Зависимость массы мозга от массы тела.
Многие авторы изучали зависимость массы головного мозга от массы тела позвоночных животных (Friedenthal, 1910; Bonin, 1937; Crile, Quiring, 1940; Stephan, Andy, 1964; Jerison, 1970, 1973, 1988; Stephan et al., 1970; Стрельников, 1953, 1970а,б; Sacher, 1976, 1978; Economos, 1980; Martin, 1981; Armstrong, 1982; Hofman, 1982, 1983; Harvey, 1988; Зотин, 1993 б) и показали, что она соответствует аллометрическому соотношению
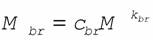 (171)
(171)
где Мbr - масса головного мозга (г); M - масса тела (г); cbr - коэффициент энцефализации; kbr- константа.
Для примера на рис.39 показана зависимость массы мозга от массы тела, основанная на измерении 883 видов млекопитающих (Harvey, Bennett, 1983). Из этого рисунка видно, что масса головного мозга млекопитающих, как и остальных позвоночных животных, сильно зависит от размеров тела. Поэтому, при сопоставлении относительных размеров мозга разных видов позвоночных животных правильней использовать коэффициент энцефализации сbr (171), а не какие-либо иные выражения: он лучше отражает качественные и количественные возможности мозга.
Вместе с тем этот признак (как и многие другие анатомические и физиологические признаки) сильно варьирует даже у животных одного вида. Поэтому для характеристики того или иного таксона важно иметь достаточно большое количество данных о массе мозга и тела - чем больше материалов такого рода мы будем иметь в своем распоряжении, тем точнее будет определены коэффициенты сbr и kbr .
В литературе имеется ряд обзоров о величине мозга и массе тела позвоночных (Никитинко, 1969; Стрельников, 1970а; Jerison, Jerison, 1988; Зотин, 19936). В некоторых работах обнаружена связь величины коэффициента энцефализации сbr с высотой организации позвоночных животных (Martin, 1981; Wyles et al., 1983; Zotin, Lamprecht, 1996). В табл.50 представлены величины коэффициента энцефализации сbr из формулы (171) для разных классов типа позвоночных, рассчитанных по данным, приводимым в монографии Стрельникова (1970а) и в обзорах других авторов (Никитинко, 1969; Блинков, 1972; Ливчак, 1972; Яблоков и др., 1972; Крушинская, Лисицына, 1983; Зотин, 19936).
В принципе коэффициент cbr, как говорилось, можно использовать в качестве одного из критериев организованности животных, так как он косвенно показывает степень совершенства нервной регуляции и разумной деятельности животных, хотя, как видно из табл.50, его увеличение не вполне совпадает с повышением степени организованности позвоночных животных. Это объясняется, как нам кажется, тем, что увеличение относительных размеров мозга и связанный с этим успех в борьбе за жизнь настолько важно для животных, что оно подхватывается естественным отбором независимо от биоэнергетического прогресса.
Таблица 50
Коэффициента энцефализации сbr в разных классах типа позвоночных. N - число изученных видов; tg - время возникновения данного класса в млн. лет (из табл.42). Для позвоночных животных принято kbr=0.56
| Класс | Cbr | N | tg |
| Myxini | 0.00004 | ||
| Cephalaspidomorphi | 0.00059 | ||
| Chondrichthyes | 0.082 | ||
| Osteichthyes | 0.021 | ||
| Amphibia | 0.011 | ||
| Reptilia | 0.015 | ||
| Aves | 0.171 | ||
| Mammalia | 0.440 |
Используя данные о времени возникновения отдельных классов позвоночных (табл.42) и данные о коэффициенте энцефализации (табл.50), можно нарисовать кривую увеличения степени организованности позвоночных в процессе прогрессивной эволюции (рис.40) и выразить ее аналитически в виде формулы:
Cbr= 0.293•ехр(0.014·tg). (172)
Согласно формуле (172) по такому показателю, как коэффициент энцефализации, скорость возрастания организованности позвоночных в процессе эволюции и заметно больше, чем скорость биоэнергетического прогресса (ß=0.0059) и прогрессивной эволюции (ß=0.0063) (140).
Теперь мы приведем расчеты коэффициента cbr, для разных отрядов классов млекопитающих и птиц по данным из тех же обзоров, что и указаных выше (Никитинко, 1969; Стрельников 1970а; Блинков, 1972; Ливчак, 1972;
Яблоков и др., 1972; Крушинская, Лисицына, 1983; Jürgens, Prothero, 1987; Зотин, 19936). Расчет правильным с точки зрения статистики методом для видов млекопитающих и птиц (А.А.Зотин, 1999) показал, что коэффициент для млекопитающих равен kbr=0.575±0.017, a для птиц kbr=0.532±0.020. Очевидно, что различия лежат в пределах ошибки измерений, поэтому мы подсчитали общий для обоих классов коэффициент kbr и получили его равным kbr=0.56. Расчеты показывают:
для млекопитающих Mhr= 0.44·M0.56 , (173)
для птиц Мbr = 0.17·М0.56,
Коэффициент kbr=0.56 мы и приняли для обоих классов при вычислении коэффициента энцефализации в отдельных отрядах, семействах и видов млекопитающих и птиц. Сопоставление коэффициента энцефализации с временем существования того или иного таксона показано в табл.51. По данным, приведенным в этой таблице можно построить кривую прогресса организованности в реальной эволюции млекопитающих и птиц (рис.41). Получаем типичные экспоненты, которые можно выразить с помощью следующих формул (сведенья времени возникновения отдельных отрядов птиц взяты из руководства Кэрролла, 1993 а):
Млекопитающие -> Сbr = 1.25 · ехр(0.026 · tg ), (174)
Птицы -> Cbr=0.14·ехр(0.002·tg). (175)
Судя по этим расчетам, эволюция относительных размеров мозга в классе млекопитающих и птиц шла с разной скоростью, причем млекопитающие эволюционировали быстрее птиц. Формула (174) показывают также, что эволюционный прогресс организованности по коэффициенту энцефализации, шел примерно с такой же скоростью, как по сопоставимой продолжительности жизни млекопитающих (170) и собственно прогрессивной эволюции (137).
Показано, что размеры мозга соответствуют энергетическому
метаболизму животных (Стрельников, 1953, 19706; Hopson, 1977). Это естественно, так как согласно термодинамике информационных процессов, чем совершенней регуляция и управление в термодинамической системе, тем больше она требует затрат энергии. Согласно же нашему допущению совершенство управления процессами, протекающими в животном организме, как и рассудочная деятельность, коррелируют с относительными размерами мозга. Однако, учитывая, что при биоэнергетическом прогрессе млекопитающих ß=0.0068, можно считать, что прогресс организованности в классе млекопитающих, хотя и связан с биоэнергетическим прогрессом, но шел быстрее, чем последний.
Таблица 51
Коэффициента энцефализации сbr в разных отрядах (и некоторых семействах) млекопитающих и птиц. N - число изученных видов; tg - время таксона в млн. лет назад (из табл.42).
| Отряд | Cbr | N | tg | Отряд | Cbr | N | tg |
| Класс Mammalia | Класс Aves | ||||||
| Marsupialia | 0.150 | Sphenisciformes | 0.036 | ||||
| Insectivora | 0.085 | Struthioniformes | 0.059 | ||||
| Chiroptera | 0.113 | Galliformes | 0.079 | ||||
| Rodentia | 0.148 | Apodiformes | 0.097 | ||||
| Lagomorpha | 0.151 | Columbiformes | 0.098 | ||||
| Edentata | 0.241 | Gruiformes | 0.109 | ||||
| Hyracoidea | 0.265 | Charadriiformes | 0.118 | ||||
| Carnivora | 0.350 | Pelecaniformes | 0.121 | ||||
| Artiodactyla | 0.378 | Anseriformes | 0.123 | ||||
| Perissodactyla | 0.390 | Ciconiiformes | 0.150 | ||||
| Pinnipeda | 0.567 | Falconiformes | 0.166 | ||||
| Primates | 0.774 | Passeriformes | 0.178 | ||||
| Сем. Hominidae | 2.758 | Сем. Corvidae | 0.297 | ||||
| Proboscidea | 1.204 | Piciformes | 0.247 | ||||
| Cetacea | 1.324 | Strigiformes | 0.271 |
| Ceм. Delphinidae | 1.684 | Psittaciformes | 0.329 |
Прогресс организованности млекопитающих в процессе эволюции, учитываемый с помощью коэффициента энцефализации, установлен и на палеонтологическом материале (Jerison, 1961). На рис.42 показаны собранные этим автором данные о зависимости массы мозга от массы тела, рассчитанных по обнаруженным остаткам млекопитающих олигоцена и эоцена по сравнению с современными млекопитающими. Согласно проведенным нами расчетам (Зотин, Зотин, 1995) по данным, приведенным на рис.43, коэффициент энцефализации млекопитающих в эоцене равнялся 0.026, в олигоцене - 0.055 и у современных млекопитающих - 0.115.