Низшие беспозвоночные
Для большинства типов и классов беспозвоночных показана аллометрическа зависимость (46). Однако для многих типов низших беспозвоночных из-за ограниченного числа измеренных видов животных точность определения коэффициентов в подобного рода зависимости невелика. Для нас особенно важным является точное определение коэффициента k, поэтому независимо от того, какое значение этого коэффициента получено в различных работах, мы принимаем его равным 0.75.
Тип Porifera(Губки)
В отношении представителей типа Porifera в литературе имеются лишь единичные наблюдения о потребления кислорода взрослыми особями. Этих данных слишком мало, чтобы рассчитать величину коэффициента а и критерий упорядоченности Сr. Кроме того, губки большей частью образуют обширные колонии и бывает трудно определить о какой массе организма должна идти речь, когда мы определяем коэффициент а и критерий упорядоченности Сr. Поэтому мы не будем приводить данных о стандартном обмене губок, хотя нам известно несколько работ, в которых измерялась интенсивность дыхания некоторых видов губок (Gordon et al., 1955; Ивлев,
Яковлева, 1963; Виленкина, Виленкин, 1969; Cotter, 1978).
ТипCtenophora(Гребневики)
Не очень многочисленные данные о потреблении кислорода взрослыми формами гребневиков, полученные при разных температурах, мы приводили к температуре 20°С по формуле (69).
Зависимость дыхания от массы тела у представителей типа гребневиков показана на рис. 14(1). Как видно из этого рисунка характер кривой для типа гребневиков заметно отличается от кривой для типа кишечнополостных. Расчет по данным, приводимым на этом рисунке, показал, что аллометрическая зависимость скорости потребления кислорода от массы тела у гребневиков имеет вид:

Однако число измерений и диапазон масс измеренных гребневиков не велик, поэтому мы будем считать, что и в этом случае k=0.75.
С данными об интенсивности потребления кислорода представителей этого типа животных дело обстоит примерно так же, как и с представителями некоторых других низших беспозвоночных: с одной стороны данных довольно мало, с другой - представители этого типа из-за пелагического образа жизни сильно оводнены. Средняя величина содержания воды в теле гребневиков (по данным Лазаревой, 1961; Ikeda, 1970; Ивлевой, Литовченко, 1978) равна 97.5± 0.5% (N=10, п=11). Так как мы принимаем, что эталонное содержание воды в теле водных животных без учета приспособления к пелагическому образу жизни равно 86.5%, то в данные о коэффициенте упорядоченности для гребневиков внесена поправка, рассчитанная по формуле (80).
Все известные нам материалы по стандартному обмену и критерию упорядоченности гребневиков представлены в табл.7.
Таблица 7 Масса тела M (г), интенсивность потребления кислорода 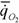 (мл/г-ч) при
(мл/г-ч) при
температуре опыта t°C , коэффициент а (мВт/г) и критерий упорядоченности Сr у представителей типа Ctenophora(Гребневики).
| Виды | M | 
| t° | Авторы | a | Cr |
| Beroe ovenereis | 0.00050 | (1)* | 0.0156 | 0.23 | ||
| Cestus veneris | 0.00375 | (1) | 0.0609 | 0.99 | ||
| 0.0029 | (1) | 0.0471 | 0.77 | |||
| Hormiphora | 0.43 | 0.027 | 6.7 | (2) | 0.379 | 6.17 |
| palmata | 0.26 | 0.037 | 6.3 | (2) | 0.472 | 7.70 |
| Pleurobrachia | 0.072 | 0.053 | (3) | 0.072 | 1.94 | |
| globosa | 0.10 | 0.050 | (3) | 0.073 | 1.98 | |
| Pleurobrachia plieus | 0.25 | 0.0147 | (4) | 0.053 | 1.45 | |
| 0.25 | 0.0014 | (4) | 0.020 | 0.54 | ||
| 0.18 | 0.0133 | (6) | 0.049 | 1.31 | ||
| 0.268 | 0.0093 | (6) | 0.038 | 1.02 |
* Авторы: (1) Ledebur, 1939; (2) Ikeda.1970; (3) Rajagopal, 1962; (4) Лазарева, 1961; (5) Павлова, 1968; (6) Кузьмичова, 1980.
При расчете критерия упорядоченности учитывалось сильная оводненность этих животных. Для типа гребневиков в целом получили следующие величины коэффициента а и критерия упорядоченности Сr.
Тип Ctenophoraа = 0.113±0.043; Сr=2.17±0.67 (88)
Тип Coelenterata(Кишечнополостные)
Для типа Coelenterataв литературе имеется значительно больше данных об интенсивности потребления кислорода животными, чем для типа Porifera или Ctenophora. Но и здесь при рассмотрении описанных в литературе результатов
сталкиваешься с известными трудностями. Во-первых, разные
авторы измеряли потребление кислорода при разных температурах, и следовало привести их данные к температуре 20°С. Так как данных о влиянии температуры на дыхание представителей этой группы животных недостаточно, мы приводили стандартный обмен к 20°С по обобщенному уравнению Крога (69). Во-вторых, многие авторы рассчитывали интенсивность дыхания не на сырую, а на сухую массу, что затрудняет сравнение стандартного обмена кишечнополостных как внутри типа, так и с представителями других типов животных. Чтобы пересчитать потребление кислорода на сырую массу мы приняли во внимание, что содержание воды в теле взрослых кишечнополостных по данным, приводимым в работах Виноградова (1934), Яковлевой (1964), Ikeda (1970), Ивлевой, Литовченко (1978) и Ивлевой (1981) равно: в классе Hydrozoa (Гидроидные) - 96.4+0.4% (N=13, n=18); в классе Siphonophora (Сифонофоры) - 96.0+0.6% (N=9, n=10); в классе Scyphozoa (Сцифоидные) - 97.0±0.4% (N=11, n=16) и в классе Anthozoa (Коралловые полипы) - 84.0±0.9% (N=11, n=15).
Обработанные указанным способом данные о скорости потребления кислорода различными видами кишечнополостных приведены на рис. 14(2). По расчетам Ивлевой и Литовченко (1978) у Hydrozoa и Scyphozoa k = 0.66. Нам кажется, что это значение коэффициента k занижено. И действительно, расчет по данным, приводимым на рис. 14(2) показал, что аллометрическая зависимость скорости потребления кислорода от массы тела у кишечнополостных (n=36) имеет вид:
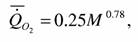
где коэффициент k=0.078±0.022, т.е.достоверно не отличается от эталонной величины k=0.75.
При вычислении критерия упорядоченности Сr учитывалась также, что многие кишечнополостные, как показано выше, сильно оводнены, что связано с приспособлением к пассивному пелагическому образу жизни. Для того, чтобы получить для них критерий упорядоченности и сравнить его с критерием упорядоченности других животных нужно было привести их массу тела к некоторой сопоставимой величине. Мы это делаем, внося соответствующую
поправку в коэффициент а при вычислении критерия
упорядоченности Сr по формуле (79), с учетом того, что эталонное содержание воды в теле водных животных мы приняли равным 86.5%.
При вычислении критерия упорядоченности следует также учитывать меболитическую компенсацию, т.е. разницу в стандартном обмене кишечнополостных, живущих в умеренном климате по сравнению с тропическим или субтропическим. Расчет коэффициентов а у кишечнополостных, живущих в разных климатических зонах, показал, что аи / at = 1.8, но и в этом случае мы приняли стандартный показатель аи / at = 1.66.
Прежде чем переходить к рассмотрению данных о коэффициенте а и критерии упорядоченности Сr, приводимых в табл.8, следует учесть еще одно замечание. У большинства коралловых полипов очень трудно определить массу колонии, а, следовательно, и сравнить интенсивность дыхания с массой всей колонии. Кроме того, в некоторых случаях в колонии полипов речь идет о симбиозе кишечнополостных с одноклеточными водорослями, что затрудняет определение дыхания самих полипов. Поэтому мы не учитывали довольно многочисленные наблюдения об интенсивности потребления кислорода колониальными кишечнополостными.
Таблица8
Масса тела M (г), интенсивность потребления кислорода 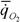 (мл/г•ч) при температуре опыта t°, коэффициент а (мВт/г) и критерий упорядоченности Сr у представителей типа Coelenterata(Кишечнополостные).
(мл/г•ч) при температуре опыта t°, коэффициент а (мВт/г) и критерий упорядоченности Сr у представителей типа Coelenterata(Кишечнополостные).
| Виды | M | 
| t° | Авторы | a | Cr |
| Класс Scyphozoa | 0.179 | 3.48 | ||||
| Aequorea | 3.6 | 0.017** | (3)* | 0.307 | 4.98 | |
| Aurélia aurita | 28.5 | 0.0039 | (4) | 0.051 | 1.36 | |
| 0.0036 | (6) | 0.058 | 1.57 | |||
| 0.0042 | (6) | 0.085 | 2.29 |
| 0.0046 | (7) | 0.099 | 2.65 | |||
| Cassiopea | ||||||
| Xamachana | 0.0125 | (8) | 0.222 | 3.60 | ||
| Cavmarina hastata | 0.00424 | (8) | 0.105 | 1.71 | ||
| 0.0077 | (9) | 0.101 | 1.64 | |||
| Chrysaora | ||||||
| Hysoscella | 0.006 | (10) | 0.184 | 2.99 | ||
| Cyanea sp. | 0.0129 | (10) | 0.320 | 5.19 | ||
| Gonionemus sp. | 0.041** | 0.0533 | (И) | 0.051 | 1.38 | |
| Rhizostoma pulmo | 0.0153 | (12) | 0.201 | 5.41 | ||
| 0.00798 | (9) | 0.245 | 6.60 | |||
| Rhizostoma pulmo | 0.0035 | (4) | 0.116 | 3.13 | ||
| 0.0081 | (6) | 0.361 | 5.67 | |||
| Phizostoma | ||||||
| octopus | 0.0071 | (10) | 0.294 | 4.78 | ||
| Класс Hydrozoa | 0.194 | 3.54 | ||||
| Aglatha digitala | 0.095 | 0.029 | 7.8 | (1) | 0.254 | 4.11 |
| Chelophyes | ||||||
| Appendiculata | 0.103 | 0.036 | (2) | 0.114 | 3.07 | |
| Sarsia mirabilis | 0.373 | 0.017 | 7.6 | (1) | 0.214 | 3.45 |
| Класс Anthozoa | 0.426 | 6.81 | ||||
| Actinia equina | 2.14 | 0.024 | (13) | 0.163 | 3.94 | |
| 10.00 | 0.0134 | (13) | 0.134 | 3.24 | ||
| 0.70 | 0.040 | (14) | 0.172 | 4.17 | ||
| 2.29 | 0.066** | (15) | 0.384 | 9.28 | ||
| 1.67 | 0.047** | (22) | 0.300 | 7.25 | ||
| Anemonia sulcataë | 0.0134 | (16) | 0.294 | 4.29 | ||
| 0.0127 | (9) | 0.279 | 4.07 | |||
| Anthopheura | ||||||
| Elegantissima | 1.14 | 0.088** | (17) | 0.430 | 6.26 |
| Bunodosma | ||||||
| Cavernatu | 2.0 | 0.060** | 22.5 | (18) | 0.323 | 4.71 |
| Calliactis | ||||||
| Parasitica | 8.0 | 0.061** | (19) | 0.807 | 11.77 | |
| Ceriantheopsis | 12.6 | 0.011 | 11.5 | (20) | 0.239 | 3.48 |
| 5.6 | 0.055 | (20) | 0.337 | 4.91 | ||
| Metridium senile | 1.14 | 0.105** | (17) | 0.513 | 7.48 | |
| Pennatula rubra | 4.4 | 0.049** | (19) | 0.558 | 8.14 | |
| Pteroides griseum | 9.26 | 0.024** | (19) | 0.329 | 4.80 | |
| 12.03 | 0.074** | (19) | 0.773 | 11.26 | ||
| Renilla kolliker | 2.73 | 0.124** | 18.5 | (21) | 1.014 | 14.78 |
| Veretillum | ||||||
| Cynomorium | 4.11 | 0.055** | (19) | 0.616 | 8.98 |
* (1) Ikeda, 1970; (2) Павлова, 1975; (3) Arai, 1986; (4) Яковлева, 1964; Павлова, 1968; (6) Кузьмичова, 1980; (7) Аннинский, 1990; (8) McClendon, 1917; (9) Ledebur, 1939; (10) Kriger, 1968; Rajagopal, 1962; (12) Krumbach, 1927; (13) Ивлева, 1964; Виленкина, 1966; (15) Griffiths, 1977; (16) Buddenbroch, 1924; (17) Shicket al., 1979; (18) Ellington, 1982; (19) Brafield, Chapman, 1965; (20) Sassaman, Mangum, 1974; (21) Chapman, 1972; (22) Ивлева, 1981. ** Авторы рассчитывали потребление кислорода на сухую массу животного, мы пересчитали на сырую массу, учитывая данные о содержании воды в теле животного.
Как следует из данных, приводимых в табл.8, наивысший стандартный обмен и критерий упорядоченности наблюдается в классе коралловых полипов, наименьшей - в классе сцифоидных. Поэтому мы можем располагать эти классы в макросистематике кишечнополостных в следующем порядке: