Acariformes —>Amblypygi —> Scorpiones —> Parasitiformes —> —> Aranei —> Solifugae —> Opiliones
Таблица 11
Коэффициент а (мВт/г) и критерий упорядоченности Сr в разных таксонах класса Arachnida (Паукообразные). N - число видов, n - число измерений (Зотин и др., 1998а).
| Таксон | N | n | a | Cr |
| Отряд Acariformes | 0,277±0,028 | 4,87±0,50 | ||
| Группа сем. Oribatei | 0,214±0,015 | 3,67±0,26 | ||
| CeM.Euphthiracaridae | 0,090 | 1.54 | ||
| Сем. Carabodidae | 0,118 | 2.02 | ||
| Сем. Phthiracaridae | 0,119 | 2.05 | ||
| Сем. Phthiracaridae | 0,119 | 2.05 | ||
| Сем. Phthiracaridae | 0,119 | 2.05 | ||
| Сем. Galumnidae | 0,147 | 2.53 | ||
| Сем. Metrioppiidae | 0,164 | 2.82 | ||
| Сем. Nanhermanniidae | 0,168 | 2.89 | ||
| Сем. Nothridae | 0,175 | 3.00 | ||
| Сем. Damaeidae | 0,175 | 3.01 | ||
| Сем. Camisiidae | 0,178 | 3.06 | ||
| Сем. Mycobatidae | 0,217 | 3.74 | ||
| Сем. Oribatulidae | 0,220 | 3.77 | ||
| Сем. Euzetidae | 0,220 | 3.78 | ||
| Сем. Chamobatidae | 0,231 | 3.98 | ||
| Сем. Belbidae | 0,232 | 3.98 | ||
| Сем. Oribatellidae | 0,238 | 4.08 | ||
| Сем. Achipteriidae | 0,258 | 4.43 | ||
| Сем. Oppiidae | 0,258 | 4.43 | ||
| Сем. Liacaridae | 0,267 | 4.58 | ||
| Сем. Ceratozetidae | 0,340 | 5.85 | ||
| Сем. Nanorchestidae | 0,341 | 5.86 | ||
| Сем. Hypochthoniidae | 0,346 | 5.95 | ||
| Сем. Xenillidae | 0,450 | 7.72 |
| Подотряд Trombidiformes | 0,573±0,159 | 9,84±2,73 | ||
| Сем. Hydrachnidae | 0,033 | 0,57 | ||
| Сем. Halacaridae | 0,155 | 2,67 | ||
| Сем. Tydeidae | 0,276 | 4,74 | ||
| Сем. Stigmaeidae | 0,302 | 5,18 | ||
| Сем. Scutacaridae | 0,486 | 8,34 | ||
| Сем. Bdellidae | 0,654 | 11,23 | ||
| Сем. Rhagidiidae | 0,868 | 14.91 | ||
| Сем. Eylaidae | 1,400 | 24,06 | ||
| Группа ceм. Acaridiae Сем. Acaridae | 0,588±0,152 | 12,32±1,87 | ||
| Отряд AmblypygiСем. Tarantulidae | 0,228±0,032 | 5,72±0,80 | ||
| Отряд Scorpiones | 0,272±0,023 | 6,84±0,58 | ||
| Сем. Buthidae | 0,239 | 6,00 | ||
| Сем. Scorpionidae | 0,258 | 6,49 | ||
| Сем. Chactidae | 0,407 | 10,23 | ||
| Сем. Vejovidae | 0,630 | 15,84 | ||
| Отряд Parasitiformes | 0,513±0,084 | 8,82±1,44 | ||
| Надсем. Uropodoidea | 0, 197±0,021 | 3,38±0,36 | ||
| Сем. Trachytidae | 0,157 | 2,70 | ||
| Сем. Uropodidae | 0,173 | 2,98 | ||
| Сем. Epicridae | 0, 197 | 3,38 | ||
| Сем. Pachylaelaptidae | 0,320 | 5,50 | ||
| Надсем. Gamasoidea | 0,617±0,077 | 10,60±1,33 | ||
| Сем. Veigaiaidae | 0,412 | 7,09 | ||
| Сем. Eviphididae | 0,441 | 7,58 | ||
| Сем. Parasitidae | 0,573 | 9,84 | ||
| Сем. Rhodacaridae | 0,676 | 11,62 | ||
| Сем. Macrochelidae | 1,008 | 17,32 | ||
| Надсем. Ixodoidea | 0,716±0,323 | 12,29±5,56 | ||
| Сем. Ixodidae | 0,201 | 3,36 |
| Сем. Argasidae | 1,230 | 21,13 | ||
| Отряд Aranei (Пауки) | 0,601±0,048 | 11, 19±0,77 | ||
| Подотряд Liphistiomorphae Сем. Liphistiidae | 0,200 | 2,80 | ||
| Подотряд Mygalomorphae | 0,232±0,011 | 5,40±0,25 | ||
| Сем. Ctenizidae | 0,163 | 3,80 | ||
| Сем. Plectreuridae | 0, 199 | 4,63 | ||
| Сем. Filistatidae | 0,216 | 5,03 | ||
| Сем. Theraphosidae | 0,251 | 5,86 | ||
| Сем. Gnaphosidae | 0,269 | 6,27 | ||
| Сем. Atypidae | 0,276 | 6,44 | ||
| Сем. Clubionidae | 0,290 | 6,75 | ||
| Подотряд Araneomorphae | 0,690±0,053 | 12,65±0,86 | ||
| Сем. Sicariidae | 0,335 | 7,81 | ||
| Сем. Pisauridae | 0,612 | 8,59 | ||
| Сем. Lycosidae | 0,611 | 9,86 | ||
| Сем. Salticidae | 0,495 | 10,60 | ||
| Сем. Oxyopidae | 0,517 | 12,06 | ||
| Сем. Thomisidae | 0,564 | 13,13 | ||
| Сем. Araneidae | 0,685 | 13,50 | ||
| Сем. Theridiidae | 0,834 | 19,43 | ||
| Сем. Agelenidae | 0,890 | 20,74 | ||
| Сем. Linyphiidae | 1,694 | 25,79 | ||
| Отряд Solifugae (Сольпуги) Сем. Eremobatidae | 0,746±0,047 | 17,39±1,08 | ||
| Отряд Opiliones | 3,019±0,493 | 48,51±6,18 | ||
| Сем. Ischyropsalidae | 1,838 | 46,22 | ||
| Сем. Phalangiidae | 3,526 | 49,49 |
Конечно, требуется дальнейшая работа по построению макросистемы паукообразных. Пример, приведенный в табл.11, показывает, что это можно сделать, используя знание величин сопоставимого стандартного обмена, хотя и очевидно, что требуется дальнейшее накопление данных о коэффициенте а и критерии
упорядоченности Сr. Последнее требует получения дополнительных материалов о содержании воды в теле разных видов паукообразных и тщательное исследование их экологии.
Суммируя данные, приведенные в табл.11, мы можем записать в настоящее время следующие величины коэффициента а и критерия упорядоченности Сr для класса паукообразных в целом (n=244):
Класс Arachnidaa=0.530±0.045; Сr=9.64±0.70 (93)
Класс Crustacea(Ракообразные)
Данных о дыхании ракообразных в литературе имеется даже больше, чем для паукообразных, не говоря уж о меростомовых или многоножках (Винберг, 1950, Ивлев, 1963, Сущеня, 1972, Алексеева, 1999). Несмотря на это, при их использовании встают те же трудности, что и случае других классов членистоногих.
Таблица 12
Содержание воды в теле представителей разных отрядов класса ракообразных - Crustacea N - число видов, n - число измерений [по данным Виноградова (1937), Ikeda (1970) и Ивлевой (1981)].
| Отряд | H2O | N | n |
| Amphipoda | 81.8 | ||
| Branchiopoda | 90.7 | ||
| Copepoda | 82.3 | ||
| Cumacea | 76.8 | ||
| Cirripedia | 76.2 | ||
| Decapoda | 74.3 | ||
| Euphausiacea | 77.9 | ||
| Isopoda | 72.7 | ||
| Mysidacea | 70.0 | ||
| Ostracoda | 79.7 | ||
| Schizopoda | 77.8 | ||
| Stomatopoda | 72.0 |
Во-первых, многие авторы проводили свои измерения не при
температуре 20°С, что необходимо делать при определении стандартного обмена пойкилотермных животных. Для приведения таких измерений к температуре 20°С можно было бы использовать уравнение Крога, приводимое Сущеней (1972):
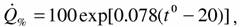
но мы предпочли использовать данные, собранные Алексеевой (1999), согласно которой
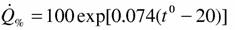
Во-вторых, часто приводят данные о дыхании в расчете на сухую массу ракообразных. Для того чтобы пересчитать эти данные на сырую массу, как принято в нашей работе, пришлось воспользоваться материалами о содержании воды в теле представителей разных отрядов ракообразных (табл. 12). В среднем для всего класса содержание воды в теле ракообразных, судя по табл.12, равно 77.7±0.7% (N=68, n=83).
В третьих, большинство ракообразных имеют развитые наружные покровы и при вычислении критерия упорядоченности также приходится учитывать содержание воды в теле согласно формуле (78).
Как отмечалось, данных о дыхании ракообразных довольно много. Это позволило определить некоторым авторам коэффициент k в аллометрической зависимости скорости потребления кислорода от массы тела с достаточной точностью. Согласно расчетам Сущени (1972), проведенным на основании данных о дыхании 70 видов ракообразных k=0.759. Алексеева (1999), используя данные о дыхании 188 видов ракообразных (п=487), получила k=0.713±0.035 (рис. 17). Эти величины достоверно не отличаются от принятого нами значения коэффициента k=0.15 для всех животных.
В табл.13 представлены данные о коэффициенте а для разных таксонов класса ракообразных, полученные Алексеевой (1999), и критерии упорядоченности этих же таксонов, рассчитанные нами. В тех случаях, когда нам были известны данные о климатическом поясе, в котором жили изученные виды ракообразных, мы использовали соотношение ау/ат=1.66 для тропических видов и αу/αT=1 во всех остальных случаях.
Таблица 13.
Коэффициент а (мВт/г) (Алексеева, 1999) и критерий упорядоченности Сr в разных отрядах класса Crustacea (Ракообразные) N - число изученных видов, n - число измерений.
| Таксон* | а | N | n | Сr |
| Подкласс Malacostraca | 1.068±0.079 | 18.40 ± l.31 | ||
| Отряд Cumacea | 0.318 | 6.89 | ||
| Отряд Mysidacea | 0.743±0.201 | 13.32±3.52 | ||
| Отряд Decapoda | 0.770±0.055 | 14.61±1.05 | ||
| Отряд Spelaogriphacea | 1.047 | 16.90 | ||
| Отряд Isopoda | 1.089±0.258 | 18.49±4.37 | ||
| Отряд Tanaidacea | 0.911 | 19.57 | ||
| Отряд Amphipoda | 1.362±0.181 | 21.46±2.85 | ||
| Отряд Euphasiacea | 1.976±0.311 | 32.51±4.99 | ||
| Подкласс Maxillopoda | 1.185±0.073 | 20.32±1.92 | ||
| Отряд Cirripeda | 0.191±0.085 | 3.12±1.39 | ||
| Подотр. Thoracica | 0.191±0.085 | 3.12±1.39 | ||
| Отряд Copepoda | 1.292±0.123 | 22.17±2.04 | ||
| Подотр. Harpacticoida | 0.835 | 12.91 | ||
| Подотр. Cyclopoida | 1.287 | 19.91 | ||
| Подотряд Calanoida | 1.297±0.127 | 22.34±2.11 | ||
| Подкласс Ostracoda | 1.129 | 30.26 | ||
| Отряд Ostracoda | 1.129 | 30.26 | ||
| Подкласс Branchiopoda | 2.502±0.481 | 41.71±8.03 | ||
| Отряд Phyllopoda | 1.769±0.382 | 30.15±6.73 | ||
| Подотр. Cladocera | 1.078±0.330 | 19.49±6.56 | ||
| Подотр. Notostraca | 2.395±0.447 | 38.67±7.21 | ||
| Подотр. Conchostraca | 6.491 | 104.84 | ||
| Отряд Anostraca | 5.955±1.586 | 122.39±10.31 |
* Классификация дана по работе Бирштейна (1968)
Макросистематику для отрядов и подотрядов ракообразных, данные о критерии упорядоченности которых показаны в табл.13, можно представить в следующем виде:
Подкласс Malacostraca
Cumacea —> Mysidacea —> Decapoda —> Spelaogriphacea —> —> Isopoda —> Tanaidacea —> Amphipoda —> Euphasiacea
Подкласс MaxillopodaCirripeda —> Copepoda (Harpacticoida —> Cyclopoida —> Calanoida)
Подкласс OstracodaOstracoda
Подкласс BranchiopodaPhyllopoda (Cladocera —> Notostraca —> Conchostraca) —> Anostraca
В целом для класса ракообразных имеем:
Класс Crustaceaa=1.208±0.073, Сг=20.72±1.22 (94)
Класс Insecta (Насекомые)
Долгое время в класс насекомых включали два подкласса низших и высших насекомых, а также инфракласс щетинохвосток (Бей-Биенко, 1980). Однако позднее низших насекомых и щетинохвосток выделили в отдельные классы и в классе Insecta оставили только крылатых насекомых (Росс и др., 1985). Чтобы упростить свою задачу мы будем придерживаться старой таксономии и начнем свое рассмотрение с низших насекомых. Однако прежде чем переходить к обсуждению биоэнергетического прогресса и прогрессивной эволюции в классе насекомых, следует обсудить трудности, с которыми связано сопоставление данных по стандартному обмену насекомых. Они во многом те же, что и для других беспозвоночных.
Во-первых, многие авторы проводили свои измерения не при 20°С, и, как отмечалось, приходится приводить их данные по кривой Крога к этой
температуре. Как говорилось, по нашему мнению, для всех пойкилотермных животных существует единая зависимость потребления кислорода от температуры (67). Но пока это утверждение не доказано достаточно строго, мы будем использовать кривые Крога для отдельных отрядов насекомых, полученные нами ранее (Табл.2). Для всех остальных насекомых будем использовать единую кривую Крога (69) (Зотин, Алексеева, 1997) вида:

По этой формуле мы рассчитывали дыхание при 20°С для тех видов насекомых, для которых она определена при иной температуре.
Во-вторых, нам удалось собрать достаточно подробные данные о дыхании представителей 10 отрядов насекомых (они включают в себя наиболее распространенные виды) и менее подробные о 2 отрядах (Донцова и др., 1985; Алексеева и др., 1993; Алексеева, Зотин, 1995, 1996, Зотин, Алексеева, 1997). Эти данные показали, что аллолометрическая зависимость потребления кислорода от массы тела у насекомых, показанная на рис. 15, не является одной и той же для всех отрядов. Как видно из рис.18, каждый из отрядов имеет свой коэффициент а и k в аллометрической зависимости (46). В то же время расчет показывает, что согласно данным, приведенных на рис.15, аллометрическое соотношение для класса насекомых в целом выражается соотношением:
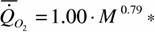
где а=0.985±0.053, k=0.794±0.068. Величина k достоверно не отличается от константы Хемингсена, т.е. для расчетов коэффициента α и критерия упорядоченности Сr отдельных отрядов и семейств насекомых можно использовать единый для всех животных коэффициент k = 0.75. Это позволяет стандартизировать данные о коэффициенте а при вычислении критериев упорядоченности отдельных семейств и отрядов насекомых и сделать некоторое общие заключения о прогрессивной эволюции в классе насекомых.
В-третьих, разные группы насекомых имеют хитиновые покровы развитые в разной степени. Хитиновый покров насекомых не участвует в окислительных реакциях и в то же время входит в определение сырой и сухой массы при ее измерении путем взвешивания. Это следует учитывать при расчете критерия упорядоченности, так как при определении коэффициента а по формуле (74) в
нее вводят данные о массе насекомых. Поэтому при определении критерия упорядоченности Сr, по которому мы судим насколько те или иные группы животных продвинулись по пути прогресса, мы вынуждены использовать косвенный прием, основанный, как и в случае с низшими беспозвоночными, на определении содержания воды в теле насекомых, приводя их дыхание к условному организму с содержанием воды в 70% - цифру, характерную для большинства наземных животных (Culebras et al., 1977; Edney, 1977).
Для того, чтобы осуществить это, нам естественно необходимо знать содержание воды в теле представителей отдельных таксонов насекомых. Мы провели такие определения и собрали литературные данные (Табл.14). Как видно из таблицы, содержание воды в теле насекомых колеблется от 58 до 82%. Для тех отрядов, для которых мы не нашли данных о содержании воды в теле, мы использовали среднюю цифру, полученную для 56 видов из табл. 14, равную 67.0%. За эталонное содержание воды при расчете критерия упорядоченности мы принимали, как и для других наземных животных, величину 70%. При вычислении критерия упорядоченности (74) для насекомых это учитывалось по формуле (76).
Таблица 14.
Содержание воды в теле взрослых насекомых (Зотин. Алексеева, 1997). N - число изученных видов, n - число измерений.
| Отряд | H2О(%) | N | n |
| Collembola (Ногохвостки) | 82.2 | ||
| Thysanura (Щетинохвостки) | 70.0 | ||
| Orthoptera (Прямокрылые) | 68.7±1.5 | ||
| Blattodea (Тараканы) | 69.3 | ||
| Copeognatha (Сеноеды) | 66.0 | ||
| Isoptera (Термиты) | 71.5 | ||
| Homoptera (Равнокрылые) | 69.0 | ||
| Hemiptera (Клопы) | 65.3 | ||
| Plecoptera (Веснянки) | 70.0 | ||
| Coleoptera (Жуки) | 58.3±2.6 |
| Lepidoptera (Бабочки) | 63.9±3.1 | ||
| Hymenoptera (Перепончатокрылые) | |||
| Сем. Apidae (Пчелы) | 62.1 | ||
| Сем. Formicidae (Муравьи) | 65.1±1.0 | ||
| Diptera (Двукрылые) | 14.1+2.2 |
В-четвертых, при определении коэффициента упорядоченности следует учитывать, так называемую, температурную компенсацию у насекомых, живущих в разных климатических зонах (Ивлева, 1981). Поэтому при расчетах коэффициента упорядоченности приходится учитывать климатический пояс обитания вида. В настоящее время мы можем это сделать достаточно грубо, относя одних насекомых к умеренной, других - к тропической зоне. В табл.4 приведены эти данные. Как видно из табл.4, имеются существенные различия в коэффициенте а у представителей тропических насекомых по сравнению с насекомыми умеренной зоны. Эти данные недостаточно надежны. Кроме того, кажется разумным принять, что температурная компенсация одинакова у всех животных. Поэтому мы принимаем, что для всех животных она равна, в среднем, судя по данным, приводимым в табл.4, ау / ат=1.66.
Таким образом, в целом для насекомых при вычислении критерия упорядоченности использовали формулу:
 (95)
(95)