Критерии организованности
Негэнтропийные эффекты. Прежде всего, можно говорить, в принципе, об использовании для эволюционных проблем понятия негэнтропийного эффекта 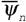 - чем больше величина
- чем больше величина 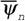 , тем лучше организована живая система. Можно, по-видимому, использовать для характеристики организованности живых систем и диссипативную функцию
, тем лучше организована живая система. Можно, по-видимому, использовать для характеристики организованности живых систем и диссипативную функцию 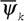 (50) в том смысле, что при прочих равных условиях, чем меньше тратится энергии на управление, тем совершеннее организованность системы.
(50) в том смысле, что при прочих равных условиях, чем меньше тратится энергии на управление, тем совершеннее организованность системы.
В живых системах, которые можно определить, как открытые, далекие от равновесия, организованные системы, способные к саморазмножению и саморазвитию, всегда имеют место негэнтропийные эффекты. Они заключаются в том, что в соответствии с равенством (50) в устойчивом состоянии такой системы функция внешней диссипации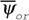 меньше, чем в точно такой же по физическому и химическому устройству, но не организованной системе
меньше, чем в точно такой же по физическому и химическому устройству, но не организованной системе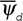 (рис. 1). Это определяется тем, что организованность клеток, зародышей и целых организмов возникает в результате работы целой системы управления, реализующей записанную в геноме программу. Поэтому в них всегда имеют место негэнтропийные эффекты: локальный негэнтропийный эффект биохимической регуляции, связанный с наличием в клетках аллостерических ферментов, активность которых регулируется конечными или промежуточными продуктами реакций, и сложной сети взаимодействующих биохимических процессов; интегральный негэнтропийный эффект биохимической регуляции, выражающийся в компартментализации биохимических процессов, мембранном контроле метаболизма, переходом ферментов из активной формы в неактивную и т.д.; гормональный негэнтропийный эффект и негэнтропийный эффект нервной регуляции, необходимые для поддержания строго определенного уровня теплопродукции и дыхания организма (Зотин, 1988).
(рис. 1). Это определяется тем, что организованность клеток, зародышей и целых организмов возникает в результате работы целой системы управления, реализующей записанную в геноме программу. Поэтому в них всегда имеют место негэнтропийные эффекты: локальный негэнтропийный эффект биохимической регуляции, связанный с наличием в клетках аллостерических ферментов, активность которых регулируется конечными или промежуточными продуктами реакций, и сложной сети взаимодействующих биохимических процессов; интегральный негэнтропийный эффект биохимической регуляции, выражающийся в компартментализации биохимических процессов, мембранном контроле метаболизма, переходом ферментов из активной формы в неактивную и т.д.; гормональный негэнтропийный эффект и негэнтропийный эффект нервной регуляции, необходимые для поддержания строго определенного уровня теплопродукции и дыхания организма (Зотин, 1988).
Кроме того, в живых системах протекает множество эндотермических, сопряженных процессов, что также связано с наличием негэнтропийных эффектов.
Из указанных негэнтропийных эффектов реально измеримым в настоящее время является только интегральный негэнтропийный эффект. Измерить его можно, сопоставляя теплопродукцию или дыхание целых организмов и их гомогенатов, в которых тем или иным способом увеличена до максимума проницаемость наружных мембран митохондрий. В таких гомогенатах все интегрирующие связи и компартменты разрушены и негэнтропийный эффект проявляется в значительном увеличении энергетического обмена. К сожалению, до сих пор не проведено опытов, где бы помимо гомогенизации и добавления избытка субстратов дыхания проводили бы увеличение проницаемости митохондриальных мембран, что позволило бы определить реальный, а не частичный интегральный негэнтропийный эффект. Однако само наличие интегрального негэнтропийного эффекта у ооцитов и зародышей не вызывает сомнений (Зотин, Зотина, 1993).
Управление движением. Универсальные критерии организованности, по-видимому, могут быть получены в результате изучении энергетических затрат животных на управление, т.е. с помощью измерения диссипативной функции
 (50) при протекании различных процессов и, в частности, при осуществлении движения. Эта проблема достаточно сложна, так как способы перемещения животных очень разнообразны (ходьба и бег на двух и четырех конечностях, лазанье, прыжки, ползанье, разные типы плавания и полета), а сравнение степени совершенства управления можно проводить только при одинаковом типе движения (Зотин, 1988). Кроме того, необходимо изучать энергетику движений в строго сопоставимых условиях: бег на тредбане, плавание в гидродинамической или полет в аэродинамической трубах. Сравнивать между собой энергетическую стоимость движения животных в разных средах достаточно трудно, а, главное, особенности движения животных характеризуют скорее экологические особенности их жизни, чем положение на шкале прогресса.
(50) при протекании различных процессов и, в частности, при осуществлении движения. Эта проблема достаточно сложна, так как способы перемещения животных очень разнообразны (ходьба и бег на двух и четырех конечностях, лазанье, прыжки, ползанье, разные типы плавания и полета), а сравнение степени совершенства управления можно проводить только при одинаковом типе движения (Зотин, 1988). Кроме того, необходимо изучать энергетику движений в строго сопоставимых условиях: бег на тредбане, плавание в гидродинамической или полет в аэродинамической трубах. Сравнивать между собой энергетическую стоимость движения животных в разных средах достаточно трудно, а, главное, особенности движения животных характеризуют скорее экологические особенности их жизни, чем положение на шкале прогресса.
Высшая нервная деятельность. Безусловно полезным было бы разработать критерии для характеристики высшей нервной деятельности животных. Такие попытки не раз предпринимались (Крушинский, 1977), но большинство предложенных критериев сильно зависели от условий проведения опытов, т.е. мало пригодны для сравнения совершенно различных животных. В
настоящее время успешно можно использовать для целей сравнения только такой косвенный показатель высшей нервной деятельности животных, как относительные размеры мозга, который, как считают многие авторы (Никитинко, 1965, 1969; Крушинский, 1977; Крушинская, Лисицына, 1983), коррелирует с уровнем высшей нервной деятельности животных.
Гомеостаз. В живых системах поддержание устойчивого состояния или возвращение в устойчивое состояние после его нарушения связано с процессом управления и регуляции, который получил название гомеостаз, а поддержание устойчивости развития и роста организмов - гомеорез.
Существуют различные определения понятия гомеостаза (Cannon, 1929, 1932;Розен, 1969;Новосельцев, 1978). Мы придерживаемся следующего (Зотин, Зотина, 1987, 1993; Зотин, 1988): гомеостаз - процесс поддержания устойчивого состояния организованных систем.
С точки зрения этого определения механизмы гомеостаза направлены на поддержание минимальной величины функции внешней диссипации (19), (51). В биологических системах, как и в любых других, функцию внешней диссипации 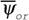 (50) можно приравнять интенсивности теплопродукции
(50) можно приравнять интенсивности теплопродукции  (14) или дыхания
(14) или дыхания 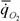 (65), т.е. гомеостаз состоит в поддержании на постоянном уровне основного (стандартного) обмена. Все остальные параметры организма также поддерживаются на постоянном уровне, так как они связаны с диссипативной функцией (8), а, следовательно, и с функцией внешней диссипации.
(65), т.е. гомеостаз состоит в поддержании на постоянном уровне основного (стандартного) обмена. Все остальные параметры организма также поддерживаются на постоянном уровне, так как они связаны с диссипативной функцией (8), а, следовательно, и с функцией внешней диссипации.
Гомеостаз, благодаря целой системе регуляций и управлений, создает некоторую независимость организмов от изменений, происходящих в среде. Совершенство этой системы регуляции показывает степень продвинутости организмов по пути прогрессивной эволюции (Медников, 1973; Грант, 1980) и степень их организованности. Другими словами, надежность и устойчивость гомеостаза можно было бы использовать для характеристики степени организованности живых систем. Необходима поэтому разработка количественных критериев гомеостаза, что и было частично сделано в предшествующих работах (Зотин, Зотина, 1987, 1993; Зотин, 1988).
Первый критерий, который был назван мерой надежности гомеостаза, опирается на следующие рассуждения. Если гомеостаз надежен, то в ответ на
изменение факторов внешней среды организованная система мало отклоняется от устойчивого состояния, и чем надежнее гомеостаз, тем меньше это отклонение. В термодинамике неравновесных процессов мерой отклонения системы от устойчивого, стационарного состояния является увеличение функции внешней диссипации. Поэтому, чем надежнее организована система, чем надежнее ее гомеостаз, тем в меньшей степени возрастает функция внешней диссипации при изменении того или иного параметра окружающей среды. Вслед за Хаскиным (1975) мы назвали величину отклонения 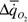 при изменении параметра среды Δx энергетической ценой гомеостаза. Если выразить
при изменении параметра среды Δx энергетической ценой гомеостаза. Если выразить 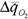 в % от минимальной величины
в % от минимальной величины 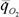 , энергетическая цена гомеостаза η% равна:
, энергетическая цена гомеостаза η% равна:
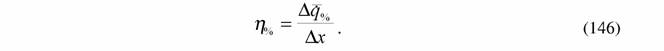
|
| Для характеристики надежности удобнее пользоваться величиной, пропорциональной надежности. Поэтому в качестве меры надежности гомеостаза (М) лучше принять величину, обратно пропорциональную энергетической цене гомеостаза: |

|
| Вычисление η% по формуле (146) несколько осложняется тем обстоятельством, что интенсивность теплопродукции или дыхания может зависеть от изменения параметров внешней среды нелинейным образом. Мы приняли (Зотин, 1988; Зотин, Зотина, 1993), что эта зависимость имеет вида параболы (рис.1): |
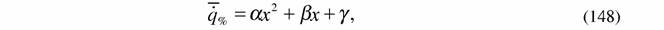
|
где α , β , γ - коэффициенты.
Если учесть, что при оптимальном значении параметра x 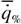 = 1 , то формулу (148) можно преобразовать следующим образом: = 1 , то формулу (148) можно преобразовать следующим образом:
|
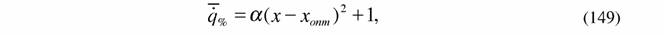
|
где хопт = -β/2α - значение параметра в точке оптимума при 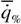 = 1.
= 1.
Энергетическая цена гомеостаза (146) соответствует первой производной 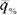 по параметру х:
по параметру х:
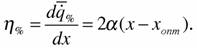
Из этой формулы следует, что энергетическая цена линейно зависит от параметра внешней среды х. Это вполне естественно, но не удобно для определения энергетической стоимости гомеостаза, так как в этом случае следует условиться о какой точке кривой (149) идет речь и определять η% только в этой точке. От этого недостатка свободна вторая производная 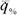 по параметру х, которую и можно принять в качестве энергетической цены гомеостаза:
по параметру х, которую и можно принять в качестве энергетической цены гомеостаза:
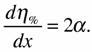
В качестве меры надежности в этом будет выступать величина:
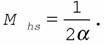 (150)
(150)
Как видно из (150), мера надежности обратно пропорциональна энергетической цене гомеостаза. Следовательно, как и энергетическая цена, мера надежности характеризует эффективность и экономичность управляющих и регулирующих систем гомеостаза.
Не менее важным параметром, характеризующим надежность гомеостаза является ширина зоны изменений параметров внешней среды, в пределах которой функция внешней диссипации сохраняет величину, близкую к минимальной. Мы назвали эту зону - зоной оптимальности гомеостаза (Зотин, Зотина, 1987, 1993; Зотин, 1988). Величина зоны оптимальности является вторым критерием надежности гомеостаза организованной системы.
Если обратится к схеме, показанной на рис.1, то мера надежности связана с наклоном стенки чаши, в которой находится шарик, а зона оптимальности - шириной дна чаши. Чем шире зона оптимальности, тем более надежной является система регуляции гомеостаза. Так как изменение функции внешней диссипации в зависимости от изменения параметра среды описывается уравнением параболы
(149), то определить зону оптимальности можно только некоторым искусственным путем. Мы условились, что зона оптимальности соответствует
ширине зоны изменения параметра х, в пределах которой  остается
остается
минимальной и меняется не более чем на 10% (Зотин, Зотина, 1987, 1993; Зотин, 1988). Исходя из этого, зону оптимальности (z hs) можно вычислять по формуле
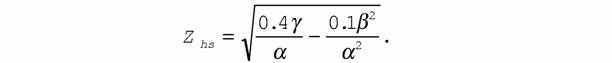
| |
| Учитывая (149), эту формулу можно преобразовать к виду: | |

| (151) |
| Таким образом, мера надежномти Мhs (150)и зона оптимальности Zhs (151) связаны друг с другом соотношением: | |

| (152) |
Предложенные критерии надежности гомеостазов (150) и (151) являются одновременно критериями организованности живых систем, так как они характеризуют совершенство регуляции и управления в организме.
Гомеорез. Как отметил Медников (1973), в процессе прогрессивной эволюции происходит совершенствование не только гомеостазов, но и гомеорезов. Термин гомеорез введен Уоддингтоном (Waddington, 1957; Уоддингтон, 1970) для того, чтобы подчеркнуть стабилизированность и устойчивость процессов развития организмов.
Существуют различные определения понятия гомеореза. По нашему мнению (Зотин, Алексеева, 1984; Зотин, Зотина, 1987, 1993) гомеорез, как и гомеостаз, должен характеризовать не саму устойчивость изменения организованной системы, так как понятие устойчивости стационарного процесса полностью определено интегральным принципом наименьшей диссипации энергии (20, 54), а регуляции этой устойчивости. Естественно, что гомеорез имеет прямое отношение к процессам развития, роста и старения организмов (Alekseeva et al., 1992; Зотин, Зотина, 1993; Zotin, 1995).
Гомеорез представляет собой непрерывный ряд гомеостазов, изменяющихся во времени по определенной траектории. В соответствии с этим гомеорез поддерживает траекторию изменений организованной системы на
минимальном уровне диссипации энергии (рис.2Б). Согласно Уоддингтону (1970) эта наиболее устойчивая траектория носит название креод. Креод характеризуется не только минимальной суммарной интенсивностью потребления кислорода (Зотин, 1988; Зотин, Зотина, 1993), но и оптимальными признаками развития организмов по морфологическим показателям (Waddington, 1957; Уоддингтон, 1970), по фенотипической изменчивости (Захаров, 1987), по вариабельности, длительности тех или иных стадий развития (Ярославцева и др., 1992).
Очевидно, что надежность гомеореза определяется совершенством системы регуляций, которая при этом используется. Поэтому количественные критерии надежности гомеореза, как и гомеостаза, могут быть использованы в качестве критериев организованности живых систем. Было показано, что такие критерии можно получить, идя тем же путем, что и при построении количественных критериев надежности гомеостазов (Зотин, 1988; Зотин, Зотина, 1993). Во-первых, это мера надежности гомеореза Mhr, которая тем больше, чем меньше тратится энергии на сохранение устойчивости изучаемого стационарного процесса при изменении параметров внешней среды. Во-вторых, это зона оптимальности гомеореза Zhr, которая измеряется шириной зоны изменения параметров среды, в пределах которой устойчивость стационарного процесса не изменяется или изменяется мало. Из этих критериев следует, что чем меньше энергетическая стоимость регуляции устойчивости развития, чем шире зона оптимальности, тем надежнее процессы онтогенеза. В заключение следует отметить, что хотя использование метода суммарного потребления кислорода, фенотипической изменчивости и вариабельности стадий развития для изучения надежности гомеореза достаточно универсальны, к настоящему моменту наши знания о мере надежности и зоне оптимальности развития слишком ограничены, чтобы их использовать для характеристики степени организованности животных и, тем более, растений. Можно надеется, что такая возможность когда-нибудь появится.