Константа Рубнера
Еще один критерий эволюционного прогресса, который может быть получен в цифровом выражении, предложен Э.С.Бауэром (Bauer, 1931; Бауэр, 1935) и назван им константой Рубнера. Этот критерий основан на следующем.
В процессе развития, роста и старения организмов происходит снижение стандартного обмена, прерываемое на некоторых стадиях эмбрионального и раннего постэмбрионального развития различными уклонениями от этого основного процесса (Зотин, 1974; Зотин, Зотина, 1976, 1993). Согласно Коловей (Galloway, 1974) у всех гомойотермных животных независимо от вида, предельная величина снижения интенсивности потребления кислорода к моменту смерти, наступающей в результате старения, равна 0.97 мВт/г. Хотя эта величина, по-видимому, завышена, она показывает, что продолжительность жизни имеет естественный предел. Следовательно, за весь онтогенез организм может использовать для осуществления процессов развития, роста и старения строго определенное количество энергии на грамм массы тела. Отсюда можно принять, что средняя интенсивность потребления кислорода взрослым организмом умноженная на продолжительность его жизни является величиной постоянной для данного вида, т.е.
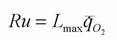 (83)
(83)
где Zmax - максимальная продолжительность жизни животного (годы);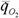 средняя интенсивность потребления кислорода (мВт/г); Ru - константа, которую Бауэр (1935) и предложил назвать константой Рубнера, так как именно этот ученый впервые показал, что потребление энергии на протяжении жизни млекопитающих разных видов приблизительно одинаково (Rubner, 1908a,b).
средняя интенсивность потребления кислорода (мВт/г); Ru - константа, которую Бауэр (1935) и предложил назвать константой Рубнера, так как именно этот ученый впервые показал, что потребление энергии на протяжении жизни млекопитающих разных видов приблизительно одинаково (Rubner, 1908a,b).
Бауэр (1935) понимал, что для определения количества энергии, потребляемой организмом на протяжении всей жизни, следует использовать не сумму средних величин интенсивности дыхания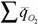 , а интеграл вида
, а интеграл вида Сравнительно недавно Райс (Reiss, 1989) пришел к сходной идее, предлагая характеризовать продолжительность жизни животных от момента
Сравнительно недавно Райс (Reiss, 1989) пришел к сходной идее, предлагая характеризовать продолжительность жизни животных от момента
оплодотворения до смерти с помощью понятия удельного метаболизма всей жизни, которое может быть выражено формулой:
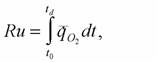 (84)
(84)
где t0, - время оплодотворения; td - время смерти. Константа Рубнера в этом случае характеризует не только продолжительность жизни, но и скорость старения организма и в этом отношении имеет заметное преимущество перед определением (83). Однако для вычисления ее по формуле (84) необходимо иметь данные об изменении интенсивности дыхания животного в разные периоды онтогенеза. Таких данных в литературе немного, поэтому, при вычислении константы Рубнера для подавляющего числа видов животных приходится пользоваться формулой (83).
Аллометрическое соотношение для константы Рубнера можно получить и чисто теоретическим путем. Было обнаружено (Sacher, 1959, 1978; Lindstedt, Calder, 1976; Calder, 1976, 1984; Blueweiss et al., 1978; Prinzinger, 1979, 1989; Economes, 1980; Шмидт-Ниельсон, 1987; Гаврилов, 1991), что максимальная продолжительность жизни животных связана с массой тела аллометрическим соотношением
 (85)
(85)
где Lmax - максимальная продолжительность жизни (годы); M - масса тела (г); lт и k1 - коэффициенты. Нас будет интересовать в данном, случае коэффициент k1. Показано, что для млекопитающих k1=0,12 и для птиц k1=0,16 (Зотин, 1979). В тоже время мы приняли, как и многие другие авторы, что коэффициент b из аллометрического соотношения для зависимости интенсивности дыхания от массы тела (47) равен для всех животных -0.25 (66). Отсюда следует, что аллометрическое соотношение для константы Рубнера равно
 (86)
(86)
Очевидно, что константа Рубнера не может быть одинаковой у всех групп животных, так как они сильно отличаются по уровню стандартного обмена и тем более по продолжительности жизни.
Бауэр (Bauer, 1931; Бауэр, 1935) учел это обстоятельство и пришел к выводу, что в ходе прогрессивной эволюции происходит изменение константы Рубнера. Он предложил использовать константу Рубнера для характеристики прогрессивной эволюции. И действительно по его расчетам (табл.5) оказалось, что эта константа возрастает по мере усложнения строения животных.
Таблица5 Максимальная продолжительность жизни Lmax (годы), масса тела M (г), скорость потребления кислорода 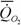 (ккал/день) и константа Рубнера Ru (кДж/г) у различных типов и классов животных (Бауэр, 1935).
(ккал/день) и константа Рубнера Ru (кДж/г) у различных типов и классов животных (Бауэр, 1935).
| Вид | Lmax | M | 
| Ru |
| Colenterata | ||||
| Carmanina hastata | 0,2 | 0,007 | 0.193 | |
| Rhizostoma pulmo | 0,2 | 0,083 | 0.416 | |
| Echinodermata | ||||
| Cucumaria sp. | 5,0 | 0,022 | 19,4 | |
| Vermes | ||||
| Hirudo sp. | 20,0 | 0,008 | 28,2 | |
| Lumbricus sp. | 10,0 | 0,023 | 131,9 | |
| Mollusca | ||||
| Limnaeus sp. | 4,0 | 2,3 | 0,022 | 67,3 |
| Ostrea virginica | 10,0 | 50,0 | 0,086 | 30,3 |
| Arthropoda | ||||
| Astacus sp. | 25,0 | 31,0 | 0,137 | 194,4 |
| Palinurus sp. | 25,0 | 1,27 | 223,5 | |
| Homarus sp. | 25,0 | 2,47 | 350,5 | |
| Vertebrata | ||||
| Equus cballus |
| Ovis ammon | ||||
| Canis familiaris | ||||
| Felis silvestris | 9,5 | |||
| Homo sapiens |
Так как в состав константы Рубнера (83) входит интенсивность потребления кислорода ( ), то ее увеличение характеризует возрастание
), то ее увеличение характеризует возрастание
упорядоченности организмов. Но, с другой стороны, в ее состав входит максимальная продолжительность жизни, которая зависит от степени организованности животных, от экологических условий существования вида и др. Таким образом, константа Рубнера, как критерий прогрессивной эволюции, страдает некоторой неопределенностью.
Из сказанного следует, что требуются дополнительные, уточненные данные о величине максимальной продолжительности жизни животных, исследование зависимости этой величины от условий жизни, от величины мозга и др. прежде чем использовать константу Рубнера для уточнения "шкалы прогресса" в животном царстве.