ХРОМОСОМНАЯ ТЕОРИЯ НАСЛЕДСТВЕННОСТИ
Сцепленное наследование признаков. Как мы отмечали в прошлой лекции, независимое наследование признаков при ди- и полигибридном скрещивании бывает в случае, если гены этих признаков локализованы в разных хромосомах. Но количество хромосом ограничено по сравнению с количеством признаков. У большинства животных организмов число хромосом не превышает 100. В то же время число признаков, каждый из которых контролируется по крайней мере одним геном, значительно больше. Так, например, у дрозофилы изучено 1000 генов, которые локализованы в четырех парах хромосом, у человека известно несколько тысяч генов при 23 парах хромосом и т.д. Отсюда следует, что в каждой паре хромосом располагается много генов. Естественно, что между генами, которые находятся в одной хромосоме, наблюдается сцепление, и при образовании половых клеток они должны передаваться вместе.
Сцепленное наследование признаков открыли в 1906 г, английские генетики В.Бетсон и Р.Пеннет при изучении наследования признаков у душистого горошка, но они не смогли дать теоретическое объяснение этому явлению. Природу сцепленного наследования выяснили американские исследователи Т. Морган и его сотрудники С. Бриджес и А. Стертевант в 1910 году. В качестве объекта исследований они избрали плодовую мушку дрозофилу очень удобную для генетических опытов. Достоинства этого объекта исследования следующие: малое число хромосом (4 лары), высокая плодовитость, быстрая сменяемость поколений ( 12-14 суток). Мухи дрозофилы серого цвета, с красными глазами, имеют маленькие размеры (около 3 мм), легко разводятся в лабораторных условиях на простых по составу питательных средах. У дрозофилы выявлено большое число мутантных форм. Мутации затрагивают окраску глаз и тела, форму и размер крыльев, расположение щетинок и др.
Изучение наследования разных пар признаков и их расщепления при дигибридномскрещивании позволило обнаружить наряду с независимым комбинированием признаков явление сцепленного наследования. На основании изучения большого числа признаков было установлено, что все они распределяются на четыре группы сцепления в соответствии с числом хромосом у дрозофилы. Сцепленное наследование признаков связано с локализацией группы определенных генов в одной хромосоме.
Мысль о локализации генов в хромосомах была высказана Сеттоном еще в 1902 году, когда им был обнаружен параллелизм в поведении хромосом в мейозе и наследовании признаков у кузнечика.
Наиболее четкая разница в поведении сцепленных и независимо наследующихся генов выявляется при проведении анализирующего скрещивания.
Рассмотрим это на примере. В первом случае возьмем признаки, гены которых расположены в разных хромосомах.
А В а в
Р === === х === ===
а в а в
Гаметы: АВ, Ав,аВ,авав
А В А в а В а в
F === === ; === === ; === === ; === ===
а в а в а в а в
В результате мы получили потомство четырех фнотипических классов в соотношении: 1 : 1 : 1 : 1. Другие результаты будут, если гены А и В локализованы в одной хромосоме.
А В а в
Р =*===*= х =*===*=
а в а в
Гаметы: А В,а ва в
А В а в
F =*===*= ; =*===*=
а в а в
Таким образом, если гены находятся в одной хромосоме в потомстве при анализирующем скрещивании, мы получим два класса потомков похожих на отца и на мать и не будет потомков с признаками отца и матери одновременно.
Опыты, подтверждающие сцепленное наследование признаков, были проведены Т.Морганом на дрозофиле. Для скрещивания были взяты особи серые с нормальными крыльями (доминантные признаки) и черные с зачаточными крыльями (рецессивные признаки). В результате опытов были получены потомки только серые крылатые и черные с зачаточными крыльями.
На основании проведенных экспериментов Т.Морган сформулировал закон сцепленного наследования признаков: признаки, гены которых располагаются в одной хромосоме, наследуются сцепленно.
Неполное сцепление. Явление кроссинговера.Наряду с полным сцепленным наследованием признаков Т.Морган в своих опытах с дрозофилой обнаружил и неполное сцепленное наследование. При неполном сцепленном наследовании одновременно с формами, похожими на родителей, были обнаружены организмы, у которых наблюдались признаки обоих родителей. Однако соотношение этих форм не было равным как при независимом комбинировании.В потомстве явно преобладали формы, схожие с родителями, а организмов рекомбинантов было значительно меньше.
Схема неполного сцепленного наследования признаков.
А В а в
Р =*===*= х =*===*=
а в а в
Гаметы: А В,а в,а В,А ва в
без кроссин. кроссоверные
А В а в а В А в
F ====; ====; ====; ====
а в а в а в а в
рекомбинанты
Объяснить этот факт можно следующим образом. Если гены А и В расположены в одной хромосоме, а в гомологичной ей хромосоме расположены рецессивные аллели а и в, то отделиться друг от друга и вступить в новые сочетания гены А и В могут только в том случае, если хромосома, в которой они расположены, будет разорвана на участке между этими генами и затем соединена с участком гомологичной хромосомы. В 1909 году Ф. Янсенс, изучая мейоз у земноводных, обнаружил в диплотене профазы 1 хиазмы (перекресты хромосом) и высказал предположение, что хромосомы взаимно обмениваются участками. Т.Морган развил это представление в идею об обмене генами приконьюгации гомологичных хромосом, а неполное сцепление было объяснено им как результат такого обмена и названо кроссинговером.
Схема кроссинговера.
 |  | 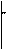 |  |  | | ||||||
А а А а А а
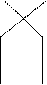 |
В в в В в В
Кроссинговер может быть одинарным, как показано на схеме, двойным и множественным. Кроссинговер возник в процессе эволюции. Он приводит к появлению организмов с новыми сочетаниями признаков, т.е. к увеличению изменчивости. Изменчивостьже является одним из движущих факторов эволюции.
Частота кроссинговера определяется по формуле и выражается в процентах или морганидах ( 1 морганида равна 1% перекреста ).
число рекомбинантов
 Р кроссинговера = х 100%
Р кроссинговера = х 100%
общее число потомков
Если, например, общее число потомков, полученное в результате анализирующего скрещивания, равно 800, а число кроссоверных форм – 80, то
частота кроссинговера будет:
 Р кросс. = х 100% = 10% ( или 10 морганид)
Р кросс. = х 100% = 10% ( или 10 морганид)
Величина перекреста зависит от расстояния между генами. Чем дальше удалены гены друг от друга, тем чаще происходит перекрест. Установлено, что количество кроссоверных особей к общему числу потомков никогда не превышает 50%, так как при очень больших расстояниях между генами чаше происходит двойной кроссинговер и часть кроссоверных особей остается неучтенной.
Явление кроссинговера, установленное генетическими методами на дрозофиле, нужно было доказать цитологически. Это сделали в начале 30 годов Штерн на дрозофиле и Б. Мак-Клинтон на кукурузе. Для этого были получены гетероморфные хромосомы, т.е. хромосомы, различающиеся внешне с локализацией в них известных генов. В этом случае у кроссоверных форм можно было видеть рекомбинантные хромосомы и сомнений о наличии кроссинговера не возникало.
Процесс протекания кроссинговера зависит от многих факторов. Большое влияние на кроссинговер оказывает пол. Так, у дрозофилы кроссинговер происходит только у самок. У тутового шелкопряда кроссинговер отмечается у самцов. У животных и человека кроссингавер происходит у обоих полов. На частоту кроссинговера влияют также возраст организмов и условия среды.
К. Штерн показал, что кроссинговер может возникать не только в мейозе, при развитии половых клеток, но в некоторых случаях и в обычных соматических клетках. П о-видимому соматический кроссинговер широко распостранен в природе.
Линейное расположение генов в хромосомах. Карты хромосом. После того как была установлена связь генов с хромосомами и обнаружено, что частота кроссинговера всегда вполне определенная величина для каждой пары генов, расположенных в одной группе сцепления, встал вопрос о пространственном расположении генов в хромосомах. На основании многочисленных генетических исследований Морган и его ученик Стертевант выдвинули гипотезу линейного расположения генов в хромосоме. Изучение взаимоотношения между тремя генами при неполном сцеплении показало, что частота перекреста между первым и вторым, вторым и третьим, первым и третьим генами равна сумме или разности между ними. Так, если в одной группе сцепления расположены три гена - А, В и С, то процент перекреста между генами АС равен сумме процентов перекреста между генами АВ и ВС, частота перекреста между генами АВ оказалась равной АС - ВС, а между генами ВС = АС - АВ. Приведенные данные соответствуют геометрической закономерности в расстояниях между тремя точками на прямой. На этом основании был сделан вывод, что гены расположены в хромосомах в линейной последовательности на определенном расстоянии друг от друга. Используя эту закономерность, можно строить карты хромосом.
Карта хромосомы это схема, на которой показано, какие гены локализованы в данной хромосоме, в каком поряке и на каком расстоянии друг от друга они располагаются. Для построения карты хромосом проводят анализирующее скрещивание и определяют частоту кроссинговера. Например, установлено, что в хромосоме локализованы три гена М, N и К. Частота перекреста между генами М и N составляет 12%, между М и К - 4 % и между N и К - 8%. Чем больше частота кроссинговера, тем дальше друг от друга расположены гены. Используя эту закономерность, строим карту хромосомы.
М 4% К 8% N
 о о о
о о о
После построения генетических карт встал вопрос о том, отвечает ли расположение генов в хромосоме, определенное на основании частоты кроссинговера, истинному расположению. С этой цепью генетические карты нужно было сравнить с цитологическими.
В 30 годах нашего столетия Пайнтер открыл в слюнных железах дрозофилы гигантские хромосомы, строение которых можно было изучать под микроскопом. Хромосомы эти имеют характерный для них поперечный рисунок в виде дисков разной толщины и формы. Каждая хромосома по длине имеет специфические рисунки дисков, что позволяет отличать разные ее участки друг от друга. Появилась возможность сравнить генетические карты с фактическим расположением генов в хромосомах. Материалом для проверки служили хромосомы, у которых вследствие мутаций возникли различные хромосомные перестройки:не хватало отдельных дисков, или они были удвоены. Диски служили маркерами, с их помощью определяли характер хромосомных перестроек и место расположения генов, о существовании которых было известно на основании данных генетического анализа. При сопоставлении генетических карт хромосом с цитологическими было установлено, что каждый ген находится в определенном месте (локусе) хромосомы и что гены в хромосомах расположены в определенной линейной последовательности. В то же время было обнаружено, что физические расстояния между генами на генетическойкарте не вполне соответствуют установленным цитологически. Однако это не снижает ценности генетических карт хромосом для предсказания появления особей с новыми сочетаниями признаков.
На основании анализа результатов многочисленных исследований на дрозофиле и других объектах Т. Морган сформулировал хромосомную теорию наследственности, сущность которой заключается в следующем:
- материальные носители наследственности - гены находятся в хромосомах, располагаются в них линейно на определенном расстоянии друг от друга;
- гены, расположенные в одной хромосоме, относятся к одной группе
сцепления.Число групп сцепления соответствуют гаплоидному числу хромосом;
- признаки, гены которых находятся в одной хромосоме, наследуются сцепленно;
- неполное сцепленное наследование признаков связано с явлением кроссинговера, частота которого зависит от расстояния между генами;
- на основании линейного расположения генов в хромосоме и частоты кроссинговера как показателя расстояния между генами можно построить карты хромосом.
Лекция 5