Направление и скорость прогрессивной эволюции и биоэнергетического прогресса
Хотя критерий эволюционного прогресса, использованный Ивлевым и Дольником, пока не получил широкого признания (особенно среди профессиональных эволюционистов), нет сомнений, что он может быть положен в основу количественной характеристики эволюционного прогресса, включая расчет скорости биоэнергетического прогресса и прогрессивной эволюции.
Исходя из данных, приводимых в табл..41 и рис.28, можно считать, что биоэнергетический прогресс и прогрессивная эволюция в животном царстве происходили в следующем направлении:
Простейшие —> Гребневики —> Кишечнополостные —> —> Черви —> Иглокожие —> Членистоногие —> Моллюски —> —> Оболочники —> Полухордовые —> Позвоночные,
в типе моллюсков:
Панцирные —> Двустворчатые —> Брюхоногие —> Головоногие, (130)
в типе членистоногих:
Многоножки —> Паукообразные —> Ракообразные —> Насекомые (131)
в типе позвоночных:
Ланцетниковые —> Круглоротые —> Хрящевые рыбы —>
—> Костные рыбы —> Земноводные —> Пресмыкающиеся —> (132)
—> Млекопитающие —> Птицы
Значимость этих утверждений становится еще более очевидной, если мы сопоставим величину коэффициента а каждой из изученных групп животных со временем обнаружения представителей этих групп в палеонтологической летописи (Зотин, 1979, 1984; Зотин, Криволуцкий, 1982; Leuschner, 1989; Zotin, 1990).
Скорость эволюционного прогресса в животном царстве. Как видно из табл.41, 42 и рис.28, в процессе эволюции происходило последовательное появление новых типов животных с более высоким уровнем энергетического обмена и критерия упорядоченности. Эти типы животных, как говорилось, по всем признакам обладали более сложным строением, совершенным функционированием и поведением. Мы можем, поэтому, судить по кривой, приводимой на рис.28, о скорости биоэнергетического прогресса и скорости прогрессивной эволюции в целом. Эти скорости можно выразить конкретными величинами с размерностью мВт/г· миллион лет.
Прежде чем делать это, следовало бы решить еще один вопрос. Речь идет о количественном описании кривых, показанных на рис.28. Указанные кривые могут быть аппроксимированы различными функциями: степенной,
экспонентой, параболой, гиперболой, функцией Гомперца и т.д. Однако, по нашему мнению, для их аппроксимации лучше всего подходит экспонента (Зотин, Зотин, 1995). Этот вывод имеет хороший теоретический фундамент в виде принципа наискорейшего спуска, согласно которому уклонение подсистемы от стационарного состояния происходит приблизительно по экспоненте (45). Расчет по данным, приводимым в табл.41 и 42, показывает, что биоэнергетический прогресс и прогрессивная эволюция на уровне типов животных в процессе эволюции описывается экспонентой с константами
 (136)
(136)
Здесь коэффициент а выражен в мВт/г, а геологическое время tg - в миллионах
лет.
Различие степенных коэффициентов в формулах (136), по-видимому, связано с появлением гомойотермии в типе позвоночных (в случае критерия упорядоченности учитывалась температура тела гомойотермных животных). Если рассчитывать биоэнергетический прогресс только для беспозвоночных, то степенной коэффициент оказывается таким же, как и в случае прогрессивной эволюции:
 (137)
(137)
Формулы (136) и (137) не только показывает общую тенденцию эволюционного прогресса, но и позволяют оценить скорость этого процесса для любого отрезка эволюции по производной da/dt и скорость прогрессивной эволюции по производной dCr/dt, согласно формуле (136).
По кривой, показанной на рис.6, ранее уже предпринималась попытка определить скорость биоэнергетического прогресса (но не прогрессивной эволюции, так как авторы не различали этих понятий) в каждый из геологических периодов Земли (Зотин, Криволуцкий, 1982; Зотин, 1984; Zotin, 1990). Теперь мы можем уточнить эти величины, опираясь на кривую рис.28 и формулу (136), как и скорости прогрессивной эволюции, опираясь на кривую рис.28 и формулу (136), по производным da/dt и dCr/dt для каждого
геологического периода (табл.43). Расчет показывает, что скорость изменения стандартного обмена, выраженная коэффициентом а, т.е. скорость биоэнергетического прогресса животных в разные геологические периоды, возрастала по мере приближения к современному времени.
Конечно, цифры, показывающие скорость и увеличение скорости эволюционного прогресса (табл.43), будут уточняться по мере накопления экспериментальных данных о стандартном обмене и палеонтологических наблюдений о времени возникновения отдельных типов животных (это видно хотя бы на примере наших расчетов, проведенных в разные годы - Зотин, 1984; Zotin, 1990; Зотин, Зотин, 1995), но общая тенденция биоэнергетического прогресса и прогрессивной эволюции в целом ясна: скорость того и другого возрастает в процессе биологической эволюции.
Таблица 43
Протяженность геологического времени tg, (млн. лет), скорость биоэнергетического прогресса da/dtg (мВт/г млн. лет) и прогрессивной эволюции dCr/dtg (млн. лет-1) животных в разные геологические периоды.
| Геологический период | tg | 
| 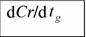
|
| Кембрий | 0.0013 | 0.019 | |
| Ордовик | 0.0020 | 0.027 | |
| Силур | 0.0028 | 0.036 | |
| Девон | 0.0039 | 0.046 | |
| Карбон | 0.0057 | 0.062 | |
| Пермь | 0.0090 | 0.088 | |
| Триас | 0.0119 | 0.110 | |
| Юра | 0.0168 | 0.145 | |
| Мел | 0.0265 | 0.207 | |
| Третичный период | 0.0418 | 0.296 | |
| Четвертичный период | 0.0529 | 0.356 |
Прогресс в отдельных типах животных. По мере того, как будут накапливаться экспериментальные данные о стандартном обмене разных видов
животных, можно будет рассмотреть вопрос и о скорости эволюционного прогресса внутри отдельных крупных таксонов порядка типа или класса. Основная трудность, как и в случае построения общей кривой биоэнергетического прогресса и прогрессивной эволюции для всего животного царства (рис.28), заключается в точном установлении времени возникновения того или иного класса или отряда животных. Данные по этому поводу постоянно меняются. И все же в настоящее время мы можем рассчитывать скорость прогрессивной эволюции внутри отдельных типов животных (моллюски, членистоногие, позвоночные) с некоторой долей уверенности. Это очень важно для проблем, рассматриваемых в данной книге, так как принципиальным является вопрос о том, имеется ли лишь одна единая кривая биоэнергетического прогресса для всего животного царства или в каждом из типов или даже классов животных скорость эволюционного прогресса является величиной, характерной для данного таксона или для времени его возникновения.
Согласно данным о критерии упорядоченности схема расположения классов в типе моллюсков имеет вид (130), в типе членистоногих - (131) и в типе позвоночных - (132). В соответствии с этими схемами мы можем построить кривые прогрессивной эволюции в типах моллюсков, членистоногих и позвоночных (рис.29). В принципе они имеют сходный характер и имеют вид экспоненты. Но реальная скорость изменения критерия упорядоченности, т.е. скорости прогрессивной эволюции, отличается в разных типах животных (рис.29). Общая тенденция прогрессивной эволюции - она ускоряется по ходу биологической эволюции. Изменения коэффициента а и критерия упорядоченности Сr при биоэнергетическом прогрессе и прогрессивной эволюции в типе моллюсков могут быть записаны в виде:
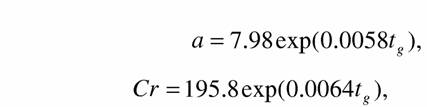
| (138) |
| в типе членистоногих: | |
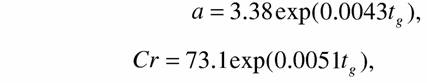
| (139) |
в типе позвоночных:
a = 7.91exp(0.0059tg), (140)
Cr = 64.5exp(0.0063tg).
В принципе эти формулы могут быть использованы для сравнения скорости биоэнергетического прогресса и скорости прогрессивной эволюции в указанных типах животных по коэффициенту β из уравнения (45): чем больше этот коэффициент, тем больше скорость эволюционного прогресса. Биоэнергетический прогресс и прогрессивная эволюция в разных типах животных, как видно из формул (138)-(140), шли приблизительно с одинаковой скоростью.
Прогресс в отдельных классах животных. Можно продолжить уточнение кривых прогрессивной эволюции и распространить их на отдельные классы животных. На рис.30 показаны кривые изменения критерия упорядоченности разных отрядов в классе насекомых, моллюсков и млекопитающих. Судя по этим данным, в отдельных классах животных прогрессивная эволюция шла с разной скоростью. Расчет уравнения биоэнергетического прогресса и прогрессивной эволюции в классе насекомых, по данным, приводимым в табл.41 и 42, показал, что они могут быть записаны в виде:
a = 2.36exp(0.0016•tg) (141)
Cr = 47.5exp(0.0022•tg )
в классе костных рыб:
a = 1.77exp(0.0024•tg) (142)
Cr = 32.0exp(0.0019•tg)
в классе млекопитающих:
a= 34.8exp(0.0108•tg) (143)
Cr = 114.8exp(0.0030•tg )
По формулам (141)-(143) с помощью степенного коэффициента можно судить о скоростях биоэнергетического прогресса и прогрессивной эволюции в классе насекомых, костных рыб и млекопитающих, а также сравнить эти скорости в разных классах. Мы видим, что биоэнергетический прогресс и прогрессивная эволюция в классе насекомых и костных рыб шли приблизительно с одинаковой скоростью, а в классе млекопитающих прогрессивная эволюция шла медленее, чем биоэнергетический прогресс. Если сравнивать скорость биоэнергетического прогресса в разных классах животных, то она возрастала, судя по формулам (141-143), в последовательности: насекомые —> костные рыбы —> млекопитающие, - а скорость прогрессивной эволюции - в последовательности: костные рыбы —> насекомые —> млекопитающие.
Приводимые сопоставления скоростей биоэнергетического прогресса и прогрессивной эволюции на уровне типов и классов лишь приблизительны. Они будут уточняться по мере получения новых данных о стандартном обмене животных и времени возникновения таксонов в процессе реальной эволюции. Но они показывают, что количественное описание этих процессов в реальном масштабе времени в принципе возможно.
Таковы факты, показывающие направленность эволюционного прогресса. Конечно, требуется еще огромная работа по уточнению кривых эволюционного прогресса (рис.28-30), и констант, входящих в соотношение (136-143). Но такая работа была бы вполне оправдана, учитывая важность сделанных выводов для многих проблем эволюционного ученья, систематики и палеонтологии. Особенно важным, с нашей точки зрения, является следствие, вытекающее из данных, приводимых в табл.41 и на рис.28, согласно которому макроэволюция на уровне крупных таксонов в указанные геологические периоды шла с усилением энергетического обмена.
Опираясь на эти выводы, кажется разумным строить макросистематику животных любых крупных таксонов, опираясь на данные о стандартном обмене и критерии упорядоченности. Преимуществом такого подхода является то, что
он, с одной стороны отражает эволюцию в реальном масштабе времени, с другой - показывает направление прогрессивной эволюции в том или ином таксоне.
Основной вывод, который можно сделать из изложенного выше состоит в том, что в процессе макроэволюции происходило усиление энергетического обмена. Это не единственное, но, по нашему мнению, главное содержание прогрессивной эволюции, во многом определяющее и все остальные направления эволюционного прогресса и, в частности, увеличение организованности живых организмов в процессе прогрессивной эволюции. В дальнейшем мы остановимся на последнем более подробно, но и здесь следует отметить, что увеличение сложности строения и функционирования животных происходит на базе повышения энергетического обмена.