Gt; Chiroptera —> Primates —> Cetacea —> Perissodactyla
Последовательность расположения отрядов по этой схеме отличается не только от предложенной другими авторами (табл.38), но и макросистемы отрядов млекопитающих, описанной в работе Зотина и Владимировой (1991), хотя она основана примерно на тех же принципах, что и представленная в данной работе. Это связано в основном с тем, что в указанной работе не учтены климатические пояса, в которых живут изученные виды млекопитающих.
Конечно, представленное в табл.38 и на схеме расположение отрядов это только предварительная макросистема инфракласса Eutheria класса млекопитающих, так как для некоторых отрядов еще недостаточно число данных о стандартном обмене и о температуре тела (Sirenia, Pholidota, Tylopoda). Для других отрядов довольно сложно толковать полученные данные (Insectivora - положение отряда основано только на Сr для семейств Tenrecidae, Erinaceidae, Macroscelididae) и, наконец, для третьих отрядов вообще отсутствуют данные о стандартном обмене (Proboscidea)
В целом для млекопитающих коэффициент а и критерий упорядоченности Сr равны:
Класс Mammaliaa=24.29±0.09; Cr=104.0±0.2 (125) Отряд Insectivora a=39.50±0.50; Cr=155.8±0.96
Τаблица 38. Расположение отрядов в макросистематике млекопитающих по разным авторам.
| По Соколову (1984) | 1* | ||||||
| (1) Monotremata | |||||||
| (2) Marsupialia | |||||||
| (3) Edentata | |||||||
| (4) Insectivora | |||||||
| (5) Dermoptera | |||||||
| (6) Chiroptera | |||||||
| (7) Primates | |||||||
| (8) Carnivora |
| (9) Pinnipedia | |||||||
| (10) Cetacea | |||||||
| (11) Sirenia | |||||||
| (12) Proboscidea | |||||||
| (13) Perissodactyla | |||||||
| (14) Hyracoidea | |||||||
| (15) Tubulidentata | |||||||
| (16) Artiodactyla | |||||||
| (17) Tylopoda | |||||||
| (18) Pholidota | |||||||
| (19) Rodentia | |||||||
| (20) Lagomorpha |
Авторы: (1) Соколов, 1973, 1978, 1979; (2) Наумов, 1965; (3) Гептнер и др., 1961; (4) Матвеев, 1949; (5) Кашкаров, Станчинский, 1940; (6) Брэм, 1930; (7) Холодковский, 1918.
Класс Aves (Птицы)
В некоторых работах приведено довольно много данных о стандартном обмене птиц, но в них присутствует явная систематическая ошибка, так как другие авторы получили у сходных видов птиц заметно иные результаты. Это относится к работам Шилова (1968), Yarbrough (1971) и Hails (1983). Следует отметить, что ошибка в этих работах носила систематический характер. Для того, чтобы использовать полученные ими данные мы внесли поправку на основании работ других авторов.
Для птиц получено большое число данных по стандартному обмену (обмену покоя в термонейтральной зоне) и его зависимости от массы тела (King, Farner, 1961; Lasievski, Dawson, 1967; Шилов, 1968; Kendeigh et al., 1977; Зотин, 1979, 1988; Calder, 1981, 1984; Gavrilov, Dolnik, 1985). В этом классе позвоночных наблюдаются заметные различия в коэффициентах, входящих в аллометрическое соотношение (46), у разных отрядов и семейств (Zar, 1968; Зотин, 1979). Особенно велики различия в стандартном обмене отряда воробьинообразных и остальными отрядами птиц. Это различие настолько велико, что нельзя получить единую зависимость стандартного обмена от массы
тела для всего класса птиц (рис.27). Из рис.27 следует, что стандартный обмен представителей отряда воробьиных во всем диапазоне масс заметно выше, чем остальных птиц.
Как и в случае млекопитающих, разными авторами получены довольно близкие величины коэффициентов в аллометрической зависимости (46) для птиц: для 60 видов а=21.6, k=0.74 (King, Farner, 1961); для 48 видов воробьиных а=43.1, &=0.724 и 72 видов неворобьиных а=26.2, k=0.723 (Lasiewski, Dawson, 1967); для 99 видов воробьиных а=40.7, k=0.74 и 85 видов неворобьиных а=25.1, k=0.74 (Зотин, 1979); для 138 (п=242) видов воробьиных и 90 (п=151) неворобьиных птиц по данным Гаврилова и Дольник (Gavrilov, Dol'nik, 1985) соответственно α=34.5, k=0.15 и α=26.2, k=0.73 (Зотин, 1988). Использование собранных нами данных о стандартном обмене птиц [159 видов воробьиных (n=247) и 180 видов неворобьиных (n=254)] и применение статистически правильного метода определения коэффициентов k в аллометрических соотношениях (Зотин, 1999) показало, что аллометрические соотношения для птиц имеют вид:
воробьинообразные :

|
| где k=0.720±0.182; остальные птицы (без представителей отряда колибриобразных): |

|
| где k=0.712±0.070; колибриобразные: (126) |
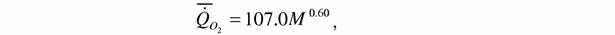
|
| где k=0.599±0.192. |
Полученные значения коэффициента k достоверно не отличаются от константы Хемингсена. Таким образом, мы вправе использовать для вычисления коэффициента а и а' стандартный для всех птиц коэффициент k=0.75.
При расчетах Cr y птиц, подобно млекопитающим, следует учитывать, что они являются гомойотермными животными. Температура тела птиц даже в
термонейтральной зоне равна примерно 40-41°С, это намного выше, чем 20°С, при которой определяется стандартный обмен у пойкилотермных животных. Поэтому, чтобы сравнить критерий упорядоченности тех и других необходимо, как уже говорилось в отношении млекопитающих, привести стандартный обмен по кривой Крога (69) к 20°С. Мы сделали это, учитывая средние данные о температуре тела представителей отдельных отрядов птиц (Dawson, Hudson, 1970; Гаврилов, Зотин, 1998).
Большую трудность при вычислении критерия упорядоченности представляет учет климатической зоны, в которой живут те или иные виды птиц. Дело в том, что многие из них являются перелетными, т.е. гнездуются они в северном полушарии, а на зиму мигрируют в теплые страны. В подобных случаях мы использовали коэффициент ау/аT равный не 1.66, а 1.33.
Данные о стандартном обмене и критерии упорядоченности для отдельных отрядов птиц приведены в табл.39. Как видно из этой таблицы колибриобразные имеют самый высокий критерий упорядоченности.
Согласно Карташеву (1974), в классе птиц имеется 33 отряда. В нашем обзоре (Гаврилов, Зотин, 1998) собраны данные о массе и интенсивности дыхания представителей 26 отрядов птиц (табл.39). Для некоторых отрядов известны данные о дыхании 2-3 видов птиц, поэтому их положение в системе отрядов птиц, расположенных в порядке возрастания критерия упорядоченности (табл.39), достаточно условно. В отношении остальных отрядов, представленных в табл.39, кроме воробьинообразных, также пока невозможно гарантировать правильность их позиции. И дело не только в том, что для многих отрядов получено недостаточное число данных о дыхании, но и в особенностях экологии разных птиц, которая может вносить заметную ошибку в определение уровня стандартного обмена, особенно когда он мало отличается в разных отрядах. Известно, например, что колибри в состоянии покоя впадают в оцепенение, характеризующееся очень низким уровнем интенсивности дыхания (Pearson, 1953). Поэтому для этих птиц стандартный обмен следует изучать не в состояние покоя, как для других животных, а перед началом полета или сразу после окончания полета, когда колибри выходят из состояния оцепенения или еще не успели в него попасть. Но это в свою очередь приводит к тому, что у них определяют не стандартный, а активный обмен. Этим можно объяснить
чрезвычайно высокую величину коэффициента а и критерия упорядоченности для колибриобразных, показанную в табл.39.
Несмотря на сказанное, нам кажется правильным строить макросистематику по отрядам в классе птиц в соответствии с критерием упорядоченности Сr, который, во-первых, является реально измеримой величиной, и, во-вторых, отражает последовательность возникновения отрядов в процессе эволюции.
Сложнее дело обстоит с попыткой построения макросистематики птиц на уровне семейств. К настоящему времени, пожалуй, можно наметить такую макросистему только для отряда воробьинообразных, так как для этого отряда имеется достаточно большое число измерений дыхания.
В этом отряде 72 семейства (Караташев, 1974). В нашей работе (Гаврилов, Зотин, 1998) собраны данные об интенсивности дыхания и массе тела только 26 семейств. В табл.40 приводятся средние величины коэффициента а в термонейтральной зоне и критерий упорядоченности Сr для этих семейств. При расчете критерия упорядоченности принималось, что для всех видов воробьинообразных температура тела равна 40.6°С. Для перелетных птиц, зимующих в теплых странах, принято ау/аT=1.33.
Таблица 39
Коэффициент а, мВт/г (по данным о стандартном обмене в термонейтральной зоне), средняя температура тела t°C и критерий упорядоченности Сr (приведенный к температуре 20°С), в разных таксонах класса Aves (Птицы). N - число изученных видов, n - число измерений.
| Таксоны | а | t°C | Сг | N | n |
| Надотряд Impennes | 22.34±0.80 | 70.75±2.52 | |||
| Отряд Sphenisciformes | 22.34±0.80 | 38.2 | 70.75±2.52 | ||
| Надотряд Neognathae | 32.77±0.59 | 112.35±1.26 | |||
| Отряд Apterygiformes | 11.88 | 38.4* | 60.28 | ||
| Отряд Caprimulgiformes | 15,91±1.71 | 40.0* | 63.12±8.18 | ||
| Отряд Gruiformes | 21.03 | 40.9 | 64.75 | ||
| Отряд Ralliformes | 22.32 | 40.5 | 68.99 |
| Отряд Anseriformes | 24.25±1.15 | 41.0* | 71.37±2.37 | ||
| Отряд Cuculiformes | 21.07 | 42.1* | 71.68 | ||
| Отряд Columbiformes | 21.93±1.02 | 40.6 | 72.73±3.83 | ||
| Отряд Falconiformes | 21.90±l.46 | 40.2 | 76.9±5.53 | ||
| Отряд Casuariiformes | 16.76 | 39.8* | 76.91 | ||
| Отряд Strigiformes | 18.46±1.06 | 37.9 | 77.25±5.23 | ||
| Отряд Ciconiiformes | 23.52±0.89 | 40.9* | 82.07 | ||
| Отряд Galliformes | 22.75±1.01 | 40.1 | 86.36±2.02 | ||
| Отряд Piciformes | 27,78 | 41.0* | 92.25 | ||
| Отряд Procellariiformes | 29.07±2.97 | 39.2 | 92.78±5.05 | ||
| Отряд Trogoniformes | 21,83 | 40.5 | 94.40 | ||
| Отряд Lariformes | 30.43±43 | 40.3 | 95.62±5.96 | ||
| Отряд Coraciiformes | 26,59 | 40.0* | 96.12 | ||
| Отряд Charadriiformes | 29.33±1.40 | 40.4* | 101.86±4.75 | ||
| Отряд Psittaciformes | 26.57±1.35 | 41.1 | 109.19±5.56 | ||
| Отряд Rheiformes | 23.65 | 39.5 | 113.99 | ||
| Отряд Struthioniformes | 24.25 | 39.5 | 114.19 | ||
| Отряд Passeriformes | 37,24±0.47 | 40.6 | 116.35±1.41 | ||
| Отряд Alciformes | 43.3±3.15 | 40.1 | 116.71±8.49 | ||
| Отряд Pelecaniformes | 28.29 | 40.6 | 121.33 | ||
| Отряд Apodiformes | 26,52±1.10 | 40.1* | 126.91±7.98 | ||
| Отряд Coliif ormes** | 59,74±5.01 | 286.08±22.1 |
* Из Dawson, Hudson, 1970; ** В основой по данным Krüger et al., 1982
Таблица 40
Коэффициент а, мВт/г и критерий упорядоченности Сr в разных семействах отряда Passeriformes(Воробьинообразные). N - число изученных видов, n - число измерений.
| Таксон | а | Сr | N | n |
| Подотряд Passeres | 37.29±0.47 | 115.63±1.37 | ||
| Сем. Paradoxornithidae | 38.38 | 99.15 |
| Сем. Bombycillidae | 38.52 | 99.50 | ||
| Сем. Parulidae | 29.22±0.58 | 100.39±2.00 | ||
| Сем. Alaudidae | 32.20 | 103.48 | ||
| Сем. Cinclidae | 24.38 | 104.55 | ||
| Сем. Fringillidae | 38.02±0.63 | 105.63±1.65 | ||
| Подсем. Pyrrhuloxiinae | 35.28±1.33 | 98.49±1.56 | ||
| Подсем. Emberizinae | 35.20±l.03 | 98.67±2.71 | ||
| Подсем. Fringillinae | 40.52±0.74 | 111.88±2.02 | ||
| Сем. Corvidae | 32.63±2.30 | 107.83±6.18 | ||
| Сем. Oriolidae | 32.84 | 112.84 | ||
| Сем. Laniidae | 32.94 | 113.15 | ||
| Сем. Sylviidae | 35.82±0.88 | 120.72±2.39 | ||
| Сем. Muscicapidae | 35.25 | 121.10 | ||
| Сем. Motacillidae | 35.26±0.83 | 121.14±2.87 | ||
| Сем. Sittidae | 40.71 | 121.31 | ||
| Сем. Paridae | 44.23±1.91 | 124.79±6.61 | ||
| Сем. Icteridae | 36.63±2.75 | 125.85±9.44 | ||
| Сем. Hirundinidae | 36.90±1.98 | 126.78±6.81 | ||
| Сем. Ploceidae | 38.63± | 129.14±3.27 | ||
| Сем. Turdidae | 37.67±1.35 | 129.43±4.63 | ||
| Сем. Regulidae | 51.30 | 132.52 | ||
| Сем. Sturnidae | 43.54 | 135.17 | ||
| Сем. Prunellidae | 46.53 | 136.86 | ||
| Сем. Troglodytidae | 47.00 | 138.90 | ||
| Сем. Drepanididae | 37.18 | 159.41 | ||
| Подотряд Tyranni | 35.05±0.48 | 145.01±2.11 | ||
| Сем. Tyrannidae | 37.33 | 128.24 | ||
| Сем. 9 | 31.06 | 133.18 | ||
| Сем. Pipridae | 48.76 | 209.11 |
Подотряды, семейства и подсемейства воробьинообразных в табл.40 расположены в порядке возрастания критерия упорядоченности. Здесь также нет
полной уверенности в правильности расположения этих подразделений. В частности, семейство американских славковых (сем. Harulidae), по-видимому, незаслуженно имеет довольно низкий показатель. Дело в том, что о дыхании представителей этого семейства мы можем судить только по работе Ярбро (Yarbrough, 1971), а все его данные о стандартном обмене явно занижены. Следовательно, для правильного расположения отдельных семейств в отряде воробьинообразных, так же, как отдельных отрядов в классе птиц, в соответствии с критерием упорядоченности требуется еще значительная работа по накоплению данных о стандартном обмене разных видов птиц. Следует только отметить, что расположение семейств в отряде воробьинообразных (табл.40), так же, как отрядов в классе птиц (табл.39), в порядке увеличения критерия упорядоченности не совпадает с расположением, приводимым в книге Карташева (1974).
Возвращаясь к стандартному обмену и критерию упорядоченности для всех птиц, опираясь на данные, приводимые в табл.39, напишем:
Класс Avesa=32.64±0.58; Cr=l 11.9+2.4
Класс Aves(без воробинных) а=28.20±0.99; Сr=107.5±4.6 (127)
Отряд Passeriformes a=37.24±0.47; Сr=116.4±1.4
Отряд Coliiformes а=59,74±5.01 ; Cr=286.1±22.l
Подводя итоги для всего типа позвоночных, получим в среднем по данным, приводимых в формулах (119)-(127):
Тип Vertebrata a=6.93; Сr=33.61 (128)