Высшие беспозвоночные
К высшим беспозвоночным животным относят типы членистоногих (Arthropoda), моллюсков (Mollusca), иглокожих (Echinodermata), щетинкочелюстных (Chaetognatha), оболочников (Tunicata) и полухордовых (Hemichordata).
О дыхании представителей некоторых типов высших беспозвоночных в литературе имеется не много данных. Относительно же членистоногих и моллюсков имеется обширная литература. Для каждого из этих двух типов животных можно достаточно точно определить величины коэффициентов а и определить критерий упорядоченности не только для классов и отрядов, но даже для отдельных семейств - особенно в классе насекомых. Кроме того, для насекомых и моллюсков имеются достаточные для наших целей палеонтологические материалы о времени возникновения отдельных отрядов в процессе эволюции. Это позволяет построить кривую биоэнергетического прогресса, аналогичную показанной на рис.6, но уже не для всего животного царства, а отдельно для типа моллюсков или членистоногих и даже для отдельных классов этих типов. Мы рассмотрим, поэтому, данные о стандартном обмене в типе членистоногих и моллюсков с особенной полнотой.
Тип Arthropoda(Членистоногие)
Тип членистоногих включает в себя несколько классов животных, из которых мы рассмотрим меростомовых, многоножек, паукообразных, ракообразных и насекомых. Он содержит до 3 миллионов видов, из которых 90% составляют насекомые (Гиляров, 1968). Данные о стандартном обмене последних достаточно многочисленны, хотя обработка их, как и данных о других членистоногих, из-за
небрежности ряда авторов, измерявших, но не указавших в своих
работах массу изученных животных или приводящих только сухую массу, представляют известные трудности. Есть и другие трудности, о которых подробнее будет сказано в соответствующем месте (чрезмерно развитый хитиновый скелет у жуков и клещей, повышенная температура тела у пчел, связанная со способностью роя пчел к терморегуляции и др.). Все это вносит ошибку в вычисление критерия упорядоченности для отдельных таксонов, которую можно уменьшить только увеличением числа наблюдений о стандартном обмене и внесением соответствующих поправок.
Особенно большую ошибку при определении коэффициента а и критерия упорядоченности Сr вносит малое число наблюдений о дыхании и малый диапазон массы исследованных групп членистоногих. Эту ошибку, как и при изучении других типов животных, можно уменьшить введением единой величины коэффициента k=0.75 для всего типа членистоногих.
По данным Райхле (Reichle, 1968), полученным на основании изучения 16 видов членистоногих, в аллометрической зависимости дыхания от массы тела (46) коэффициент k=0.84. Сущеня (1972) получил для 70 видов ракообразных k= 0.759. Нами использовано значительно больше данных о зависимости скорости потребления кислорода от массы тела у членистоногих (рис.15): для 14 видов многоножек (Вызова, 1972), 173 видов паукообразных (Зотин и др., 1998а), 168 видов ракообразных (Алексеева, 1999) и 384 видов насекомых (Зотин, Алексеева, 1997) - всего для 739 видов членистоногих. На основании этих наблюдений можно было рассчитать коэффициенты в аллометрической зависимости (46) по методике, описанной ранее (Зотин, 1999). При расчете данных по всем членистоногим величина коэффициента к составила 0.739±0.057, т.е. аллометрическое соотношение для членистоногих равно:
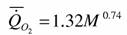
Как видно из этой зависимости коэффициент k для членистоногих достоверно не отличается от принятой нами для всех животных величине k=0.75, которую мы и будем использовать при расчете коэффициента а и критерия упорядоченности Сr для всех видов, семейств, отрядов и классов членистоногих.
Как уже отмечалось, все классы членистоногих содержат очень большое число видов. Особенно многочисленны насекомые (по некоторым высказываниям - больше
двух миллионов видов). Поэтому, хотя для некоторых классов мы приводим не слишком многочисленные данные о стандартном обмене, мы рассмотрим отдельно каждый класс, расположив их в следующей последовательности: меростомовые, многоножки, паукообразные, ракообразные, насекомые. Для насекомых, ракообразных и паукообразных в литературе имеется такое большое число наблюдений о стандартном обмене, что коэффициент а и критерий упорядоченности Сr можно рассчитать не только для отдельных отрядов, но и для некоторых семейств.
Класс Merostomata(Меростомовые)
Дольник (1968) приводит оценки коэффициентов аллометрической зависимости между дыханием и массой тела у представителей отряда меченосцев, расчитанные по данным Г.Г.Винберга (табл.1) [большинство видов этого класса членистоногих вымерли и в современное время сохранились лишь несколько видов отряда Xiphosura (Ланге, 19696)]. Судя по данным, приводимым в табл. 1, аллометрическая зависимость для этого класса членистоногих имеет вид
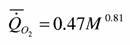
Исходя из этого, можно подсчитать и критерий упорядоченности для класса меростомовых, считая, что представители этого класса обитают в умеренном поясе:
Класс Merostomata a=0.47; Cr=2.64 (91)
Класс Myriapoda(Многоножки)
Данные по скорости потребления кислорода многоножками имеются в литературе в основном для отряда Diplopoda. В табл.10 приведены не очень многочисленные наблюдения для представителей класса многоножек, взятые в основном из обзора Бызовой (1972) и дополненные небольшим числом более новых работ. Все значения приведены к температуре 20°С с помощью кривой Крога. При вычислении критерия упорядоченности мы учитывали, что содержание воды в теле взрослых многоножек по данным Reichle (1968) и Edney (1977) равно 74.0% (N=4, n=4).
Таблица 10
Масса тела M (г), скорость потребления кислорода 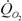
(мВт/животное) при 20°, коэффициент а (мВт/г) при k = 0,75 и критерий упорядоченности Cr y представителей класса Myriapoda (Многоножки) (Вызова, 1972).
| Вид | M | 
| a | Cr |
| Amblyiulus continentalis | 0.140 | 0.038 | 0.166 | 2.37 |
| Chromatoiulus rossicus | 0.143 | 0.120 | 0.516 | 7.38 |
| Glomeris connexa | 0.083 | 0.0572 | 0.370 | 6.28 |
| Japonaria laminata | 0.300 | 0.143 | 0.353 | 5.04 |
| Julus sp. | 0.110 | 0.185 | 0.969 | 13.84 |
| Levizonus thaumasius | 0.280 | 0.157 | 0.408 | 5.83 |
| Pachyulus flavipes | 0.060 | 0.0842 | 0.695 | 9.93 |
| 1.700 | 0.591 | 0.397 | 5.67 | |
| 2.541 | 0.741 | 0.368 | 5.26 | |
| Polydesmus angustus | 0.009 | 0.0136 | 0.465 | 6.65 |
| P. inconstans | 0.008 | 0.0113 | 0.422 | 6.04 |
| Polydesmus sp. | 0.170 | 0.0763 | 0.288 | 4.12 |
| Sarmatiulus kessleri | 0.377 | 0.100 | 0.208 | 2.97 |
| Schizophyllum sabulosum | 0.265 | 0.169 | 0.458 | 6.54 |
| S. caspium | 0.350 | 0.0735 | 0.162 | 2.31 |
| Turanodesmus dmitriewi | 0.010 | 0,0135 | 0.427 | 6.10 |
Опираясь на представленные материалы, получим для класса многоножек (п=16):
Класс Myriapodaa=0.405±0.048; Сг=5.78±0.69 (92)
Класс Arachnida(Паукообразные)
Получено довольно много данных о дыхании разных видов пауков (Anderson, 1970; Greenstone, Bennett, 1980; Anderson, Prestwich, 1982; Prestwich, 1983;), клещей (Webb, 1970; Бызова, 1972; Wood, Lawton, 1973; Luxton, 1975) и других
паукообразных (Phillipson, 1962, 1963; Dresco-Dermout, 1969, 1973; Anderson, 1970; Paul et al., 1989).
Как мы уже не раз отмечали (Зотин, 1988; Алексеева, Зотин, 1995, 1996) у пойкилотермных животных существует, по-видимому, единая кривая зависимости скорости потребления кислорода от температуры. Но пока это не будет доказано достаточно строго имеет смысл пользоваться кривой зависимости дыхания от температуры для отдельных типов или классов животных. Для класса паукообразных была получена зависимость вида (69), но с несколько иным коэффициентом k (Зотин и др., 1999)
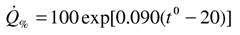
Эта зависимость была использована для расчета стандартного обмена и коэффициента а для всего класса паукообразных, а также для отдельных отрядов, подотрядов и семейств (Зотин и др., 1998а). Как говорилось выше, существует достаточно обширная литература о стандартном обмене паукообразных. В некоторых работах приводится аллометрическая зависимость дыхания от массы тела. В большинстве случаев она получена для сравнительно небольшого числа видов: для 16 видов ракообразных и паукообразных (Reichle, 1968), для 12 (Anderson, Prestwich, 1982) и 26 (Greenstone, Bennett, 1980) видов пауков, для 36 видов клещей (Wood, Lawton, 1973). Вместе с тем в литературе существует значительно больше материалов о дыхании паукообразных. Мы, например, собрали данные о потребления кислорода 173 видов паукообразных (число измерений n=244). К сожалению, многие из цитированных нами авторов (Зотин и др., 1998а), проводили свои измерения не при стандартной температуре и поэтому для вычисления 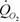
пришлось их данные приводить к температуре 20°С по формуле, приведенной выше. Кроме того, некоторые авторы рассчитывали дыхание на сухую массу паукообразных, не приводя величины сырой массы. В этом случае пришлось воспользоваться данными о содержании воды в теле паукообразных, приводимых в работах Эдни (Edney, 1977), Лакстон (Luxton, 1975) и некоторых других авторов (Chernova et al., 1971, Edgar, 1971, Van Hook, 1971, Block, 1976, Steigen, 1976). Оказалось, что в среднем скорпионы содержат 68.5± 1.08% воды (n=6), пауки - 75.8±1.98% (n=6), клещи - 57.9±1.81 (n=28).
Обычно приводят единую аллометрическую зависимость дыхания от массы тела в изучаемом классе животных. При этом не учитывают, что изучаемый класс
животных может быть гетерогенен по параметрам аллометрических уравнений. Расчет показал, что у паукообразных различные таксоны имеют различные коэффициенты а, в то время как коэффициенты k, достоверно не отличаются и в среднем равны 0.769±0.040. Эта величина достоверно не отличается от константы Хемингсена, поэтому при расчетах коэффициентов а и критерия упорядоченности для отдельных таксонов класса паукообразных мы использовали k=0.75. В этом случае зависимость между скоростью потребления кислорода и массой тела у паукообразных в логарифмической системе координат хорошо аппроксимируется рядом параллельных прямых (рис. 16).
Для расчета критерия упорядоченности Сr [по формуле (74)], как отмечалось, необходимо помимо того, что мы сказали о вычислении коэффициента а, учесть экологические условия жизни и стандартизовать величину живой массы организмов на коэффициент а'. Для стандартизации массы тела паукообразных мы использовали формулу (76), которая включает в себя данные о содержании воды в теле, приведенные выше. Для отрядов, для которых мы не нашли данных о содержании воды в теле, принимается, что оно равно стандартному содержанию, равному 70% Н2О. В отношении экологических условий жизни паукообразных, как и других животных, мы смогли учесть только жизнь в тропическом или умеренном поясах. При этом мы приняли, что соотношение ау/ат=1.66.
Учитывая все сказанное, получим для изученных отрядов паукообразных коэффициенты а и критерии упорядоченности Сr (табл. 11).
Систематика класса Arachnida дана по Ланге (19696). Отряды, подотряды и семейства в табл. 11 расположены в порядке возрастания критерия упорядоченности Сr. Приведенные в табл.11 данные можно поэтому использовать для построения предварительной макросистематики паукообразных. Она несколько отличается в зависимости от того, основываться ли на величине коэффициента а или критерия упорядоченности Сr. В данной работе, в отличие от предыдущей (Зотин и др., 1998а), мы предпочитаем опереться на коэффициент упорядоченности Сr. В этом случае, согласно данным, приводимым в табл.11, отдельные отряды паукообразных можно расположить в следующем порядке: