Макросистематика насекомых
Класс насекомых включает от 24 до 30 отрядов (классификация класса находится еще в процессе становления и точное число отрядов не установлено). Из них мы будем говорить только о 12 наиболее распространенных отрядах, для которых имеются данные о стандартном обмене (табл. 15-24). На их примере мы попытаемся показать, как можно располагать отряды в классе насекомых по критерию упорядоченности.
В настоящее время отсутствуют общепризнанные критерии порядка расположения отрядов в классе насекомых. Это видно хотя бы по тому, как менялось положение отрядов в макросистеме насекомых в разные годы и у разных авторов для указанных 12 отрядов (Зотин, Алексеева, 1997). Каждый автор располагал отряды так, как ему казалось удобным. Было бы правильней, как нам кажется, выработать общие для макросистематики приемы расположения более мелких таксонов внутри более крупных. Большинство авторов считают, что таким критерием являются родственные связи, т.е. степень родства разных таксонов. Это, конечно, правильный путь построения естественной макросистемы животных и растений, но вносит значительный произвол при практическом построении системы. Мы предлагаем
комбинированный путь построения макросистемы: когда очевиден признак выделения крупных групп животных, например, насекомых с полным и неполным превращением, использовать его в качестве основного. В остальных случаях ориентироваться на критерий упорядоченности Сr, как признак, измеряемый количественно, а не качественно. Это уменьшит степень произвола, вносимого в макросистематику животных. Критерий упорядоченности показывает степень удаления организмов от состояния равновесия, от состояния той первичной среды, в которой возникли живые системы на Земле (Зотин, 1984), т.е. он основан на эволюционном принципе. Расположение отрядов в макросистеме класса насекомых с точки зрения указанных принципов осуществлен в табл.25. Тот же принцип, как нам кажется, может быть положен в основу расположения семейств внутри отдельных отрядов насекомых. В таблице 25 отряды и семейства насекомых расположены в соответствии с величиной критерия упорядоченности и эта макросистематика сопоставлена с макросистематикой, приводимой Плавильщиковым (1957). Как видно из этого сопоставления имеется заметное различие между макросистемой, основанной на критерии упорядоченности и принятой некоторыми авторами. Отряд Hymenoptera помещен последним в табл.25, хотя критерий упорядоченности (105) наименьший среди насекомых с полным превращением. Это объясняется тем, что в этом отряде имеются суперорганизмы, коэффициент а которых, а, следовательно, и критерий упорядоченности (106), намного выше, чем у других насекомых.
Таблица25.
Макросистематика отрядов и семейств класса насекомых, основанная на критерии упорядоченности [табл. 15-24 и формулы (97)-(105)], в сопоставлении с макросистематикой, приводимой в определителе Плавильщикова (1957).
| Расположение отрядов и семейства насекомых | ||
| По Плавильщикову | По критерию упорядоченности | Сr |
| Низшие насекомые | ||
| Отряд Collembola | Отряд Collembola | 8.32 |
| Отряд Thysanura | Отряд Thysanura | 8.72 |
| Высшие насекомые | ||
| Насекомые с неполным превращением | ||
| Отряд Blattodea | Отряд Blattodea | 16.51 |
| Отряд Isoptera | Отряд Hemiptera | 19.43 |
| Отряд Hemiptera | Сем. Naucoridae | 5.91 |
| Сем. Coreidae | Сем. Coreidae | 14.86 |
| Сем. Naucoridae | Сем. Pyrrhocoridae | 16.56 |
| Сем. Oncodidae | Сем. Oncodidae | 23.16 |
| Сем. Pentatomidae | Сем. Pentatomidae | 28.04 |
| Сем. Pyrrhocoridae | Отряд Dermoptera | 19.78 |
| Отряд Dermoptera | Отряд Isoptera | 29.14 |
| Отряд Plecoptera | Отряд Plecoptera | 30.15 |
| Отряд Odonata | Отряд Orthoptera | 34.40 |
| Отряд Orthoptera | Сем. Gryllidae | 26.56 |
| Сем. Tettigoniidae | Сем. Acrididae | 35.83 |
| Сем. Gryllidae | Сем. Tettigoniidae | 38.11 |
| Сем. Acrididae | Отряд Homoptera | 36.64 |
| Отряд Homoptera | Отряд Odonata | 44.32 |
| Высшие насекомые Насекомые с полным превращением | ||
| Отряд Coleoptera | Отряд Coleoptera | 26.08 |
| Сем. Carabidae | Сем. Cucujidae | 0.56 |
| Сем. Dytiscidae | Сем. Elmidae | 6.04 |
| Сем. Silphidae | Сем. Coccinellidae | 14.13 |
| Сем. Cucujidae | Сем. Curculionidae | 15.54 |
| Сем. Coccinellidae | Сем. Oedemeridae | 16.96 |
| Сем. Cantharididae | Сем. Dytiscidae | 17.99 |
| Сем. Dermestidae | Сем. Carabidae | 18.03 |
| Сем. Oedemeridae | Сем. Tenebrionidae | 21.01 |
| Сем. Chrysomelidae | Сем. Cantharididae | 21.75 |
| Сем. Meloidae | Сем. Chrysomelidae | 24.01 |
| Сем. Tenebrionidae | Сем. Scarabaeidae | 27.96 |
| Сем. Curculionidae | Сем. Silphidae | 28.36 |
| Сем. Scarabacidae | Сем. Rhynchophoridae | 34.77 |
| Сем. Passalidae | Сем. Passalidae | 39.83 |
| Сем. Elmidae | Сем. Cerambycidae | 41.13 |
| Сем. Rhipiceridae | Сем. Dermestidae | 47.50 |
| Сем. Rhynchophoridae | Сем. Meloidae | 55.38 |
| Сем. Cerambycidae | Сем. Rhipiceridae | 70.17 |
| Отряд Lepidoptera | Отряд Lepidoptera | 44.09 |
| Сем. Eriocranidae | Сем. Satyridae | 17.10 |
| Сем. Zygaenidae | Сем. Alucitidae | 20.15 |
| Сем. Pyralididae | Сем. Liparidae | 20.96 |
| Сем. Tineidae | Сем. Tineidae | 23.43 |
| Сем. Pieridae | Сем. Zygaenidae | 26.44 |
| Сем. Alucitidae | Сем. Lycaenidae | 26.97 |
| Сем. Lycaenidae | Сем. Noctuidae | 27.68 |
| Сем. Hesperiidae | Сем. Lasiocampidae | 31.06 |
| Сем. Nymphalidae | Сем. Hesperiidae | 32.91 |
| Сем. Cossidae | Сем. Pieridae | 33.09 |
| Сем. Satyridae | Сем. Nymphalidae | 40.65 |
| Сем. Noctuidae | Сем. Megalpygidae | 42.64 |
| Сем. Bombycidae | Сем. Cossidae | 45.59 |
| Сем. Lasiocampidae | Сем. Eriocraniidae | 46.40 |
| Сем. Saturniidae | Сем. Pyralididae | 46.57 |
| Сем. Sphingidae | Сем. Sphingidae | 53.01 |
| Сем. Notodontidae | Сем. Saturniidae | 55.67 |
| Сем. Liparidae | Сем. Notodontidae | 56.38 |
| Сем. Megalpygidae | Сем. Bombycidae | 76.58 |
| Отряд Diptera | Отряд Diptera | 48.43 |
| Сем. Tipulidae | Сем. Stratiomyidae | 12.15 |
| Сем. Culicidae | Сем. Anthomyiidae | 21.46 |
| Сем. Simuliidae | Сем. Hippobosicidae | 24.86 |
| Сем. Stratiomyidae | Сем. Chloropidae | 32.59 |
| Сем. Pantophthalmidae | Сем. Pantophthalmidae | 35.83 |
| Сем. Tabanidae | Сем. Drosophilidae | 36.96 |
| Сем. Cordyluridae | Сем. Culicidae | 43.61 |
| Сем. Chloropidae | Сем. Tabanidae | 47.34 |
| Сем. Asilidae | Сем. Scatophagidae | 47.65 |
| Сем. Anthomyiidae | Сем. Tachinidae | 50.25 |
| Сем. Muscidae | Сем. Muscidae | 57.80 |
| Сем. Tachinidae | Сем. Asilidae | 72.86 |
| Сем. Scatophagidae | Сем. Simuliidae | 74.87 |
| Сем. Hippoboscidae | Сем. Tipulidae | 93.35 |
| Отряд Hymenoptera | Отряд Hymenoptera | 24.31 |
| Сем. Vespidae | Сем. Dolihoderidae | 7.50 |
| Сем. Apidae | Сем. Mutillidae | U. 26 |
| Сем. Formicidae | Сем. Formicidae | 12.59 |
| Сем. Mutillidae | Сем. Apidae | 73.47 |
| Сем. Dolihoderidae | Сем. Vespidae | 94.79 |
Будущее покажет насколько оправдана переделка произвольного расположения отрядов и семейств внутри класса насекомых на макросистематику, основанную на критерии упорядоченности, т.е. в конечном счете, на данных об энергетическом метаболизме насекомых.
* * *
Подводя итоги для всего типа членистоногих, получим средние данные, исходя из формул (91), (92), (93), (94) и (107):
Тип Arthropodaa=0.881; Сr =14.38 (108)
Тип Mollusca (Моллюски)
Сложность изучения дыхания моллюсков связана главным образом с определением массы тела, так как большинство моллюсков имеют раковину,
масса которой сильно зависит от экологических условии жизни и возраста животного. Вместе с тем, у некоторых моллюсков раковина отсутствует или сильно редуцирована (голожаберные, головоногие).
Обычно авторы приводят данные о потреблении кислорода в расчете на сухую массу мягких тканей моллюска, а иногда в расчете на общую массы тела моллюска (Алимов, 1981). То и другое не совсем удобно, так как в первом случае мы имеем дело с сухой массой, в то время как для большинства беспозвоночных и особенно позвоночных животных расчет ведется на сырую массу тела, и в результате нельзя сравнивать дыхание моллюсков с дыханием других животных, что является одной из основных задач настоящего исследования. Во втором случае не принимается во внимание то, что раковина моллюсков не потребляет кислорода, в то время как она часто составляет основную часть массы животного, и данные по интенсивности потребления кислорода оказываются сильно заниженными.
По указанным выше причинам мы будем приводить только данные о потреблении кислорода моллюсков, рассчитанных на сырую массу мягких частей тела. В тех случаях, когда авторы не приводят данных о сырой массе тела, мы рассчитывали ее, учитывая содержание воды в мягких тканях. Следует помнить при этом, что содержание воды в мягких тканях представителей разных классов моллюсков несколько отличается. Особенно это относится к классу головоногих моллюсков, поверхностные ткани которых по плотности значительно отличаются от плотности тканей других моллюсков, играя, по-видимому, опорную и защитную функции. В тех случаях, когда нам не известно содержание воды в мягких тканях моллюсков, мы приняли ее равной средней величине для всего типа, которая по данным Виноградова (1937) равна 81,8% (n=84).
Подавляющее большинство моллюсков являются водными животными, поэтому при определении критерия упорядоченности мы приняли в качестве эталона содержание воды в теле равное 86,5%, а метаболитическую адаптацию ау/ат=1.66.
На рис.15 показаны данные о зависимости скорости потребления кислорода от сырой массы мягких тканей у представителей разных классов типа моллюсков. Как видно из этого рисунка аллометрические зависимости дыхания
от массы тела заметно отличаются в разных классах моллюсков, но это в основном касается уровня дыхания, а не коэффициента k. Особенно велики отличия класса головоногих от всех остальных классов моллюсков. Перейдем теперь к более детальному рассмотрению этих данных.
Класс Loricata(Панцирные моллюски)
Панцирные моллюски - это своеобразная группа животных, живущая в зоне приливов и отливов, в зоне прибоя, и их строение и физиология носит явные черты приспособления к этим условиям существования: они имеют обтекаемую форму и способны очень прочно присасываться к поверхности камней и скал (Властов, Матекин, 1968). Оказалось, что в связи с особенностями места существования, потребление кислорода у них отличается в воде и на воздухе (Murdoch, Shumway, 1980). Это вносит известные трудности при сравнении интенсивности дыхания панцирных моллюсков с другими животными. К счастью, судя по данным Мердоха и Шамвей (Murdoch, Shumway, 1980), во-первых, эти различия небольшие и не превышают 20%, во-вторых, кривая аллометрической зависимости дыхания от массы тела у изученных моллюсков в воде и на воздухе перекрещиваются и известна масса моллюсков, при которой их дыхание одинаково в обоих случаях. Для сопоставления дыхания представителей класса панцирных моллюсков с другими животными расчет интенсивности потребления кислорода следует вести на сырую массу мягких частей тела. Так как мы нашли в литературе не много данных по дыханию панцирных моллюсков, то пришлось использовать и те работы, в которых не приведена сырая масса тканей животного. В этом случае мы считали, что сухая масса составляет 19.2% от сырой. В тех случаях, когда авторы приводили только общую сырую массу панцирных моллюсков, включая раковину, мы вычисляли сырую массу тканей по формуле (Murdoch, Shumway, 1980):
МT =0.435•Моб0.969, (109)
где Мт - сырая масса тканей, Моб - общая масса моллюсков.
Учитывая малочисленность наблюдений о дыхании панцирных моллюсков и все сказанное выше, расчет сопоставимого стандартного обмена (коэффициента а) и критерия упорядоченности для панцирных моллюсков , приводимые в табл.26, нельзя считать надежным.
Таблица 26. Масса мягких частей тела М(г), интенсивность потребления кислорода 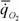 (мл
(мл
О2/г•час) при температуре t°С, a (мВт/г) при температуре 20°С и критерий упорядоченности Cr y представителей класса Loricata (Панцирные).
| Виды | M | 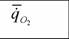
| t0 | Авторы | a | Cr |
| Acanthochiton | 6.05* | 0.0628 | (1)*** | 0.842 | 13.09 | |
| zelandicus | ||||||
| Amaurochiton | 1.86* | 0.162 | (1) | 1.219 | 18.25 | |
| glaucus | ||||||
| Cryptoconchus | 42.86* | 0.0322 | (1) | 0.705 | 10.96 | |
| porosus | ||||||
| Ischnochiton | 3.33* | 0.0664 | (1) | 0.769 | 11.94 | |
| maorianus | ||||||
| Mopalia lignosa | 4.05** | 2.207 | (2) | 0.773 | 19.94 | |
| Nuttallina cflifornica | 0.396** | 0.573 | 13.5 | (3) | 1.148 | 29.61 |
| Onithochiton | 10.42* | 0.0703 | (1) | 1.081 | 16.79 | |
| neglectus | ||||||
| Sypharochiton | 0.50* | 0.116 | (1) | 0.835 | 12.98 | |
| pelliserpontis | ||||||
| Tonicella lintata | 0.308** | 0.050 | (4) | 0.209 | 5.39 | |
| 0.364* | 0.117 | 13.5 | (3) | 0.962 | 24.82 |
Примечания: * Авторы приводят данные о сухой массе, сырую массу мы вычисляли, считая, что содержание воды в теле моллюсков составляет 81.8%. **Масса тканей вычислена по формуле (109). *** Авторы: (1) - Murdoch, Shumway, 1980; (2) Lebsack, 1975; (3) Robbins, 1975; (4) Kincannon, 1975.
Для вычисления критерия упорядоченности использовали формулу (79). При этом принималось, что содержание воды в теле панцирных моллюсков равно 81.8%, а соотношение для тропических и субтропических видов ау/ат=1.66. С учетом этих замечаний имеем для панцирных моллюсков (n=10):
Класс Loricata a=0.802±0.079; Cr=16.45±2.25 (110)
Класс Bivalvia(Двустворчатые моллюски)
Вычисление коэффициента а и критерия упорядоченности Cr y двустворчатых моллюсков затруднено не только тем, что их общая масса определяется авторами, как правило, вместе с раковиной, но и тем, что эти моллюски при работе с ними закрывают створки, захватывая заметное количество воды из среды. Поэтому, когда авторы приводят только общую сырую массу изученных ими двустворчатых моллюсков, приходится вычислять массу мягких тканей, используя косвенный прием: определяя массу раковины и мантийной жидкости по формулам зависимости того и другого от общей массы. К сожалению, таких зависимостей мы нашли в литературе немного, и поэтому по величинам, полученных таким путем, можно получать лишь приблизительные величины сырой массы двустворчатых моллюсков. Для всех представителей Bivalvia мы использовали формулы, полученные для гребешка (расчет по данным Van Dam, 1954):
Pecten grandis Мт=2.16 Моб0,61,
двух видов мидий (Зотин и др. 1987):
Mytilus edulis Mp=0.31·Моб0.95; Мм=0.24•Моб0.96
M. galloprovincialis Μp=0,46·Μоб1·03; MM=0.27·Mo61,02,
двух видов перловиц, двух видов жемчужниц и одного вида беззубок (Зотин A.A., Владимирова И.Г., неопубликовано):
Unio pictorum Мр=0.49·Моб0,96; Мт=0.45·Моб0.91
U. tumidus Мр=0.48·Моб0.97; M=0.28·Mоб1,03
Margaritifera laevis МР=0.37·Моб1.07; ΜT=0.18·Моб0.99
M. margaritifera Mp=0.34·Моб1.09; ΜT=0.16·Μο61.00
Anodonta anatina Mp=0.25·Mo61.04; Мт=0.43·Моб0.91
где Мр - масса раковины; МM - масса воды в мантийной полости; МT - сырая масса тканей; Моб - общая масса моллюска.
Плотное закрытие раковины двустворчатыми моллюсками при малейшей опасности затрудняет определение дыхания этими животными не только потому, что бывает трудно определить массу живых тканей, но и потому, что в закрытом состоянии создаются условия для анаэробиоза и скорость потребления кислорода уменьшается. Например, у Mytilus edulis дыхание «закрытых» мидий ниже
«открытых» приблизительно на 40% (Famme, 1980). Между тем большинство авторов не считаются с тем, насколько открыты раковины моллюсков в то время, когда у них определяют потребление кислорода. Это вносит дополнительную ошибку в определение дыхания двустворчатых моллюсков.
В связи с трудностями определения сырой массы тела двустворчатых моллюсков большинство авторов приводят данные о сухой массе мягких частей тела животного и ведут расчет интенсивности потребления кислорода на сухую массу. Содержание воды в мягких тканях тела двустворчатых моллюсков равно 82.5±0.4% (n=58) (Виноградов, 1937). Поэтому при расчете сырой массы изучаемых моллюсков, когда авторы не приводили ее величину, мы принимали процентное содержание сухой массы в теле равное 17.5%.
Алимов (1975, 1981) на основании данных о дыхании 20 видов двустворчатых пресноводных моллюсков получил следующую аллометрическую зависимость между скоростью потребления кислорода и общей массой тела моллюсков, включающей раковину и мантийную жидкость. Автор выражал скорость потребления кислорода в мг О2/час, мы пересчитали ее в мВт и получили
 = 0.368·М0.721
= 0.368·М0.721
Хотя константа k в этом случае равнялась 0.72, Алимов, как и многие другие авторы, считает, что допустимо принять ее для всех пойкилотермных животных равной 0.75, что мы и сделали в данной работе. В табл. 27 представлены данные о средней величине коэффициента а и критерия упорядоченности Сr в разных таксонах класса двустворчатых моллюсков.
Как следует из табл.27, расположение изученных отрядов в классе двустворчатых моллюсков следует располагать в следующей последовательности: