Два компонента генотипа — две формы наследственной изменчивости. Мутации и вариации
Естественное разделение наследственной системы эукариот на две подсистемы — ОК и ФК приводит к расширенному представлению о формах наследственной изменчивости. С мутациями в общепринятом смысле связана лишь часть наследственной изменчивости, т. е. изменения, которые непосредственно затрагивают ОК.
В первой главе уже отмечалось, что в хромосомной теории наследственности принимались "на веру" два постулата:
а) весь генетический материал хромосом состоит из генов, вся ДНК имеет информативную функцию;
б) геном как перечень генов с их аллелями тождественен понятию генотип.
Выясняется, что оба неявных постулата неточны и ведут к неверным эволюционным следствиям (А. Е.Акифьев, 1995).
М. Е. Лобашов (1976), исходивший при изучении наследственной изменчивости из физиологии клетки, предупреждал: "Наши генетические методы позволяют улавливать лишь изменения, связанные с постоянными элементами ядра и клетки. В силу этого мы обращаем внимание на изменения в хромосоме и упускаем другие". Говоря обыденным языком, исследователи нередко искали ключ не там, где он потерялся, а под фонарем, потому что там светло.
Под термином мутация в более узком сложившемся в классической генетике смысле целесообразно понимать лишь те изменения, которые прямо или косвенно затрагивают постоянные облигатные компоненты генотипа (ОК). Для обозначения самых разных изменений ФК был предложен термин вариация (Голубовский М. Д., 1985), который так или иначе уже использовался в истории генетики применительно к изменчивости (Ю. А. Филипченко, 1929). Небольшое переосмысление термина — обычная ситуация в науке.
Поистине, изучая структурную организацию генотипа у разных особей одного вида, мы постоянно сталкиваемся с вариациями на тему видового генома.
Рассмотрим: 1) характер изменений, связанных с факультативными изменениями; 2) их генетическую основу (т. е. какие именно факультативные элементы меняются и каким образом); 3) индуцирующие факторы, вызывающие активацию ФК.
Наследственные изменения, которые вызваны ФК, могут затрагивать существенные характеристики клетки и организма — от возникновения генных мутаций и до дестабилизации генома.
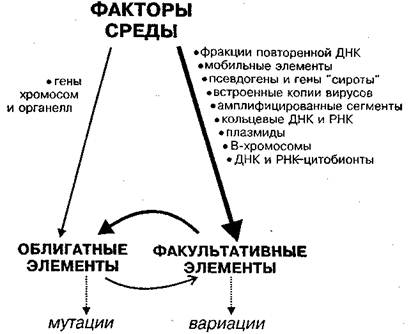
Рис. 2. Пути возникновения наследственных изменений в природе в системе СРЕДА — ФАКУЛЬТАТИВНЫЕ — ОБЛИГАТНЫЕ ЭЛЕМЕНТЫ
Справа указаны факультативные элементы, слева — облигатные элементы генома. Изменения первых обозначены как вариации, изменение вторых — мутации. Толщина линий соответствует интенсивности связи. Факультативные элементы первыми воспринимают немутагенные изменения среды, а затем возникающие вариации вызывают мутации. Поведение факультативных элементов зависит также от облигатных генов.
Мутации, согласно классической генетике, возникают случайно, с малой частотой (примерно, 1х10–6 на ген за поколение), у отдельных особей. Характер наследственных вариаций, обусловленных факультативными элементами, совсем иной: здесь возможны массовые, упорядоченные наследственные изменения. Кроме того, вариации могут возникать при таких изменениях абиотических, биотических или генетических факторов среды, при которых канонические изменения возникают редко. На рис. 2 представлена общая схема взаимодействий в триаде "Среда — ФК — ОК".
Ю. Я. Керкис (1940), полемизируя с Н. В. Тимофеевым-Ресовским, впервые пришел к правильному выводу, что спонтанные наследственные изменения возникают за счет нарушения внутриклеточного метаболизма и физиологического гомеостаза. Именно эти причины в первую очередь индуцируют изменения факультативных элементов, которые затем опосредованно, по принципу усиления, вызывают мутации ОК. Вариации возникают под действием самых обычных немутационных факторов.
Остановимся лишь на одном примере того, как возникают определенные, массовые наследственные изменения, опосредованные ФК в случае феномена гибридного дисгенеза у дрозофилы. При синдроме гибридного дисгенеза в потомстве некоторых скрещиваний возникают вспышки мутаций (генных и хромосомных), резко изменяется характер рекомбинации у самцов, появляется частичная или полная стерильность. Суть этого феномена состоит в активации тех или иных мобильных элементов при разных скрещиваниях. Так, в случае Р-М системы гибридного дисгенеза Р-элементы, содержащиеся в хромосомах линий Р-цитотипа, попадая в цитоплазму М-цитотипа (где нет репрессоров транспозиции), начинают массово перемещаться. Это сопровождается появлением нестабильных инсерционных мутаций и разрывом хромосом именно в тех районах, где локализованы Р-транспозоны данной линии (Engels, 1983; Kid–well, 1985; Lim, Simmons, 1994).
На фенотипическом уровне эти массовые вариации выражаются в появлении частичной или полной стерильности у самцов первого поколения. Частота возникновения хромосомных перестроек в дисгенных линиях может достигать фантастических величин — до 10% (Engels, 1983).
Не всегда массовые транспозиции имеют негативный эффект. Так, при переходе от глубокого инбридинга к свободному скрещиванию в селектированных Л. 3. Кайдановым линиях дрозофил за одно-два поколения происходят множественные кооперативные миграции МГЭ и одновременно резко возрастает жизнеспособность линий (Гвоздев В. А., Кайданов Л. 3., 1986, 1990).
МакКлинток (McClintock, 1978) пришла к выводу, что способность к активации факультативных элементов и реорганизации генома представляет собой род адапгивного ответа клетки на стресс. Степень реорганизации варьирует от амплификации генов, перемещения МГЭ, количественных изменений и транспозиций блоков гетерохроматина до сложных инверсий и транслокаций. Множественные вариационные и мутационные изменения способны обеспечить достижение нового адаптивного пика при стрессовых воздействиях (Кайданов и др., 1994).