рефераты конспекты курсовые дипломные лекции шпоры

- Раздел История
- /
- Номогенетический аспект законов Менделя
Реферат Курсовая Конспект
Номогенетический аспект законов Менделя
Номогенетический аспект законов Менделя - раздел История, ВЕК ГЕНЕТИКИ: ЭВОЛЮЦИЯ ИДЕЙ И ПОНЯТИЙ Изменчивость Представлялась Ч. Дарвину И Его Современникам Не...
Изменчивость представлялась Ч. Дарвину и его современникам неограниченной, беспорядочной, идущей во всех направлениях. Организм обладал как бы "восковой пластичностью". Принятие этого исходного хаоса изменений, или, как назвал А. А. Любищев, постулата о "тихогенетическом характере изменчивости", существенно для дарвинистов. Законы Менделя подчинили этот хаос математическим закономерностям. Число изменений при гибридизации вводилось в четкие рамки расщепления. До Г. Менделя многообразие форм в потомстве гибридов объясняли "ослаблением силы наследственности", не было научных принципов описания и изучения гибридов. Поэтому исследователи "терялись в противоречивых частностях и сложной мозаике признаков" (Гайсинович, 1988). Предшественники Г. Менделя наблюдали все характерные черты поведения признаков при гибридизации: доминирование (Т. Э. Найт), комбинирование в потомстве (О. Сажрэ) и принцип единообразия первого поколения и расщепления (П. Нодэн).
Но лишь Мендель установил четкие количественные закономерности в наследовании признаков в ряду поколений, куда непротиворечивым образом вошла вся предшествующая феноменология. Установленные Менделем законы наследования признаков (единообразие первого поколения, расщепление и независимое комбинирование) являются естественнонаучными законами, а не правилами, как иногда пишут. Ибо для этих законов: а) определена точная сфера применения, за пределами которой они нарушаются; б) они дают возможность предсказания и опытной проверки и в) возможность количественного описания и математической формулировки.
Выдающейся чертой менделевской работы было также постулирование связи наследования признаков с дискретными факторами половых клеток. Высказанная Менделем гипотеза чистоты гамет явилась как бы предвидением поведения хромосом в мейозе. Эта вторая сторона работы Менделя имела и имеет всеобщее значение на протяжении всего развития генетики. Но такой номогенетический (в противоположность тихогенетическому, т. е. основанному на случае) подход к анализу явлений изменчивости и наследственности не был понят не только современниками Менделя, но и дарвинистами даже спустя 35–40 лет.
Книга Ч. Дарвина "Прирученные животные и возделанные растения" (см. русский пер. 1900) поражает не только энциклопедичностью и объемом собранных сведений в области ботаники, зоологии, растениеводства, животноводства, биологии человека. Во многих случаях Ч. Дарвин сообщает о проделанной им лично экспериментальной работе с присланными ему со всего света материалами, начиная от морфологического, анатомического и статистического анализа и, кончая опытами по скрещиванию растений, разведению разных пород птиц (особенно голубей) и животных. Обсуждая, например, изменчивость пчел, он сообщает: "Мне прислали улей, наполненный мертвыми пчелами из Ямайки, куда они переселены с очень давних пор, и после тщательного сравнения их под микроскопом с моими собственными пчелами я не нашел ни малейшего следа различия" (Дарвин, 1900, с. 193).
В книге Дарвина цитируются и обсуждаются данные всех гибридизаторов растений, предшественников Менделя — И. Гэртнера, Т. Э. Найта, О. Сажрэ, П. Нодэна. Среди приведенных Ч. Дарвиным рисунков есть и рисунок, показывающий различия в изменчивости разных сортов гороха по форме стручка и семян — признаки, которые изучал и Мендель. Доходит очередь, скажем, до обсуждения изменчивости картофеля и Ч. Дарвин сообщает, что он посадил 18 сортов в смежных рядах и оценил характер изменчивости их признаков, от формы и окраски цветков и плодов до величины и формы клубней (Ibid., с. 218).
Но самое поразительное другое. При скрещивании нормальной и пелорической формы львиного зева Дарвин получил чисто менделевское соотношение 3:1, но не проник в "душу фактов" (выражение Анри Пуанкаре), как это гениально удалось Г. Менделю. С точки зрения истории науки поучительно проследить за характером опытов и мыслью Ч. Дарвина. "Я скрестил пелоричный львиный зев Antirrhinum majus с пыльцой простой формы, а эту простую форму с пыльцой пелоричного цветка. У меня получилось, таким образом, две большие гряды сеянцев, из которых ни один не был пелоричен. Нодэн получил те же результаты, скрестивши пелоричную линарию с простой формой. Я тщательно исследовал цветки 90 растений скрещенного Antirrhinum и увидел, что строение их вовсе не подвергалось изменению вследствие скрещиваний, за исключением того, что в немногих случаях маленькие зачатки пятой тычинки, существующей всегда, были развиты больше обыкновенного или вполне. Не следует думать, что совершенное исчезновение пелоричного строения у скрещенных растений зависит от полной неспособности к наследственной передаче, потому что я вырастил огромную гряду растений от пелоричных Antirrhinum, и 16 растений, переживших зиму, были столь же пелоричны, как и их родители. Это дает нам хороший пример той значительной разницы, которая существует между наследственностью признака и силой передачи их скрещенному потомству" (Дарвин, 1900, с. 328).
Иными словами, Ч. Дарвин провел реципрокные скрещивания и показал, что признак пелоричности наследственен и рецессивен. Теперь надо получить F2 и провести количественный учет расщепления, что Ч. Дарвин и сделал. Во втором поколении из 120 сеянцев 88 оказались простыми, два имели промежуточное строение между пелоричным и нормальным, а 37 были совершенно пелоричны, возвратившись, таким образом, по строению к одному из своих прародителей. Прекрасное соответствие с теоретически ожидаемым по Менделю 3:1. Но у Дарвина, в отличии от Менделя, не было ни теории, ни ожидания, т. е. всего того, что позволяет предсказывать результаты и давать истолкование отклонениям.
Получив типично менделевское соотношение, Дарвин делает вывод, что наклонность производить нормальные цветки "преобладала в первом поколении, тогда как наклонность к пелоричности, усиливавшаяся, по-видимому, в промежуточном поколении, появилась во втором поколении сеянцев... Впрочем, говоря вообще, вопрос о преимущественной передаче чрезвычайно запутан... вовсе не удивительно, что все попытки вывести какое-либо общее правило для преимущественной передачи оказались неудачными" (Дарвин, 1900, с. 328).
Мне кажется маловероятным предположение, к которому присоединяется А. Е. Гайсинович (1989), что если бы Дарвин ознакомился с работой Г. Менделя, то он полностью оценил бы ее значение. В обширной историко-научной сводке "Рост биологической мысли" Э. Майр подробно обсуждает судьбу открытия Менделя. Он также приходит к выводу, что если бы Дарвин был знаком с работой Менделя, "она вряд ли оказала на него какое-либо влияние. Ибо потребовалось много лет (даже после 1900 г.), когда "чистые дарвинисты", как они сами себя называли, поняли, что постепенная эволюция и непрерывная изменчивость могут быть объяснены в менделевских терминах. Видимо, и Ч. Дарвин столкнулся с той же самой трудностью. Он знал о работе О. Сажрэ, но она не помогла ему понять изменчивость" (Mayr, 1982, с. 725). Одно дело наблюдать, сталкиваться с фактами, или знать о них, а другое дело — понять их внутренний смысл в рамках определенной концепции.
Второй создатель теории естественного отбора А. Уоллес, вполне знакомый с работами первых генетиков, в 1908 г. в статье "Современное положение дарвинизма" писал, что закономерности Менделя касаются лишь резких качественных различий, что это частный случай, да притом хорошо известный Ч. Дарвину, и нечего развертывать сложные диаграммы и таблицы, "трубя в трубы и уверяя в их важном значении" (Уоллес, 1911). Любопытно, что спустя полвека, частным случаем большинство генетиков считало открытие Б. МакКлинток мобильных элементов. Инвариантный психологический феномен в истории науки — отнесение их к исключениям и экзотике — своего рода иммунная защита научного сообщества против новых, требующих ревизии открытий.
Можно задаться и обратным вопросом. А как бы Мендель отнесся к концепции Дарвина? Его точному аналитическому уму, видимо, претило расплывчатое дарвиновское представление о наследственной изменчивости и построенная на этой зыбкой базе концепция. На сей счет есть определенное свидетельство. "Хотя Г. Мендель посетил Англию в 1862 г., куда он поехал на Всемирную выставку, но с Дарвином он не познакомился. Возможно, что в то время он мало знал о Дарвине. В его библиотеке есть труд Дарвина "О происхождении видов" в немецком переводе Бронна, но только во втором издании, вышедшем в 1863 г. Заметки Менделя на полях этой книги, а также на полях обоих томов "Изменений животных и растений при одомашнивании" (имеющихся в библиотеке Менделя также в немецком переводе 1868 г.) показывают, что Мендель был несогласен со многими положениями Дарвина, в частности, по вопросам гибридизации и пангенезиса. Да он и не послал Дарвину своего оттиска. Отношение Менделя к эволюционному учению Дарвина с полной определенностью выразилось в его выступлении после лекции о теории Дарвина, прочитанной моравским палеонтологом и геологом А. Маковским в марте 1865 г. в научном обществе в Брно. Дав положительную оценку учению Дарвина, Маковский предоставил слово Менделю (по его просьбе), и Мендель заявил, что он придерживается учения о постоянстве видов" (Матуошкова Б., Матуошкек О., 1959). Итак, круг замкнулся. Налицо взаимное отталкивание двух подходов: номогенетического и исторического.
2.2.2. Ограничение сферы отбора: понятия генотипа и фенотипа
Хотя Ч. Дарвин придавал основное значение в отборе неопределенным и наследуемым уклонениям, однако, у него не было ясности в том, как же наследуются приобретенные в течение жизни свойства. Хаос противоречивых фактов без надлежащей теории не давал возможности прийти к определенным выводам. Ч. Дарвин заключил: "Мы смотрим на наследственность как на правило, а на ненаследственность как на аномалию. Однако свойство это, в нашем незнании, кажется нам каким-то капризным. Мы видим, что оно передает признаки то с непонятной для нас энергией, то с чрезвычайной слабостью" (Дарвин Ч., 1900, с. 335). Не было достаточной ясности в том, можно ли усилить или ослабить отбором имеющийся или вновь появившийся признак.
За экспериментальный анализ этого вопроса взялся двоюродный брат Ч. Дарвина, выдающийся ученый Ф. Гальтон (ему принадлежит метод близнецового анализа у человека). Ф. Гальтон собрал данные о росте родителей и детей в английских семьях. Кроме того, он проследил за изменением величины семян душистого горошка в двух поколениях. В обоих случаях статистический анализ популяционных данных показал, что отклонения от средней величины родителей частично передавались потомству. Ф. Гальтон сформулировал закон регрессии, из которого следовало, что каждая индивидуальная особенность родителей проявляется и у потомков, но в среднем в меньшей степени. А отсюда как бы следовал вывод о пластичности наследственности и о творческом характере отбора, путем которого можно добиться смещения средней величины признака в желательном направлении.
Основатель биометрии К. Пирсон сделал расчет, согласно которому через шесть поколений можно любое отклонение от средней зафиксировать с помощью отбора в популяции. И Ф. Гальтон и К. Пирсон стояли на позициях слитной наследственности. Биометрикам казалось, заметил Ю. А. Филипченко, что они создали новую эпоху в изучении наследственности и в скором времени выяснят законы последней, облеченные в строгую математическую форму.
Эти надежды оказались ошибочными, как было экспериментально показано датским генетиком В. Иогансеном в его опытах на генетически однородном материале — чистых линиях. В генетически однородном материале отклонения родителей от средней не наследуются, а в смеси чистых линий (в популяции) отбор мог привести к смещению средних значений. В 1903 Иогансен пришел к четкому выводу, что "подбор только отбирает представителей уже существующих типов; эти типы отнюдь не создаются подбором постепенно — они лишь отыскиваются и изолируются" (Иогансен, 1933). Иными словами, пришлось ограничить представление об отборе как о некоем творческом, действенном начале.
История спора В. Иогансена с биометриками-дарвинистами показывает, что одно применение математики не только не превращает "неточную" науку в "точную", но может затемнить существо дела. Точной наука становится тогда, когда она вырабатывает достаточно четко определенные понятия. Таковыми были предложенные В. Иогансеном понятия "ген", "генотип", "фенотип", "чистая линия", а также введенные ранее В. Бэтсоном понятия "гомозигота", "гетерозигота", "аллеломорф". Математические методы эффективны, если имеется адекватное биологическое обоснование. "Мы должны разрабатывать учение о наследственности с помощью математики, но не как математическую дисциплину", — вот вывод В. Иогансена, верный и для наших дней.
2.2.3. Мутационная теория де Фриза в аспекте видообразования
Ч. Дарвин разделял положение о том, что природа не делает скачков. Здесь заключался принимаемый дарвинистами постулат о непрерывности и о неограниченности изменчивости. Принималось, что различия между разновидностями и видами обязаны возникновению мелких непрерывных изменений. Хотя Ч. Дарвин знал, что иногда наблюдаются резкие отклонения от нормы, он считал, что эти своеобразные "уродцы" очень редки и в силу слабой жизнеспособности не имеют эволюционной судьбы.
Этот вывод Ч. Дарвина и его последователей был поставлен под сомнение исследованиями Г. де Фриза, который по праву считается основателем мутационной теории, основные положения которой остаются верны и сегодня (Лобашев, 1967, с. 287). К этому можно добавить, что к славе Г. де Фриза как переоткрывателя законов Менделя следует добавить два важных пионерских концептуальных представления: 1) о нестабильном состоянии генов и 2) о существовании критических периодов в проявлении наследственных признаков (Фриз, 1932; Vries, 1912).
Подробно рассматривая динамику идей в области учения о макроэволюции, В. И. Назаров (1991) приходит к выводу, что де Фриз высказал три кардинальных положения, составивших ядро современной теории прерывистого равновесия и находящихся сейчас в фокусе острых дискуссий: идею о неравномерности темпа мутирования и эксплозивности видообразования, представление о двух разных путях видообразования — "групповом" и "филогенетическом" (это близко к современным представлениям о кладистическом и филетическом видообразовании), а также идею независимости акта видообразования от естественного отбора, борьбы за существование.
С точки зрения истории и методологии науки интересно, что нередко исследователь специально отбирает тот материал для исследования, который соответствует его интуитивной гипотезе. Мендель специально выбрал горох как жесткий самоопылитель и специально выбрал для исследования четкие единичные признаки, поведение которых можно было с легкостью изучать в ряду поколений. Он писал, что "выбор растительной группы, которая будет служить опытам подобного рода, должен быть сделан с наивозможной осторожностью, если мы не хотим подвергнуть риску самый успех опыта". Напротив, де Фриз подыскивал для работ по анализу наследственной изменчивости вид растений, который в силу каких-то внутренних причин отличался бы сильной изменчивостью. Только после того, как Г. де Фриз просмотрел 100 видов, он остановился на энотере. И его выбор оказался фантастически удачен для задуманных целей.
На одном заброшенном картофельном поле вблизи деревни Гильверзум в 1886 г. он обратил внимание на популяцию энотеры, растения, ввезенного из Америки и одичавшего в Европе. В следующем 1887 г. он нашел на этом же поле двух мутантов (забегая вперед в терминологии) и заложил многолетний опыт по изучению частоты возникновения мутантов.
Энотера оказалась уникальной в смысле своей генетической конституции, а изучение закономерностей ее наследственной изменчивости и видообразования таит в себе еще множество загадок. Работая с энотерой, Г. де Фриз получил от первых высеянных в 1886 г. девяти растений около 53 тыс. потомков в период 1886–1899 гг. и среди них около 800, или 1,5% форм, имеющих резкие отклонения от исходного типа. Эти отклонения были названы мутациями. Мутации возникали внезапно, непредсказуемо, в разных направлениях. В ряде случаев, отдельные мутации захватывали сразу множество признаков и полностью изменяли габитус растения, причем эта совокупность признаков передавалась как дискретная единица. Такие резко отличающиеся друг от друга формы, возникающие внезапно, де Фриз назвал "элементарными видами". К их появлению отбор никакого отношения не имеет. Что касается вида в дарвиновском понимании, то он, по мнению де Фриза, представляет собой совокупность "элементарных видов", впоследствии названных линнеоном.
Совершенно сходные идеи в работе "Гетерогенезис и эволюция. К теории происхождения видов" развил необычайно талантливый ученый, директор ботанического сада при Петербургском университете, академик С. И. Коржинский (1861–1900), проживший всего 39 лет. Гетерогенезисом был назван процесс внезапного сальтационного появления форм, отличающихся комплексом новых признаков. "Эти признаки обладают большим постоянством и неизменно передаются по наследству из поколения в поколение. Таким образом, сразу возникает новая раса столь же прочная и постоянная, как и те, которые существуют с незапамятных времен" (цит. по: Филипченко, 1977, с. 162).
Что касается происхождения садовых растений, то С. И. Коржинский категорически утверждал, что во всех известных ему случаях новые разновидности возникли путем внезапных отклонений от исходных форм. В своих лекциях по генетике, опубликованных в Мюнхене в 1911 г. и в русском переводе два года спустя, Рихард Гольдшмидт заключает, что "среди ботаников заслуга накопления наибольшего числа фактических данных, имеющих отношение к внезапному возникновению растительных форм, принадлежит русскому ботанику академику С. И. Коржинскому" (Гольдшмидт, 1913, с. 135).
Инициированные де Фризом исследования по генетике, цитологии и систематике в роде Oenothera, привели к открытию фантастических механизмов в организации генетической системы размножения, способах наследственной изменчивости и видообразования. В сводке по цитогенетике (Суонсон, Мерц, Янг, 1969) ситуация с энотерой образно названа "бунтом против эволюции". Здесь оказалось "можно то, что нельзя", а именно: сочетание таких элементов генетической системы, каждый из которых сам по себе вреден, а в комбинации дает вполне удовлетворительный результат. Среди них: 1) вовлечение группы или целого набора хромосом в реципрокные транслокации, что обычно приводит к резкому падению фертильности вследствие хромосомной нестабильности гамет и зигот; 2) накопление летальных мутаций; 3) резкое ограничение рекомбинаций, когда транслокационный комплекс передается как целое; 4) неспособность целого набора хромосом проходить либо через женский, либо мужской гаметогенез.
И несмотря на это, завезенные два-три века назад в Европу из Северной Америки растения рода Oenothera образовали сложные гибридные формы видового ранга, по терминологии В. Гранта (1984) "гетерогамные микровиды". Они распространились в Европе как сорняки, а вид Ое. lamarckiana, который специально был выбран де Фризом для исследования, не только распространился по всей Европе, а позднее вновь утвердился в Америке!
Распространено мнение, к сожалению кочующее по разным учебникам и сводкам, что де Фриз обнаружил не мутации, а лишь редкие рекомбинации, выщепляющиеся в потомстве транслокационных гибридов. Но это, во-первых, не так. А во-вторых, в случае энотеры трудно отличить мутации и рекомбинации. С современных позиций очевидно, что процессы мутации и рекомбинации на молекулярном уровне переплетены и разграничение это нередко вопрос терминологии (Хесин Р. Б., 1984, с. 279). Г. де Фриз обнаружил три типа мутаций: генные, хромосомные и геномные. Среди типично генных были "brevistytis" с коротким столбиком цветка и nana — карликовые растения (Cleland, 1935,1962; Грант, 1984; Burnham, 1980).
К хромосомным мутантам, обнаруженным Г. де Фризом и его учениками в первые же годы, относятся трисомики (2n = 15 вместо 2n = 14). Сначала де Фриз выделил их как относительно константные формы, отличающиеся множеством признаков, и лишь затем был определен их хромосомный статус как трисомиков.
Он впервые обнаружил формы, которые в терминах классической генетики соответствуют геномным мутациям. А именно, полиплоидную форму gigas, которая возникла на 4-й год его опытов среди около 14000 исследованных растений. И, наконец, в разные годы среди в общей сложности 54 000 исследованных растений де Фриз зафиксировал возникновение так называемых "полумутантов" (частота их появления 0,06%). Растения этого типа, названные rubrinervis, характеризовались красными жилками на листьях, широкой красной полосой на чашечке и плодах и слабым развитием луба.
Как позднее было показано (Cleland, 1962), кариотип исходного вида Ое. lamarckiana, с которым работал Г. де Фриз, имел 12 пар хромосом, объединенных в транслокационное кольцо, и одну пару свободную. А "полумутант" rubrinervis имел 4 свободных пары и 6 пар, объединенных в кольцо. Для возникновения такой формы в потомстве отдельных растений рекомбинационным путем необходимо, чтобы в отдельных половых клетках одномоментно происходили множественные рекомбинационные обмены, причем в определенных участках! Такое событие можно представить себе только в рамках современной генетики, когда стало известно, что множественные обмены могут происходить в местах локализации мобильных элементов, а также быть следствием так называемых "транспозиционных взрывов" и множественных упорядоченных транспозиций (Engels, 1983; Герасимова, 1990; Гвоздев, Кайданов, 1986, 1990). Можно суверенностью полагать, что подобные события будут описаны и у энотеры, исследование которой сулит множество открытии в области механизмов наследственной изменчивости.
В эволюционной генетике, помимо энотеры, доводы в пользу множественных одномоментных хромосомных перестроек получены при исследовании хромосомного полиморфизма у роющих грызунов — слепушонок группы Ellobius tancrei, относящихся к надвиду Е. talpinus. Здесь были открыты так называемые "робертсоновские веера" с варьированием в числе хромосом от 54 до 32. При этом получены доводы, что "54 и 32-хромосомные кариотипы должны были возникнуть за счет резкой скачкообразной перестройки всего кариотипа в целом, приведшей к одномоментному слиянию 16–20 акроцентрических хромосом и 8–10 метацентриков" (Воронцов, 1988, 1999).
Наконец, с точки зрения путей видообразования де Фриза следует считать первооткрывателем двух новых типов видообразования: 1) хромосомного, в частности полиплоидного и 2) гибридогенного. В случае гибридогенного видообразования один вид содержит два разных хромосомных комплекса, которые передаются потомству как целое и в разных комбинациях скрещивания "порождают" новые виды. Ученик и последователь де Фриза О. Реннер открыл, что геном вида Ое. lamarckiana, с которым работал де Фриз, состоит из двух транслокационных комплексов gaudens и velans. В них все 7 пар хромосом связаны взаимными обменами и передаются совместно как одна гигантская хромосома.
При скрещивании растений вида Ое. lamarckiana (комплексы velans/gaudens) и Ое. strigosa (комплексы deprimens/stringers) получаются все четыре возможные комбинации, имеющие статус вида и распространенные в природных популяциях (Грант В., 1984, с. 368). Другими словами, в каждом скрещивании происходит "порождение видов", которое можно наблюдать воочию!
Открытые у энотеры транслокационные гибридные видовые системы, названные комплексами Реннера, обладают еще двумя поразительными особенностями. Во-первых, летальностью каждого из них в гомозиготе, так что возникает сбалансированная перманентная гетерозиготность, когда выживают только гетерозиготы. Именно этот, открытый в опытах де Фриза феномен, натолкнул Г. Меллера на идею использовать метод сбалансированных леталей для создания генетического метода количественного учета возникновения мутаций у дрозофилы. Во-вторых, один из реннеровских комплексов передается только через яйцеклетку, а другой — только через пыльцевые клетки, т. е. в гаметогенезе у гибридогенного вида происходит элиминация одного из геномов.
Кратко суммируем значение работ де Фриза для генетики видообразования и эволюционной теории. Г. де Фриз установил неравномерность темпа эволюционного процесса, наличие вспышек мутаций, которые происходят в "мутационные периоды". В период этих вспышек возникают скачкообразные мутации, резко меняющие облик растений, вызывающие резкие изменения в системе воспроизведения, в результате чего сальтационно возникают формы видового ранга. Подобные сальтационные изменения могут происходить повторно. В опытах Г. де Фриза и его учеников и последователей на энотере было открыто хромосомное видообразование (полиплоидия) и гибридогенные виды, образованные на основе транслокационных комплексов.
Справедливо считается, что "классическая дарвиновская концепция понимает видообразование как постепенный градуалистический процесс" (Воронцов, 19886). Генетические исследования де Фриза доказали возможность хромосомного сальтационного (полиплоидия) и гибридогенного видообразования. Оно оказалось повсеместно распространенным у растений. У покрытосемянных растений около 52–58% видов являются полиплоидами, по преимуществу это аллополиплоиды, растения объединяющие геномы разных видов. Причем, с такими сложными переплетениями полового, бесполого, гибридного размножения, с такими поразительными системами естественной хромосомной инженерии, что приходится расширить концепцию вида до разных по емкости видовых единиц (Грант, 1984, Малецкий, 1995).
Ведущее концептуальное открытие в этой области было сделано в 1917 году датским генетиком О. Винге, который предположил, что у межвидовых гибридов в случае удвоения у них числа хромосом, могут восстановиться мейоз и плодовитость. "Настоящее исследование имеет непосредственное отношение к этой идее. Оно может рассматриваться как экспериментальное обоснование теории гибридного происхождения полиплоидных видов", — таково введение к классической работе Г. Д. Карпеченко (1927). Он впервые синтезировал новую неизвестную в природе видовую форму Raphanobrassica, константный полиплоидный межродовой гибрид между редькой и капустой. Совершенно справедливо Н. Н. Воронцов (1999) называет синтез рафанобрассики первым случаем конструирования нового генома, того, что в конце 70-х стало называться генетической инженерией.
Через три года шведский генетик Арне Мюнтцинг впервые осуществил ресинтез дикорастущего в природе аллополиплоидного вида пикульника рода Galeopsis. Природная хромосомная инженерия создает гибридогенные полиплоидные комплексы видов, открытые и изученные американским ботаником Ледьярдом Стеббинсом. В этих комплексах геномы нескольких диплоидных исходных видов могут вступать между собой во всевозможные гибридные аллотетраплоидные комбинации. Объединяться могут сразу несколько геномов, так что предком одного вида может ни один, а несколько видов, как, например, у обычной мягкой пшеницы, у видов хлопчатника (Грант, 1984).
Неожиданно выяснилось, что гибридогенное, сходное в принципе с энотерой, видообразование регулярно встречается в ряде групп позвоночных и беспозвоночных животных. Гибридогенное видообразование описано уже у 50 видов позвоночных (Боркин, Даревский, 1980, 1989; Кирпичников, 1988; Tunner, Heppich-Turmer, 1991; Даревский, 1995). Оно принимает три основные формы: партеногенез (спермин не нужны для развития яйцеклеток видов-гибридов); гиногенез (спермин нужны лишь для активации развития, но развитие происходит на основе женских гамет и наследование матроклинно); и собственно гибридогенез, когда гибридный вид образуется на основе гибридных зигот, но один из родительских геномов селективно элиминируется в клетках полового пути в ходе гаметогенеза и мейоза.
Гибридогенез оказался широко распространен у скальных ящериц рода Lacerta, живущих в горных районов Закавказья. Среди 18 близких видов комплекса L. saxicola 7 имели гибридогенное происхождение. Все они состояли из партеногенетических самок, способных размножаться без самцов. Но они способны также скрещиваться со своими собратьями из двуполых видов и порождать новые гибридогенные виды. В последнем варианте примерно в 10% случаев возникают триплоиды. Причем у некоторых групп ящериц, вьетнамских гекконов и агам, триплоиды оказались плодовитыми Поскольку гибридогенез здесь является нормой, то (1) одни и те же партеновиды порождаются вновь и вновь в разное время и (2) одни и те же двуполые виды способны вступать в разные гибридогенные связи и порождать таким образом разные виды! (Даревский, Гречко, Куприянова, 1999).
У животных обнаружено также удивительное явление избирательной элиминации одного из геномов получило название "геномное исключение". У гибридного вида хромосомы одного из родительских видов как бы одалживаются для построения соматических тканей. Вот почему было предложено именовать такой способ видообразования как "кредитогенез" (Боркин, Даревский, 1980). Элиминация одного из родительских геномов в клетках полового пути может быть названа также как "геномоцид". Это явление было описано и у растений в случае отдаленных гибридов ячменя. Природный гибридогенез в сочетании с геномоцидом в зародышевом пути был обнаружен у некоторых видов беспозвоночных.
Особенно поразительной оказалась гибридогенная система у палочников рода Bacillus, где сосуществуют пять способов воспроизведения (Giorgi, 1992; Mantovani, Scali, 1992):
1) обычное бисексуальное размножение, 2) гибридогенез, 3) партеногенез, 4) гиногенез и 5) андрогенез — первый случай такого рода, обнаруженный как регулярное событие в природных популяциях (схема на рис. 1).
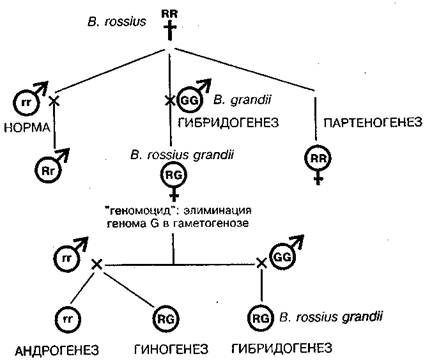
Рис. 1. Гибридогенез у видов палочника (род Bacillus) как пример недарвиновского видообразования, открытого на энотере Г. де Фризом. R и r обозначают геномы вида В. rossius у самок и самцов, G — геном В. grandii (no Mantovani, Scali, 1992; Giorgi, 1992).
2.2.4. Мутационное видообразование и биогеография. Исследования Виллиса и их оценкаде Фризом
Как известно, резкий переход Томаса Моргана от эмбриологии к генетике совершился под влиянием работ по теории мутаций Гуго де Фриза (Allen G., 1985; Музрукова, 1988, 1993). В 1904 г., через год после выхода второго тома своего труда "Мутационная теория", Г. де Фриз был приглашен в Америку для чтения лекций в университете Беркли, Калифорния. Книга его лекций под названием "Species and Varieties. Their origin by mutation" вышла в Америке первым изданием в 1904, вторым изданием в 1905 г. и третьим изданием — в 1912 г. (Vries H., 1912). Редактор, глава отдела ботаники Карнеги института в Вашингтоне МакДуглас, предпослал к книге три характерных эпиграфа:
– Происхождение видов есть явление природы. Ламарк.
– Происхождение видов есть объект изучения. Дарвин.
– Происхождение видов — объект эксперимента. Г. де Фриз,
Сам Г. де Фриз в предисловии к вышедшим лекциям отметил, что его исследования, находятся в полном соответствии с принципами Дарвина и касаются лишь углубленного анализа некоторых аспектов наследственности, изменчивости, отбора и мутаций, которые ранее были неясными. Однако спустя 20 лет Г. де Фриз более четко изложил свою позицию и свои научные симпатии.
В 1923 г. он опубликовал в авторитетном генетическом журнале "Journal of Heredity" статью под названием "Изложение теории Дж. Виллиса о происхождении видов" (Vries H., 1923). Ни эта статья, ни причины ее написания, ни результаты исследований Дж. Виллиса и его теория почти не обсуждаются и не цитируются в научно-исторической литературе, посвященной развитию эволюционной теории и ее связям с генетикой. Хорошо, например, известна концепция "макромутаций" Р. Гольдшмидта, предложенная в 1940 г. (Goldschmidt R. В., 1940). Она обычно упоминается во многих сводках по СТЭ как несостоятельная, или надуманная. И только с конца 70-х годов ситуация изменилась (Gould S. J., 1977; Воронцов Н. Н., 1988а, б).
А между тем, на что специально обратил внимание В. Назаров (1991), Р. Гольдшмидт в числе своих непосредственных предшественников называет только двух ботаников-систематиков — Г. Гаппи и Дж. Виллиса. Р. Гольдшмидт цитирует отрывок из книги J. С Willis (1922), где говорится, что для видообразования "одной большой и жизнеспособной мутации, встречающейся на участке земли в несколько квадратных ярдов и, возможно, один раз в пятьдесят лет, по-видимому, будет достаточно. Шансов заметить такую мутацию практически нет". Обсуждая недавно концептуальные аспекты теории Дж. Виллиса и математическую трактовку его данных, сделанную в 1924 г. английским математиком Юлом, Ю. В. Чайковский (1991) констатирует, "что ученый мир пренебрег и самими наблюдениями и фактами многократного точного совпадения этих наблюдений с простой и ясной теорией".
Причина такого небрежения состоит в номогенетическом характере концепции Дж. Виллиса, в отрицании одного из ключевых положений теории Дарвина.
Джон Кристофер Виллис выпустил две книги в издательстве Кэмбриджского университета. Первая из них вышла в 1922 г. и называется "Age and Areal" или "Возраст и Ареал" (Willis J. С., 1922) с подзаголовком "Изучение географического распределения и происхождения видов". В книгу включен ряд глав, написанных другими авторами, в том числе и статья Г. де Фриза "Возраст и ареал" и мутационная теория". В 1940 г. Дж. Виллис выпустил вторую книгу "Ход эволюции" с оппозиционным подзаголовком "Дифференциация и дивергентная мутация нежели естественный отбор" (Willis, 1940).
В период с 1896 г. по 1911 г. Дж. Виллис проводил ботанико-географическое изучение о. Цейлона, Южной Индии, Новой Зеландии, затем занял пост директора Ботанического сада Рио де Жанейро. "Гипотеза, которую я назвал "Возраст и Ареал", не случайное открытие, я пришел к ней постепенно, в результате почти 20-летней работы, детально изучая флору Цейлона и сопредельных стран" (Willis, 1922).
Отметить две важные оригинальные особенности исследования Дж. Виллиса:
1) количественный статистический учет числа видов в роде и числа родов в семействе в сопоставлении с характером их встречаемости; 2) сопоставление полученных распределений в разных ареалах.
Во флоре о. Цейлона среди 2809 видов покрытосеменных 809 видов были эндемичными для острова. При этом область распространения около 200 эндемиков была ограничена группой гор или даже отдельными горами. Среди 1027 родов растений 23 рода — эндемичны для о. Цейлона. При этом большинство родов было представлено одним видом, четыре рода — по 2–3 вида и только в двух эндемичных родах было больше десяти видов. Дж. Виллис разделил по степени распространения все виды на 6 классов от 1 (очень распространен) до 6 (очень редок), т. е. чем больше значение, тем реже встречаемость. Далее сопоставлялась встречаемость видов на о. Цейлоне и в Южной Индии (материк).
Общий результат был таков: виды, которые широко распространены на о. Цейлон, были частыми и на материке, в Индии. Чем реже встречался вид на о. Цейлон, тем больше была вероятность обнаружить его среди эндемиков. Виллис сделал вывод: чем раньше данный вид попал с материка на о. Цейлон, т. е. чем больше возраст вида, тем больше шансов ему распространиться. Именно "старые виды" широко распространены на о. Цейлон. Эндемики о. Цейлон — это наиболее молодые виды. Подобная картина была типична для любого рода, включающего 10–15 видов, т. е. там, где можно было провести полуколичественную оценку. Картина повторилась при сопоставлении распределения видов двух соседствующих больших островов Новой Зеландии и отстоящих от них на разное расстояние островков того же архипелага.
Роды, которые наиболее богатые видами — это наиболее старые по возрасту в том или в другом семействе Число видов в роде коррелирует также с характером их географического распределения в такой степени, что позволяет делать предсказание. Дав краткий обзор данных Дж. Виллиса, Г. де Фриз вслед за Дж. Виллисом делает следующий вывод (Vries H., 1923):
"На основе этих статистических результатов теория происхождения видов путем естественного отбора неограниченных вариаций может быть серьезно оспорена. Как правило, скорость распространения новых видов независима от их особой организации. Эта скорость одинакова для всех видов. В большинстве своем виды-эндемики возникли вовсе не в ходе адаптации к местным условиям... Систематические признаки, которые отделяют один вид от другого, не показывают какой-либо регулярной связи с борьбой за существование ".
Привожу важное феноменологическое обобщение Дж. Виллиса: "Ареал, занимаемый на данное время в данной стране какой-либо группой из родственных видов числом не менее 10, если условия относительно постоянны, зависит в основном от возраста видов этой группы в этой стране; общая картина может существенно меняться под действием природных барьеров: морей, рек, гор, изменений климата, влияния разных экологических факторов, включая человека и других причин" (Willis, 1922).
Другая эволюционная закономерность, установленная Дж. Виллисом — распределение числа видов в роде и числа родов в семействе соответствует так называемой "логарифмической кривой" (см.; обсуждение в книге Ю. В. Чайковского, 1991). Закономерность не утрачивает своей строгости, если оговариваются условия, при которых она проявляется. Например, менделевское расщепление 3:1 в реальности очень часто искажается, ибо для своего выполнения требует целого ряда условий: чистота гамет, полное проявление признака, наконец, необходимо изучить достаточное число потомков при расщеплении.
По удивительному, однако, не столь уж редкому в истории науки совпадению, две книги с серьезной критикой ведущей роли естественного отбора в видообразовании и эволюции и, напротив, выявляющие закономерные, номогенетические стороны эволюции — "Номогенез" Л. С. Берга и "Возраст и Ареал" Дж. Виллиса вышли в один и тот же год. И уже в английском издании "Номогенеза", опубликованном в 1926 г., Л. С. Берг в ряде мест цитирует Дж. Виллиса. Он приводит заключительные слова из помещенной в книге Дж. Виллиса статьи Г. де Фриза: "Привычную фразу, что виды приспосабливаются к окружающим условиям, надо, следовательно, понимать наоборот, а именно, что большинство из имеющихся сейчас видов живут в условиях, наиболее полезных для них... Выражаясь более популярным языком, можно сказать, что, в конечном итоге, виды выбирают наилучшую среду обитания. Благоприятные местные условия вызывают быстрое размножение, тогда как в других местах эти формы остаются редкими или медленно исчезают" (Берг Л. С., 1977, с. 178).
2.2.5. Номогенетические аспекты закона гомологических рядов
Закон гомологических рядов в наследственной изменчивости, сформулированный Н. И. Вавиловым (1968), включает два феноменологических обобщения:
"1. Виды и роды генетически близкие, характеризуются сходными рядами наследственной изменчивости с такой правильностью, что зная ряд форм в пределах одного вида, можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе генетически расположены в общей системе роды и линнеоны, тем полнее сходство в рядах их изменчивости.
2. Целые семейства растений в общем характеризуются определенным циклом изменчивости, проходящей через все роды и виды".
В соответствии с этим законом Н. И. Вавилов предсказал существование безлигульной формы у ржи. Такие формы не удалось найти только у ячменя, хотя на основании закона их следовало ожидать и у других растений. И действительно, А. Н. Луткову удалось получить путем облучения безлигульную мутацию ячменя.
Группы видов и роды отличаются в основном немногими признаками — "радикалами", а изменчивость других признаков сходна. Сходство изменчивости может базироваться не только на сходстве генов, но и на ограниченности формообразования. Иными словами, изменчивость вовсе не хаотична, а имеет определенное русло и может быть у разных видов сходна, несмотря на различие генов.
На основании выделенных видовых, родовых и других радикалов Н. И. Вавилов предложил перестроить систематику: "Важнейшей задачей систематики является выработка единой системы, в основу которой должны быть поставлены специфические различия видов и родов, их радикалов наряду с учетом гомологических рядов изменчивости в пределах видов". Например, для пшениц таким видовым радикалом является прежде всего кратное семи число хромосом: 14, 28 и 42 (однозернянки, твердые и мягкие пшеницы). Очевидно, что такой радикал, как кратное изменение в числе хромосом, не мог возникнуть постепенно, путем отбора, Переход происходил сразу, путем хромосомных и геномных мутаций с одного числа хромосом на другое.
Использование диагностических признаков-радикалов для построения системы классификации организмов, не зависимой от их происхождения, филогении, представляло собой, по существу, отрицание дарвиновского принципа дивергенции. Последователь. Дарвина, Э. Геккель — предложил, исходя из принципа дивергенции, строить естественную систему в форме генеалогического древа. Но если существует параллелизм, гомология, то тем самым существенно ограничивается принцип дивергенции, а сходство организмов более не может служить надежным показателем их родства.
Исходя из закона Н. И. Вавилова, получали отнюдь не адаптивное толкование многие случаи мимикрии, подражание одних видов другим в форме, окраске и т. д. Это сходство может иметь адаптивный характер, но вовсе не есть результат отбора, а представляет собой, согласно Н. И. Вавилову, " во многих случаях повторение циклов изменчивости в различных семействах и родах". Предсказание Н. И. Вавилова замечательно подтвердилось, когда впоследствии среди насекомых было обнаружено много случаев так называемого "псевдомиметизма", когда представители одного семейства "подражают" представителям многих других семейств. Часто приводимые случаи мимикрии у бабочек отнюдь не всегда носят приспособительный характер, ибо оказалось, что модель и имитатор обитают в разных местах. Подобные факты собраны в книге Л. С. Берга (1977).
Естественный отбор не создает мимикрию шаг за шагом, а отбирает те или иные закономерно возникающие формы. Н. И. Вавилов рассматривает случай поразительного сходства плоскосемянной вики, засоряющей посевы чечевицы. Семена вики — сорняка настолько сходны по окраске и размерам с семенами чечевицы, что не могут быть разделены никакой сортировальной машиной. Оба вида одновременно цветут и созревают. Обнаруженное сходство, очевидно, возникло за счет бессознательного отбора со стороны человека, когда при помощи сортировальных машин отделялись формы вик, близкие к тарелочной чечевице по размеру и форме семян. Но роль отбора здесь подчиненная. Сравнительно-географический анализ показал большое сходство двух родов в отношении изменчивости семян, что в целом, согласно Н. И. Вавилову, "является прекрасной иллюстрацией гомологических рядов" и "обусловливается общими правильностями формообразования".
По сравнению с известными Дарвину фактами "аналогичных изменений" у близких видов, фактов, которые Ч. Дарвин считал несущественными, закон Н. И. Вавилова приобрел универсальность для теории эволюции. Ибо а) он распространялся не только на близкие виды, но и на разные роды одного семейства; б) постулировалась возможность предсказывать характер изменчивости в близких родах; в) сходство морфологических признаков у близких и далеких видов получало истолкование не только историческое или филогенетическое (как результат общности происхождения и обладания общими генами), но и номотетическое, основанное на общих "правильностях формообразования" или на параллелизме действия отбора на несходные генотипы.
Неудивительно, что, придавая большое значение "общим правилам формообразования", Н. И. Вавилов с интересом отнесся к идеям Л. С. Берга, найдя в книге "Номогенез" "массу любопытного... По уклону мысли мы во многом сходимся с Львом Семеновичем" (Вавилов, 1980, с. 56). Большое влияние концепция Н. И. Вавилова оказала на А. А. Заварзина и его учение о параллелизмах клеточных и тканевых структур, разработанное в 1920–1930-е годы.
Номотетический аспект закона Н. И. Вавилова оригинально разработан в концепции типологии С. В. Мейена (1978, 1980). Помимо понятия "архетип", введенного в эволюционную морфологию в XIX в. Оуэном (см.; Канаев И. И., 1966), С. В. Мейен вводит понятия мерона и рефрена. Мерон соответствует разным внутренним частям организма или классу частей (напр.; "конечность"). Мерономия — учение о меронах — должна войти, по мысли Мейена, в теорию морфологии организмов. Важной закономерностью мерономии являются гомологические ряды Н. И. Вавилова или, по терминологии С. В. Мейена, "повторяющиеся полиморфические множества или рефрены".
Изменение в терминологии связано с тем, что сходный характер полиморфизма, сходный рефрен может затрагивать и не гомологичные части или органы. Например, в разных семействах высших растений повторяются простые, пальчатые и перистые листья. Но важно не только то, что от рода к роду повторяются эти три типа, но гомологичен и способ преобразований одного типа в два других в пределах разных родов. "Одни и те же закономерности морфологического преобразования прослеживаются на органах самого разного происхождения, совершенно различной функции". Повторность типов преобразования настолько велика, что рефрены вполне могут являться основанием для прогноза, т. е. для экстраполяции полиморфического множества в тех случаях, когда известна какая-то его одна часть (Мейен, 1980).
– Конец работы –
Эта тема принадлежит разделу:
ВЕК ГЕНЕТИКИ: ЭВОЛЮЦИЯ ИДЕЙ И ПОНЯТИЙ
На сайте allrefs.net читайте: "ВЕК ГЕНЕТИКИ: ЭВОЛЮЦИЯ ИДЕЙ И ПОНЯТИЙ"
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Номогенетический аспект законов Менделя
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.029 сек.
Новости и инфо для студентов