Chiroptera | ||||
| Сем. Vespertiliondae | Тропический | 12.73±1.22 | ||
| Умеренный | 16.59±1.45 | 1.30 | ||
| Carnivora | ||||
| Сем. Canidae | Тропический | 15.85±2.37 | ||
| Умеренный | 21.32±0.74 | 1.35 | ||
| Rodentia | ||||
| Сем. Cricetidae | Тропический | 20.24±0.86 | ||
| Умеренный | 31.85±1.21 | 1.57 | ||
| Сем. Muridae | Тропический | 17.71±1.50 | ||
| Умеренный | 37.07±5.56 | 2.09 |
Из сказанного ясно, что относительная упорядоченность животных, степень их уклонения от термодинамического равновесия можно измерить с помощью критерия упорядоченности (74), используя величину коэффициента а', т.е. несколько видоизмененный коэффициент а из аллометрических соотношений (46) и (47). Таким образом, предложение Ивлева (1959, 1963) и
Дольника (1968, 1978) использовано и нами в данной работе. Следует только учитывать, что коэффициент а характеризует биоэнергетический прогресс организмов, а критерий упорядоченности собственно прогрессивную эволюцию - удаленности организмов от состояния равновесия.
Итак, в настоящее время в качестве критерия прогрессивной эволюции, по нашему мнению, следует использовать критерий упорядоченности (74). Очевидно, что для количественной характеристики, как биоэнергетического прогресса, так и собственно прогрессивной эволюции требуется получение возможно более точных и подробных данных о стандартном обмене представителей различных таксонов от простейших до млекопитающих и птиц.