Насекомые с полным превращением
Отряд Coleoptera (Жесткокрылые).
Для этого отряда имеются данные о массе тела и дыхании (117 видов насекомых, 133 измерений) из 19 семейств (Алексеева, Зотин, 1995). Авторы приводят следующие аллометрические зависимости (46) для жуков:

Более точные расчеты дают величину k=0.730±0.050, достоверно не отличающуюся от константы Хемингсена (k=0.75). Мы, исходя из величины k=0.75, рассчитали для семейств жесткокрылых величины коэффициента а (табл.21). Критерий упорядоченности Сг, приводимый в табл.21, вычислен, исходя из наблюдений, что содержание воды в теле жуков равно 58.3% (табл.14), а стандартный обмен жуков умеренного пояса в 1.66 раз больше, чем тропического (хотя согласно данным табл.4 ау/aт=2.80).
Таблица21
Масса тела M (г), скорость потребления кислорода  (мВт), при 20°С (Алексеева, Зотин, 1995), коэффициент а (мВт/г) и критерий упорядоченности Сr для разных семейств отряда Coleoptera класса Insecta. N - число изученных видов, n - число измерений.
(мВт), при 20°С (Алексеева, Зотин, 1995), коэффициент а (мВт/г) и критерий упорядоченности Сr для разных семейств отряда Coleoptera класса Insecta. N - число изученных видов, n - число измерений.
| Семейство | M | 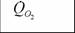
| N | n | a | Cr |
| Cucujidae | 0,000289 | 0,00904 | 0.023 | 0.56 | ||
| Elmidae | 0,000735 | 0,00164 | 0.353 | 6.04 | ||
| Coccinellidae | 0,0172 | 0,0369 | 0.827 | 14.13 | ||
| Curculionidae | 0,0598 | 0,0937 | 0.910 | 15.54 | ||
| Oedemeridae | 0,0095 | 0,0302 | 0.992 | 16.96 | ||
| Dytiscidae | 2,200 | 1,901 | 1.052 | 17.99 | ||
| Carabidae | 0,364 | 0,382 | 1.016 | 18.03 | ||
| Tenebrionidae | 0,529 | 0,460 | 0.936 | 21.01 | ||
| Cantharididae | 0,764 | 0,189 | 1.273 | 21.75 | ||
| Chrysomelidae | 0,0619 | 0,155 | 1.405 | 24.01 | ||
| Scarabaeidae | 2,057 | 3,072 | 1.236 | 27.96 | ||
| Silphidae | 0, 193 | 0,444 | 1.659 | 28.36 | ||
| Rhynchophorida | 0,00336 | 0,0070 | 1.325 | 34.77 | ||
| Passalidae | 0,563 | 0,543 | 0.832 | 39.83 | ||
| Cerambycidae | 1,183 | 1,904 | 1.663 | 41.13 | ||
| Dermestidae | 0,053 | 0,283 | 2.779 | 47.50 | ||
| Meloidae | 0,433 | 1,042 | 1.952 | 55.38 | ||
| Rhipiceridae | 0,291 | 0,581 | 1.466 | 70.17 |
Как видно из табл.21 полученные величины коэффициента а ниже, чем в отрядах прямокрылых и стрекоз. Здесь, однако, следует учитывать следующее. Все жуки обладают очень тяжелым наружным хитиновым скелетом: содержание воды в теле жуков составляет 58.3%, против 62-74% у других насекомых (табл.14). Если учесть это обстоятельство и принять, что массу жесткокрылых при расчетах критерия упорядоченности следует уменьшить на 11.7%, то по этому показателю жуки приближаются к другим насекомым. Итак, для жесткокрылых имеем:
Отряд Coleopteraα=1.235±0.072; Cr=26.08±1.75 (102)
Отряд Lepidoptera(Чешуекрылые).
Для бабочек (отряд чешуекрылых) также получено довольно много наблюдений о скорости потребления кислорода (Zebe, 1954; Bartholomew, Casey,
1978; Алексеева, Зотин, 1996). С учетом измерений, приводимых в последней работе, мы имеем данные о дыхании 74 видов (п=83) из 18 семейств чешуекрылых.
Как было показано выше (рис.18) у бабочек, как и у других насекомых, имеет место типичная аллометрическая зависимость между скоростью потребления кислорода и массой тела. Данных об этой зависимости для бабочек значительно больше, чем для многих других насекомых. Можно, поэтому, рассчитать величину коэффициентов а и k для отряда чешуекрылых (Алексеева, Зотин, 1996):
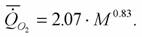
Здесь коэффициент k =0.828+0.051. Учитывая, что эта величина достоверно не отличается от константы Хемингсена, мы принимаем, что k =0.75. Эту цифру мы используем также при расчетах коэффициента а и критерия упорядоченности Сг для отдельных семейств отряда чешуекрылых (табл.22). Следует учитывать, что при вычислении критерия упорядоченности принималось, что содержание воды в теле бабочек равно 63.9% (табл.14). Принималось также, что стандартный обмен насекомых умеренного пояса в 1.66 раз больше стандартного обмена животных тропического пояса.
Таблица 22.
Масса тела M (г), скорость потребления кислорода  (мВт) при 20°С (Алексеевой и Зотина, 1996), коэффициент а (мВт/г) и критерий упорядоченности Сr для разных семейств отряда Lepidoptera класса Insecta. N - число изученных видов, n - число измерений.
(мВт) при 20°С (Алексеевой и Зотина, 1996), коэффициент а (мВт/г) и критерий упорядоченности Сr для разных семейств отряда Lepidoptera класса Insecta. N - число изученных видов, n - число измерений.
| Семейство | M | 
| N | n | a | Cr |
| Satyridae | 0.049 | 0.111 | 1.071 | 17.10 | ||
| Alucitidae | 0.0097 | 0.039 | 1.262 | 20.15 | ||
| Liparidae | 0.242 | 0.414 | 1.312 | 20.96 | ||
| Tineidae | 0.018 | 0.730 | 1.467 | 23.43 | ||
| Zygaenidae | 0.039 | 0.145 | 1.655 | 26.44 | ||
| Lycaenidae | 0.055 | 0.185 | 1.689 | 26.97 |
| Noctuidae | 0.258 | 0.349 | 1.565 | 27.68 | ||
| Lasiocampidae | 0.249 | 0.490 | 1.556 | 31.06 | ||
| Hesperiidae | 0.046 | 0.205 | 2.061 | 32.91 | ||
| Pieridae | 0.048 | 0.216 | 2.071 | 33.09 | ||
| Nymphalidae | 0.185 | 0.751 | 2.545 | 40.65 | ||
| Megalpygidae | 0.627 | 1.133 | 1.608 | 42.64 | ||
| Cossidae | 1.772 | 2.962 | 1.929 | 45.59 | ||
| Eriocraniidae | 0,015 | 0.075 | 1.752 | 46.40 | ||
| Pyralididae | 0.053 | 0.227 | 2.485 | 46.57 | ||
| Sphingidae | 1.236 | 2.402 | 2.177 | 53.01 | ||
| Saturniidae | 0.837 | 2.418 | 2.465 | 55.67 | ||
| Notodontidae | 0.367 | 0.942 | 2.126 | 56.38 | ||
| Bombycidae | 0.457 | 0.538 | 2.888 | 76.58 |
Учитывая сказанное, можно принять, что коэффициент а и критерий упорядоченности Сr для отряда чешуекрылых равен:
Отряд Lepidopteraα=2.096±0.11; Cr=44.09±2.38 (103)
Отряд Diptera (Двукрылые).
В табл.23 представлены средние величины коэффициента а и критерия упорядоченности Сr для отдельных семейств отряда двукрылых. Эти величины основаны на данных о стандартном обмене 66 видов (123 измерения), представляющих 14 семейств отряда Diptera (Алексеева и др., 1993). Согласно вычислениям авторов коэффициенты в аллометрическом соотношении для двукрылых равны
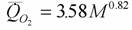
Наши вычисления дают величину k=0.770±0.011. Коэффициент k и в этом случае статистически не отличается от принятого нами для всех животных k=0.75. При вычислении критерия упорядоченности Сr учтено также, что содержание воды в теле представителей отряда двукрылых составляет 74.7% (табл.14) и что
двукрылые умеренного пояса дышат в 1.89 (табл.4) раз сильнее тропических. Исходя из этого, получим
Отряд Dipteraα=2.894±0.11; Cr=48.43±1.84 (104)
Это очень высокая величина сопоставимого стандартного обмена а и критерия упорядоченности Сr, возможно самая высокая среди насекомых (исключая, конечно, некоторых насекомых, способных создавать суперорганизмы, о которых речь пойдет ниже). Таким образом, подтверждается положение, высказанное Тыщенко (1976), согласно которому двукрылые обладают среди насекомых одним из самых высоких уровней стандартного обмена.
Таблица 23.
Масса тела M (г), скорость потребления кислорода  (мВт) при 20°С (Алексеева и др., 1993), коэффициент а (мВт/г) и критерий упорядоченности Сr в разных семействах отряда Diptera класса Insecta. N - число изученных видов, n - число измерений.
(мВт) при 20°С (Алексеева и др., 1993), коэффициент а (мВт/г) и критерий упорядоченности Сr в разных семействах отряда Diptera класса Insecta. N - число изученных видов, n - число измерений.
| Семейство | M | 
| N | η | a | Cr |
| Stratiomyidae | 0,100 | 0,1515 | 0.852 | 12.15 | ||
| Anthomyiidae | 0,0132 | 0,0586 | 1.505 | 21.46 | ||
| Hippobosicidae | 0,0055 | 0,0352 | 1.743 | 24.86 | ||
| Chloropidae | 0,0305 | 0,1667 | 2.285 | 32.59 | ||
| Pantophthalmidae | 1.746 | 2,0190 | 1.329 | 35.83 | ||
| Drosophilidae | 0,0013 | 0,0144 | 2.219 | 36.96 | ||
| Culicidae | 0,0016 | 0,0215 | 2.524 | 43.61 | ||
| Tabanidae | 0,112 | 0,6215 | 3.319 | 47.34 | ||
| Cordyluridae | 0.0309 | 0.246 | 3.341 | 47.65 | ||
| Scatophagidae | 0,0309 | 0,2462 | 3.341 | 47.65 |
| Tachinidae | 0,0567 | 0,4059 | 3.523 | 50.25 | ||
| Muscidae | 0,0155 | 0,1738 | 3.965 | 57.80 | ||
| Asilidae | 0,0903 | 0,4537 | 3.078 | 72.86 | ||
| Simuliidae | 0,0023 | 0,0549 | 5.249 | 74.87 | ||
| Tipulidae | 0,101 | 1,208 | 6.545 | 93.35 |
Отряд Hymenoptera(Перепончатокрылые).
Отряд перепончатокрылых помимо обычных сложностей, связанных с небрежностями авторов при исследовании дыхания беспозвоночных, представляет особые трудности для выяснения сопоставимого стандартного обмена и критерия упорядоченности, так как некоторые представители семейства пчелиных, входящих в этот отряд, обладают способностью к терморегуляции (Heinrich, 1981a,b; Chappell, 1984). Кроме того, многие пчелы и муравьи образуют, так называемые, суперорганизмы - рои, гнезда и муравейники, способные к поддержанию внутри высокой температуры отличной от окружающей среды. Все это заставляет по разному решать вопрос о коэффициенте а и критерии упорядоченности Сr для отдельных, изолированных особей и особей объединенных в коллектив - для суперорганизмов.
Индивидуальные перепончатокрылые. Следует прежде всего решить вопрос как быть с пчелами. Они, как говорилось, обладают способностью к терморегуляции не только в коллективе, но и в одиночном состоянии (Allen, 1959, Cahill, Lustik, 1976; Heinrich, 1981a,b; Chappell, 1984; Southwick, 1990 Moritz, Southwick, 1992). Даже в термонейтральной зоне температура их тела достигает 32-35°С (Cahill, Lustik, 1976; Heinrich, 1981a,b). Очевидно, что эту повышенную температуру тела следует учитывать при определении истинного уровня стандартного обмена пчел. Считая, что при любом измерении дыхания пчел температура их тела равна 35°С, при расчете критерия упорядоченности следует пересчитать скорость их дыхания на температуру 20°С с помощью кривой Крога (68). Это сделано в табл.24 при рассмотрении данных о стандартном обмене и коэффициенте упорядоченности. В этой таблицы данные о стандартном обмене пчел и ос (Сем. Apidae и Vespidae) приведены в расчете на
температуру среды равную 20°С, а при вычислении критерия упорядоченности внесена поправка, связанная с тем, что температура тела пчел равна 35°С.
При расчете средней величины массы тела и скорости потребления кислорода представителей этих семейств не приняты во внимание данные о Bombus vosnesenskii (Kammer, Heinrich, 1974) ввиду уникальности их массы - до 8.8 г, не характерной для подавляющего большинства представителей этого семейства (Алексеева, Зотин, 1996). Аллометрическая зависимость дыхания от массы тела (46) у пчел заметно отличается от муравьев (рис.18), поэтому мы рассчитали ее отдельно для семейства пчелиных (Алексеева, Зотин, 1995):

Таблица 24.
Масса тела M (г), скорость потребления кислорода при 20°С  (мВт), коэффициент а (мВт/г) и критерий упорядоченности Сr для разных семейств отряда Hymenoptera (Перепончатокрылые) класса Insecta. N - число изученных видов, n - число измерений (по данным, приводимым в обзоре Алексеевой и Зотина, 1996).
(мВт), коэффициент а (мВт/г) и критерий упорядоченности Сr для разных семейств отряда Hymenoptera (Перепончатокрылые) класса Insecta. N - число изученных видов, n - число измерений (по данным, приводимым в обзоре Алексеевой и Зотина, 1996).
| Семейство | M | 
| N | n | a | Cr |
| Dolihoderidae | 0.0028 | 0.0058 | 0.476 | 7.50 | ||
| Mutillidae | 0.0079 | 0.0162 | 0.611 | 11.26 | ||
| Formicidae | 0.012 | 0.0206 | 0.654 | 12.59 | ||
| Apidae | 0.146 | 0.803 | 4.122 | 73.47 | ||
| Vespidae | 0.229 | 1.948 | 5.815 | 94.79 |
Что касается муравьев (сем. Formicidae), то здесь имеются свои трудности, связанные, во-первых, с тем, что многие исследователи, изучавшие дыхание муравьев, приводят данные о сухой массе и не приводят о сырой. Поэтому, чтобы использовать довольно многочисленные работы по муравьям мы воспользовались данными о содержании сухой массы в процентах от сырой,
приводимыми в некоторых работах: Atta colombica - 37.6% (Lighten et al., 1987), Camponotus herculeanus - 30.0% (Jensen,, 1978), 37.9% (Nielsen et al., 1982), Eciton hamatum - 31.4% (Bartholomew et al., 1988), Formica polyctena - 23.0% (Coenen-Stass et al., 1980), Pogonomyrmex badius - 31.5% (Golley, Gentry, 1964), 38.5% (Porter, 1986), Sclenopsis invicta 32.1% (Elzen, 1986): средняя величина - 32.8% (Алексеева, Зотин, 1995).
Во-вторых, имеется значительный полиморфизм муравьев: известно, что насекомые даже из одного муравейника значительно отличаются друг от друга, как по морфологии, так и по массе. Это ставит вопрос о правомерности использования любых каст муравьев для определения межвидовой величины коэффициента а. Особенно велики различия как по массе так и по интенсивности дыхания рабочих муравьев от плодоносящей самки (королевы) и летающих самцов (см., например, Beraldo, Mendes, 1981). Поэтому при расчетах усредненной массы тела в семействе Formicidae и скорости потребления кислорода мы не использовали данные, полученные на самцах и самках муравьев. Аллометрическая зависимость для муравьев выражается формулой (Алексеева, Зотин, 1996):

Несмотря на то, что мы получили для пчел k=0.91, а для муравьев k=0.85, при расчете коэффициента а и критерия упорядоченности Сr мы, как и для других животных, использовали k=0.75. В конечном счете для семейства пчел и муравьев, а также для всего отряда перепончатокрылых в целом из табл.24 следуют величины коэффициента а и критерия упорядоченности Сr (с учетом данных о содержании воды в теле перепончатокрылых):
Сем. Apidae α=4.122±0.58; Cr=73.47±10.85
Сем. Formicidae α=0.654±0.061; Cr=12.59±1.53 (105)
Отряд Hymenoptera α=1.330±0.18; Cr=24.31±3.30