Caudata -> Gymnophiona -> Anura
Так же располагают отряды амфибий некоторые другие авторы (Наумов, 1950; Матвеев, 1956), но обычное расположение этих отрядов следующее: Gymnophiona - Caudata - Anura (Брэм, 1931; Кашкаров, Станчинский, 1940; Терентьев, 1961; Ананьева и др., 1988), хотя иногда располагают и иначе (Наумов, Карташев, 1979). Это показывает, что у зоологов, как и в случае млекопитающих (Зотин, Владимирова, 1991), нет четких критериев для определения порядка расположения отрядов в классе амфибий. Предлагаемый нами критерий упорядоченности может служить этой цели, хотя, конечно, требуются дополнительные данные в отношении отряда Gymnophiona (табл.35), чтобы судить о справедливости приведенной выше схемы.
Еще труднее из-за ограниченности данных о стандартном обмене построить по критерию упорядоченности макросистематику семейств в отдельных отрядах класса земноводных. Хотя мы расположили различные семейства амфибий в табл.35 в порядке возрастания критерия упорядоченности Сr, это не означает, что этот порядок не претерпит изменений по мере накопления новых данных о стандартном обмене амфибий. Единственно о чем можно пока говорить - это то, что для отряда Caudata предложенное расположение семейств (исключая семейства Cryptobranchidae и
Dicamptodontidae, для которых имеются только единичные
наблюдения о стандартном обмене) не противоречит принятому некоторыми авторами делению этого отряда амфибий на подотряды (Терентьев, 1961). Так все семейства подотряда Salamandroidea по критерию упорядоченности располагаются рядом, а подотряда Ambystomoidea - отдельно (табл.35). То же самое можно сказать о подотряде Procoela (см. Терентьев, 1961) из отряда Anura - все семейства этого подотряда расположились рядом по критерию упорядоченности (табл.35). Иначе обстоит дело с подотрядами Opisthocoela и Diplasiocoea: представители семейств этих подотрядов расползлись в разные концы списка семейств Anura, представленного в табл.35.
Таким образом, требуется еще большая работа для доказательства возможности использования критерия упорядоченности Сr для построения макросистемы в классе Amphibia, но, как нам кажется, приведенные в табл.35 данные о стандартном обмене различных семейств и отрядов амфибий позволяют надеяться, что такое использование возможно.
В целом для класса земноводных коэффициент а и критерий упорядоченности Сr равны:
Класс Amphibiaа = 0.719±0.023; Сг = 12.79±0.38 (123)
Класс Reptilia (Пресмыкающиеся)
Данные о дыхании и массе тела рептилий довольно многочисленны. Они собраны в основном в сводках Темплетона (Templeton, 1970), Беннита и Доусона (Bennett, Dawson, 1976), Андруз и Поу (Andrews, Pough, 1985) и, отчасти, Проссера (1977а) и Шмидт-Нельсена (1982). Во всех этих обзорах приводятся доказательства, что к представителям класса пресмыкающихся применимо аллометрическое соотношение (46).
При изучении аллометрической зависимости скорости потребления кислорода от массы тела у пресмыкающихся разными авторами получены несовпадающие величины коэффициента k: для черепах 0,82 (Hughes et al., 1971) и 0,86 (Bennett, Dawson, 1976); для змей 0,86 (Galvao et al., 1965), 0,66 (Vineger et al., 1970), 0.62 (Dmi'el, Borut, 1972) и 0,77 (Bennett, Dawson, 1976); для ящериц
0,54 (Dawson, Bartholomew, 1956), 0,62 (Bartholomew, Tucker, 1964) и 0,80 (Bennett, Dawson, 1976) для гекконов 0.71 (Feder, Feder, 1981); для отряда чешуйчатых в целом 0.78 (Wilson, 1974) и 0.80 (Andrews, Pough, 1985); для крокодилов 0,69 (Smith, 1976).
Мы собрали литературные данные о массе тела и дыхании 188 видов (342 измерения) пресмыкающихся (рис.25). Расчет адекватным методом (Зотин, 1999) показал, что для всего класса пресмыкающихся выполняется аллометрическая зависимость:
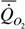 = 0.61M0.79
= 0.61M0.79
Полученный коэффициент k=0.787+0.023 близок величинам, полученным другими авторами, и достоверно не отличается от величины принятой нами в данной работе в качестве стандарта для всех животных, поэтому мы использовали при расчете коэффициента а для представителей класса пресмыкающихся k=0.75. При приведении данных разных авторов к стандартной температуре 20°С мы использовали коэффициент k1=0.085 из формулы (69). Были рассчитаны сопоставимый стандартный обмен (коэффициент а) и критерий упорядоченности Сr для 4 отрядов и 27 семейств пресмыкающихся (табл.36). В таблице отряды и семейства расположены в порядке возрастания критерия упорядоченности. Это, как говорилось, вносит некоторый новый элемент в попытку построения макросистематики, принятой в герпетологии (Тереньтев, 1961; Баранников и др., 1969; Ананьева и др., 1988).
Таблица 36.
Коэффициент а (мВт/г) при 20° С и критерий упорядоченности Сr в разных отрядах и семействах класса Reptilia(рептилии). N - число изученных видов, n - число измерений (Зотин, 1998).
| Таксон | а | Сr | N | n |
| Подкласс Anapsida | 0.810±0.069 | 15.26±1.51 | ||
| Отряд Testudines | 0.810±0.069 | 15.26±1.51 | ||
| Подотряд Trionychoidea | 0.164 | 4.04 | ||
| Сем. Trionychidae | 0.164 | 4.04 |
| Подотряд Cryptodira | 0.814±0.070 | 15.11±1.50 | ||
| Сем. Kinostemidae | 0.384 | 9.49 | ||
| Сем. Chelydridae | 0.510±0.085 | 12.61±2.09 | ||
| Сем. Emydidae | 0,852±0.101 | 15.47±1.52 | ||
| Сем. Testudinidae | 0,911±0.128 | 15.99±3.76 | ||
| Подотряд Cheloniodea | 1.288 | 31.86 | ||
| Сем. Cheloniidae | 1.288 | 31.86 | ||
| Подкласс Lepidosauria | 0.868±0.029 | 18.61±0.52 | ||
| Отряд Rhynchocephalia | 0.510 | 12.62 | ||
| Сем. Sphenodontidae | 0.510 | 12.62 | ||
| Отряд Squamata | 0.868±0.031 | 18.35±0.52 | ||
| Подотряд Amphisbaenia | 0.443 | 10.95 | ||
| Сем. Amphisbaenidae | 0.556 | 13.76 | ||
| Сем. Trogonphidae | 0.215 | 5.33 | ||
| Подотряд Serpentes | 0,819±0.039 | 16.90±0.72 | ||
| Сем. Anomalepididae | 0.494 | 12.22 | ||
| Сем. Crotalidae | 0.603 | 14.92 | ||
| Сем. Viperidae | 0.754±0.105 | 15.54±1.06 | ||
| Сем. Colubridae | 0,864±0.053 | 16,14±0.76 | ||
| Сем. Hydrophidae | 0,719±0.146 | 17.79±3.60 | ||
| Сем. Boidae | 0,827±0.087 | 20.46±2.14 | ||
| Подотряд Lacertilia | 0,905±0.043 | 19,34±0.70 | ||
| Сем. Anguidae | 0.541 | 9.16 | ||
| Сем. Gekkonidae | 0.442±0.032 | 10.93±0.78 | ||
| Сем. Anniellidae | 0.487 | 12.04 | ||
| Сем. Xantusiidae | 0.505±0.065 | 12.49±1.60 | ||
| Сем. Scincidae | 0,588±0.044 | 14.08±0.97 | ||
| Сем. Teiidae | 0.672 | 16.63 | ||
| Сем. Iguanidae | 0,866±0.057 | 19,73±1.09 | ||
| Сем. Agamidae | 0,909±0.062 | 22.48±1.53 | ||
| Сем. Helodermatidae | 0.968 | 23.95 |
| Сем. Varanidae | 1,003±0.068 | 24.82±1.68 | ||
| Сем. Lacertidae | 1,462±0.130 | 25.11±1.84 | ||
| Отряд Crocodylia | 0,880±0.102 | 21.76±2.52 | ||
| Сем. Alligatoridae | 0.863±0.104 | 21.34±2.58 | ||
| Сем. Crocodylidae | 0.947±0.322 | 23.42±7.96 |
Имеются, конечно, значительные трудности при сопоставлении критерия упорядоченности и палеонтологических данных о времени появления того или иного таксона. Эти трудности касаются, как вычисления критерия упорядоченности, так и точности определения времени происхождения изучаемого таксона. И все же выявляется достаточно четкая корреляция между величиной критерия упорядоченности и временем возникновения разных отрядов пресмыкающихся в процессе эволюции. Согласно палеонтологическим данным черепахи возникли в пермский период, чешуйчатые - в поздней перми - раннем триасе, крокодилы - в триасе (Дуршиц, Обручева, 1971). Как видно из табл.36 критерий упорядоченности для отряда черепах Сr=15.26, для отряда чешуйчатых Сr=18,35 и для отряда крокодилов Сr=21.76. Наименьшую величину критерия упорядоченности имеет отряд Rhynchocephalia (Сr=12.62), но учитывая, что нам известен стандартный обмен только одного вида этого отряда (Wilson, Lee, 1970), а также то, что этот отряд относится к подклассу Lepidosauria (т.е. в отличие от черепах, к тому же подклассу, что и чешуйчатые и крокодилы) в макросистематике класса рептилий отряды по критерию упорядоченности следует располагать в следующем порядке: