рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Математика
- /
- Принцип дифференциальной активности генов как основа гетерогенизации (регионализации) развивающегося организма
Реферат Курсовая Конспект
Принцип дифференциальной активности генов как основа гетерогенизации (регионализации) развивающегося организма
Принцип дифференциальной активности генов как основа гетерогенизации (регионализации) развивающегося организма - раздел Математика, Наш Организм Построен Из Большого Количества Клеток, Которые Организуются В К...
Наш организм построен из большого количества клеток, которые организуются в клеточные ассоциаты, ткани и органы. Количество существующих во взрослом организме клеточных типов также очень велико. Среди одних только форменных элементов крови встречаются разные виды лейкоцитов, лимфоциты, эритроциты. Принадлежность клеток к тому или иному типу, к той или иной ткани дает себе знать и в условиях культивирования тканей вне организма, где каждая тканевая система обнаруживает качественно специфичные, только ей одной присущие особенности поведения. Встает естественный вопрос: Как в условиях идентичности наследственного аппарата, при одинаковом наборе генов во всех клетках, возникает это многообразие клеток и тканей? На этот вопрос помогает ответить генетика развития. Генетика развития изучает реализацию наследственной информации в ходе онтогенеза, т.е. путь от гена к признаку. Это направление заложили в 1920-е годы исследования немецкого зоолога В. Хеккера, который назвал его феногенетикой. У истоков феногенетики стояли такие выдающиеся ученые, как Т. Морган, Р. Гольдшмидт, Э. Хадорн, С. Уодингтон, Н. К. Кольцов.
Основной вопрос генетики развития был сформулирован еще Т. Морганом: каким образом молекулярно-генетические события в ходе онтогенеза детерминируют формообразовательные процессы? Современная генетика развития сложилась на стыке четырех дисциплин – генетики, эмбриологии, биохимии и молекулярной биологии. Становление феногенетики может быть условно подразделено на несколько этапов.
Первый этап – описательный. Он приходится на начало 20-х и отчасти на 30-е годы ХХ в. Началось с того, что немецкий зоолог В. Хеккер заинтересовался тем, когда можно обнаружить различия в развитии нормальных и мутантных зародышей млекопитающих. Для этой цели он готовил гистологические срезы нормальных эмбрионов и эмбрионов-носителей той или иной мутации на разных стадиях их развития и определял стадию эмбриогенеза, на которой гистологические картины тех и других начинают отличаться. Такие стадии Хеккер назвал фенокритическими фазами. В течении 20-х – 30-х годов был накоплен огромный материал в этой области. В особенности на млекопитающих – крысах, мышах, морских свинках. А норвежский цитолог и генетик К. Боннерви создала базу для развития современного учения о наследственных нарушениях онтогенеза у человека, и это дало возможность осмыслить данные по врожденным порокам развития у человека.
Фенокритические фазы развития изучались не только у животных, но и у растений, где активно работали лаборатории Э. Бауера в Германии и Е. Синнота в США. Но для более глубокой оценки и интерпретации описательного материала требовались серьезные экспериментальные исследования, которые были проведены в течении 30-х годов ХХ в. и составили суть второго этапа.
Второй этап – экспериментальный. Он включает 30-е годы и начало 40-х годов ХХ в. В этот период, благодаря усилиям Т. Моргана, Р. Гольдшмидта, швейцарского зоолога Э. Хадорна, английского генетика С. Уоддтингтона, российской школы Н. Кольцова удалось накопить богатый экспериментальный материал, проливающий свет на некоторые закономерности генетического контроля индивидуального развития. Именно в этот период были сформулированы основные принципы феногенетики:
I. Принцип дифференциальной активности генов как основа гетерогенизации (регионализации) развивающегося организма. Вопрос о закономерностях функционирования генома возник очень рано. Еще А. Вейсман в конце ХIХ в. пытался построить стройную схему, с помощью которых удалось бы их объяснить. Он полагал, что в процессе индивидуального развития следует различать два типа клеточных делений – равнонаследственное и неравнонаследственное. При втором типе делений наследственное вещество распределяется по дочерним клеткам неравномерно, и именно это создает различия между ними и лежит в основе гетерогенизации зародыша. Согласно Вейсману, существует специфическая группа органических молекул, названных им биофорами, которые находятся в ядре клеток, и через поры ядра мигрируют в цитоплазму. Ядру, таким образом принадлежит активная роль, а цитоплазма, ее поведение и дифференцировка, как в целом структура и функции клеток, зависят от специфических групп биофор – детерминантов. В зародышевой плазме находится полный набор детерминантов: их столько, сколько типов однородных клеток имеется у взрослого организма. Детерминанты как и биофоры растут и размножаются (делятся), так что для любого числа однородных клеток определенной ткани достаточно одного исходного детерминанта. Все детерминанты, определяющие развитие различных признаков организма, сгруппированы в ядре в так называемые иды. Во время митоза иды слагаются в иданты, соответствующие хромосомам.
Следовательно, по Вейсману, возникающие в ходе развития организма различия между клетками обуславливаются сортировкой наследственных единиц (детерминантов). Эти единицы распределяются неравномерно по различным клеткам и детерминируют их специализацию. Только половые клетки имеют полный набор детерминантов, а потому оказываются способными развиваться в целый организм. Так родилась теория зародышевого пути, согласно которой уже в ходе первого деления дробления одни клетки, где сохраняется полный набор детерминантов, образуют зародышевый путь, другие клетки, где детерминанты специфически распределяются между различными соматическими клетками, образуют соматический путь.
Обычно тончайшая структура хромосом в соматических клетках не претерпевает существенных изменений и, следовательно, генотип всех клеток тела одинаков. Значит основная проблема генетики развития, изучающей действие генов в онтогенезе, заключается в выяснении того, каким образом при идентичном наборе генов во всех клетках организма формируются клеточное разнообразие и морфофункциональная специализация тканей и органов. На этот счет, начиная с 20-30-х годов ХХ в., существует две «модели» (или гипотезы) объяснения феномена.
Первая гипотеза была сформулирована Морганом, который полагал, что несмотря на одинаковый набор генов, в клетках многоклеточного организма, расположенных в разных частях развивающегося зародыша, и в разные моменты их дифференцировки функционируют разные гены, потому-то они и приобретают сначала химическое, а затем и морфологическое своеобразие.
Вторую гипотезу выдвинул Гольдшмидт. Он предположил, что во всех клетках одинаково работают все гены, но их продукты испытывают разную судьбу в разных частях зародыша. Именно там они подвергаются селективному отбору, так что наблюдается не дифференциальная активность генов в разных клетках, а дифференциальное функционирование их продуктов.
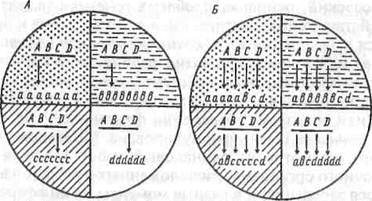
Схема, иллюстрирующая гипотезы Т.Моргана и Р.Гольдшмидта.
A—D — гены; а. b, с, d— геноконтролируе-мые продукты.
II. Принцип ведущей роли ядерно-цитоплазматических отношений в регионализации зародыша. Этот принцип феногенетики был сформулирован в 30-е годы.… Ведущая роль ядра в регуляции формообразования подтверждается рядом опытов,… В ранних эксперементально-эмбриологических исследованиях удалось выяснить, что по крайней мере в ходе начальных стадий…Рис. 5. Детерминация анимально-вегетативного градиента в яйце дрозофилы посредством действия трех независимых генных систем. Локализация продуктов соответствующих генов в яйце.
Вторая система генов контролирует формирование дорсо-вентрального градиента, например ген toll, при отсутствии которого эмбрион сильно «дорсализован». Третья система генов контролирует формирование градиентов в оогенезе. Эта группа генов контролирует формирование терминальных структур, т.е. акрона (несегментированного головного конца) и тельсона (несегментированного хвостового конца). Ключевую роль в этом процессе играет ген torso. При отсутствии его продукта ни акрон, ни тельсон не развиваются, так что эмбрион оказывается полностью сегментированным.
Весь процесс индивидуального развития осуществляется на основе двух типов воздействия генов друг на друга: активирующего и тормозящего. Регионализации эмбриона, спецификация его клеток, их взаимовлияния в ходе морфогенеза основаны на «игре» этих факторов и установлении некоего баланса таких антиномических состояний, разных в различных областях развивающегося зародыша. Итогом этого тонко сбалансированного процесса является неравномерное распределение генопродуктов вдоль эмбриональных осей, так что создается система полярных градиентов распределения биологически активных веществ, своеобразная химическая мозаика, химически переформированный «план строения» организма, воплощаемый в жизнь в ходе онтогенеза.
Решающую роль в последующих этапах детерминации играют регуляторные гены. Формирование специфичного для данной ткани набора активных регуляторных генов и составляет суть клеточной детерминации.
В реализации плана строения организма ключевое значение имеет его сегментация, разделение на головной, грудной, брюшной отделы и их производные. Этот процесс является универсальным в животном мире и характеризуется двумя основными признаками – количеством сегментов и их качеством. Соответственно различают две группы генов, ответственных за развитие этих признаков: сегрегационные и гомеозисные. Сегрегационных генов, детерминирующих число сегментов у дрозофилы, известно более двух десятков. Их мутации нарушают становление переднезадней (анимально-вегетативной) полярности сегментов, в результате чего происходит их слияние, уменьшается количество и образуются не жизнеспособные уроды. За открытие и изучение этих генов американский генетик Э.Льюис и ряд других ученых были удостоены Нобелевской премии.
Различают несколько групп сегрегационных генов. Это гены материнского эффекта, которые контролируют формирование градиентов в ходе оогенеза, GAP-гены, pair-rule-гены, гены сегментарной полярности, последовательно осуществляющие сегментацию зародыша и подготавливающие почву для функционирования гомеозисных генов.
Сегрегационные гены последовательно активируются в процессе индивидуального развития. GAP-гены начинают функционировать на синцитиальной стадии развития, когда к 10-11-му циклу клеточного деления ядра мигрируют к периферии развивающегося эмбриона и «прочитывают» позиционную информацию, возникшую благодаря активности генов материнского организма, формируя на поверхности яйца синцитий. В результате этого процесса зародыш подразделяется на несколько пространственных доменов.
На фоне специфического распределения продуктов gap-генов под их влиянием активируются Paire-rule-гены, которые дробят зародыш на повторяющиеся домены шириной по два парасегмента, в одном из которых этот ген активен, в другом нет. Уже к 14-му циклу дробления активность этих генов концентрируется в определенных ядрах. В конце концов, благодаря функционированию генов Pair-rule зародыш подразделяется на отдельные сегменты. Нарушение их функционирования ведет к выпадению отдельных (чередующихся) сегментов. Кроме того, они имеют некоторое значение и в формировании головных структур. Различные сочетания белковых продуктов генов Pair-rule активируют гены сегментарной полярности. Эти гены детерминируют границы конкретных сегментов. Мутации, нарушающие их функционирование, влекут за собой нарушения в развитии отдельных сегментов – четных или нечетных, в зависимости от того, какой из генов сегментарной полярности мутировал. Экспрессия сегрегационных генов подготавливает «почву» для функционирования ключевой системы генов, обеспечивающих качественную спецификацию сегментов, - системы гомеозисных генов.

Рис.6. Последовательная активация сегрегационных генов в онтогенезе.
А — каскад генов, осуществляющих процесс последовательного разделения тела развивающегося эмбриона дрозофилы на сегменты.Б — транскрипты гена fushi tarazu локализованы в семи полосках соответственно семи парасегментам.
Название группы гомеозисных генов происходит от термина «гомеозис», который ввел в 1894 г. один из классиков генетики У.Бэтсон. под гомеозисом он понимал превращение одной части тела в другую. Гомеозисные гены, следовательно, не представляют собой нечто самостоятельное, но являются частью специфической системы генов, контролирующих сегментацию тела насекомых, в частности дрозофилы и других организмов. Примером гомеозисных мутаций является превращение антенны в ногу или аристы в ногу.
Гомеозисные гены, которых у дрозофилы описано около полусотни, контролируют качественные особенности сегментов и в свою очередь подразделяются на два комплекса: Antennapedia-complex и Bithorax-complex. Гены принадлежащие к ANT-C, контролируют развитие головных сегментов, в том числе интеркалярного, максиллярного, мандибуллярного, лабиального, а также грудных сегментов. При утрате функции гена Antp область тела, включающая заднюю часть первого грудного сегмента Т1, весь второй грудной сегмент Т2 и переднюю часть Т3, приобретают свойства головных сегментов, что проявляется в образовании головных структур в грудной области. Наоборот, в случае эктопической активности гена Antp, вызванной например, мутацией, которая нарушает нормальную для данной области тела репрессию данного гена, происходит образование грудных структур на голове. Очевидно ген Antp участвует в выборе программ развитияв направлении головы или груди. Гены комплекса BX-C ответственны за спецификацию грудных и брюшных сегментов. Этот комплекс состоит из трех «отделов»: Ultrabithorax, ответственного за развитие грудных сегментов, а также abdomen-A и аbdomen-B, контролирующих дифференцировку брюшных сегментов. Все они построены и функционируют по единому принципу. Пониманию функционального значения этих генов помогают их мутации. Например, мутация bithoraxoid дает нам такое проявление в фенотипе: первый брюшной сегмент развивается как заднегрудь и несет поэтому жужжальца и ноги. Муха, обладательница этой мутации, кроме дополнительных дефектных крыльев (всего их шесть) имеет восемь ног вместо обычных шести. Все гомеозисные гены имеют общие нуклеотидные последовательности которые получили название гомеобокс, а коллинеарный им фрагмент белковой молекулы – гомеодомен. Именно область гомеодомена ответственна за соединение регуляторного белка с ДНК. Гомеобокс был открыт Мак-Гиннисом в начале 1980-х годов.
У млекопитающих выявлено 38 гомеобокссодержащих генов. Они получили название HOX-генов. Как гомеозисные гены дрозофилы, так и HOX-гены могут формировать компактные группы – кластеры. У человека 4 кластера расположены на 2, 7, 12, 17 хромосомах. В отличии от дрозофилы, у млекопитающих определенная структура контролируется не отдельными HOX-генами, а специфичной для этой структуры совокупностью экспрессией нескольких HOX-генов.
Внимание к гомеозисным генам велико еще и потому, что они имеют прямое отношение к регуляции морфогенетических процессов. Морфогенез заключается в образовании компартментов. Что это такое? Дело в том, что клеточные клоны, потомки отдельных клеток-предшествениц, объединяются и образуют поликлон. Появляется целый ряд поликлонов, которые формируют морфогенетические зоны. Эти зоны отделены друг от друга четкими границами и обозначаются термином компартмент. Ф.Крик и П.Лоуренс предположили, что гомеозисные гены как раз и ответственны за образование компартментов и являются своеобразными генами-селекторами, поскольку именно они «выбирают» путь развития клетки. Этот путь определяется бинарными решениями – к примеру, стать ли клетке передней или задней, дорсальной или вентральной, грудной или крыловой.
Одним из интересных явлений, наблюдаемых в генетике развития, является феномен коллинеарности в расположении генов и контролируемых ими признаков. Этот удивительный феномен означает соответствие линейного расположения генов линейной последовательности распределения детерминируемых ими признаков в теле развивающегося зародыша. Признаки, которые контролируют гомиозисные гены (голова, грудь, брюхо), расположены точно в той же последовательности, что и соответствующие комплексы генов (ANT-C, BX-C, abd-A, Abd-B). Иными словами, в определенном участке хромосомы как бы «нарисована» в молекулярных терминах миниатюрная мушка-дрозофила или мышка, или человечек. Получается нечто похожее на то, о чем думали преформисты XVII в. Только они полагали, что в половой клетке присутствует миниатюрный человек в буквальном смысле этого слова, в действительности же его «образ записан», но в определенной структуре клетки, несущей наследственную информацию (хромосома), и на специфическом языке (молекулярно-генетическом).
Однако существуют регуляторные гены, контролирующие гомеозисные гены, так же как и гены, контролируемые ими. Дифференциальная экспрессия обеспечивается взаимодействием многих регуляторных генов. Активация одних генов запускает экспрессию других. Такая сложная морфогенетическая цепочка может быть инициирована одним геном, который «включает» комплекс последовательных формообразовательных событий. Гены, запускающие такой «каскад», получили в биологии развития название «гены-господа», а структурные гены – «гены-рабы». Между ними располагаются многочисленные регуляторные гены, к которым принадлежат и гомеозисные. Название этим генам предложил выдающийся шведский цитолог Я.Э.Эдстрем еще в начале 60-х годов ХХ в. - «Master Genes» («гены-господа») и «Slaves-Genes» («гены-рабы»).
В.Геринг проверил эти соображения эксперементально. Он использовал в своих опытах очень интересную модельную систему. С помощью микроинъекций генноинженерных конструкций получали два типа линий трансгенных дрозофил. В геном первого типа трансгенных дрозофил вводили так называемую GAL4-систему, где она попадала под разные геномные тканеспецифические энхансеры и, следовательно, активировалась в разных участках тела мухи. GAL4 – это дрожжевой активатор транскрипции. Его эффект распространяется лишь на те гены, перед которыми расположена так называемая последовательность UAS, содержащая пять GAL4-связывающих сайта. Эту последовательность вводили в геном второго типа трансгенных дрозофил. Если к ней «прицепить» еще и репортерный ген бактериальной галактозидазы, то с помощью окрашивания специальным реактивом можно обнаружить, где эта последовательность будет активирована в результате воздействия продукта GAL4. Геринг скрещивал линии этих двух типов между собой, причем к UAS-последовательности был присоединен ген eyeless, мутация которого вызывает отсутствие глаз у мухи. Легко заметить, что в такого рода опытах могут получаться сочетания, в которых ген ey будет через UAS-последовательность активироваться в эктопических местах. Это связано с тем, что GAL4 у разных линий находился под контролем разных энхансеров, в том числе и таких, которые функционировали в области крыльев, антенн, ног и т.д. В результате Герингу удалось получить мух, у которых глаза могли появиться в самых невероятных местах – на ногах, крыльях, антеннах.

Рис.9. Образование глаза в необычном
месте тела дрозофилы, продемонстриро-
ванное в опытах В.Геринга.
Из опытов Геринга можно сделать ряд заключений.
1. Действительно существуют «гены-господа» и «гены-рабы».
2. Сложнейшая морфогенетическая реакция, завершающаяся формированием целого органа, может быть запущена одним, «главным» геном, который, следовательно, является ответственным за процессы морфогенеза, разрешая или запрещая (в случае мутации) целый сложный комплекс формообразовательных событий.
3. Морфогенетические процессы основываются на молекулярно-генетических событиях, специфика формы обуславливается спецификой последовательных ткане- и органоспецифических синтезов, разрешенных активацией «главного» гена.
4. Соответствующие молекулярно-морфогенетические системы являются высококонсервативными и обеспечиваются в высшей степени гомологичными молекулярно-генетическими системами у самых разнообразных таксономических единиц.
Активность регуляторных генов предопределяет тканевую специфичность, реализуемую в процессе дифференциации.
Дифференциация– это процесс специализации клеток, обуславливающий их морфофизиологические различия. Другими словами, это реализация той программы, которая была намечена детерминацией. Различные виды клеток эукариотического организма синтезируют как одинаковые белки, так и специфические. В зависимости от типа клеток и стадии их развития может варьировать и уровень продукции любого белка. В связи с этим в биологии развития различают два вида эукариотических генов:
1) гены «домашнего хозяйства» (housekeeping) – связаны с поддержанием универсальных клеточных функций. Проявляются во всех клетках.
2) гены «роскоши» - связаны с осуществлением специализированных клеточных функций, специфичных для отдельных типов клеток. Высокий уровень активности генов «домашнего хозяйства» является обычно предварительным этапом дифференциации, следующим после установления детерминации. Этот этап, вероятно, не имеет тканевой специфичности. Известно, что клеточная детерминация происходит задолго до формообразовательных процессов.
Дифференциальная экспрессия генов «роскоши» обуславливает дифференциацию клеток в определенном направлении. В результате дифференциальной активности генов формируются различные клеточные линии, а на их основе – ткани и органы. Дифференциация как и детерминация, является многоступенчатым процессом. Так, на дифференцировку нервной ткани влияют экспрессия ее собственных генов, генов смежных нейронов, генов удаленных нейронов, отростки которых достигают этой клетки. Также на нее влияет дифференциальная экспрессия клеток окружающих глиальных клеток, генетические системы эндокринных и нейроэндокринных органов.
По мере развития зародыша усиливаются связи между клетками и увеличивается их влияние друг на друга. Влияние клеточных структур, определяющее развитие других клеточных структур, называется эмбриональной индукцией.
Другими словами под эмбриональной индукцией понимают взаимодействие эмбриональных закладок, ведущее к формообразовательному эффекту. Этот эффект достигается через реакцию ткани-мишени, которая становится детерминированной к определенному типу развития. Далее детерминированное состояние реализуется в процесс дифференцировки.
Первичной эмбриональной индукцией принято называть взаимодействие хордомезодермы и презумптивной нейроэктодермы в ходе гаструляции, обуславливающее процесс первичного органогенеза – формировании нервной трубки. В результате этого взаимодействия запускается цепь морфогенетических (формообразовательных) событий. В зависимости от типа образуемых при этом структур различают следующие варианты первичной эмбриональной индукции:
· архенцефалическая – в результате нее образуются предний мозг, глаз, хрусталик;
· дейтеренцефалическая – средний мозг, задний мозг, слуховые пузырьки;
· мезодермально-энтодермальная – представлена тремя вариантами:
1. спино-каудальная – спинной мозг, хвостовая хорда, сомиты;
2. туловищно-мезодермальная – туловищная хорда, сомиты, почечные канальцы, мезотелий, кровяные островки;
3. энтодермальная – глотка, пищевод, кишечник.
Явление первичной эмбриональной индукции открыл в 20-е годы ХХ в. выдающийся немецкий эмбриолог Г.Шпеман. он считал, что весь процесс индивидуального развития складывается из цепи эмбриональных индукций, шаг за шагом определяющих формообразование, дифференцировку органов и их систем и становление внешнего облика развивающейся особи, ее регионализацию.
Шпеман, проводя опыты по трансплантации участка губы бластопора от одного зародыша тритона на брюшную часть другого зародыша, получил удивительный результат: на брюшной стороне эксперементального зародыша формировалась вторая нервная пластинка, развившаяся позднее в нервную трубку. На каждой ее стороне образовывались слуховые пузырьки, ряды сомитов и таким образом возникала вторая закладка осевых органов. В 50-60-е годы ХХ в. голандский эмбриолог П.Нюкооп продемонстрировал, что первым индуцирующим событием в развитии зародыша является не воздействие хордомезодермы на презумптивную нервную пластинку, а стимуляция энтодермой преобразования смежных клеток в хордомезодермальную закладку. По сути дела, это событие и является истинно первичной эмбриональной индукцией.
Каждая эмбриональная индукция характеризуется тремя факторами: индуктор, компетентная ткань, взаимодействие индуктора и компетентной ткани. Индуктор – тканевая закладка, воздействующая на компетентную ткань. Характеризуется рядом особенностей: образованием индуцирующих агентов, созреванием способности к индукции, автономностью созревания. Индуцирующие агенты это низкомолекулярные белки, которые выделяются индуцирующей тканью и воспринимаются компетентной тканью, вызывая формообразовательный эффект. Предполагалось существование двух индуцирующих агентов – нейтролизующего и мезодермализующего, от соотношения которых зависит региональная специфичность индуцирующего эффекта. Они образуются еще в период оогенеза и распределяются по объему развивающего яйца неравномерно, формируя градиенты распределения, важные для нормально процесса эмбрионального развития.
Созревание способности к индукции заключается в регионализации – постепенном приобретении каждой частью индуцирующей ткани способности индуцировать определенный набор структур. На самых ранних стадиях созревания она обладает способностью индуцировать лишь образование пигментных клеток, затем переднеголовных структур и, наконец, мезодермальных структур. Индуцирующая хордомезодерма постепенно расчленяется на зоны, дающие разный региональный эффект. Созревание индуктора осуществляется автономно, независимо от окружающих тканей зародыш. В ходе созревания изменяются индуцирующие свойства ткани в строго определенном направлении и закономерно.
Компетентная ткань это та тканевая закладка, которая подвергается действию индуктора и отвечает на него формообразовательным процессом. Ее характеризуют три свойства: компетенция, автономность созревания, эффект минимальной масс.
Компетенция это физиологическое состояние реагирующей системы, в котором она способна воспринимать воздействие индуктора. До сих пор неизвестны молекулярно-генетические основы компетенции, но предполагают, что она может быть связана с образованием рецепторов, способных «опознавать» молекулы индуктора и обеспечивать их транспортировку внутрь клетки.
Созревание компетентной ткани, как и созревание индуктора, автономно, строго направлено, закономерно и не зависит от тканевого окружения. Чтобы компетентная ткань реагировала на действие индуктора, кроме всего прочего, необходимо наличие в ней определенного, минимального количества клеток, т.е. требуется некоторый «порог массы».
Взаимодействие индуктора и компетентной ткани характеризуют следующие три параметра: проникновение индуцирующих агентов в компетентную ткань, пространственные закономерности взаимодействия, временные закономерности взаимодействия. В опытах Ж.Браше, К.Гробстайна, Л.Саксена и др. было показано, что отделение индуктора от компетентной ткани с помощью мембранных фильтров, предотвращающих их контакт, не допускающих диффузию индуцирующих агентов, не предотвращает индукцию. Более того, если прокультивировать индуцирующую ткань в физиологическом растворе, затем удалить ее и поместить в этот раствор компетентную ткань, последняя обнаружит способность ко всем формообразовательным процессам, происходящим в ходе обычной индукции. Это наводит на мысль, что для осуществления индуцирующего эффекта важен не контакт индуктора и компетентной ткани, а транспорт индуцирующих агентов в компетентную ткань.
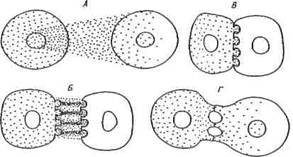
Рис.13. Возможные механизмы передачи индукционных влияний
А — диффузия; Б — взаимодействие молекул матрикса с комплементар-
ными структурами мембран клеток-мишеней; В— взаимодействие между поверхностными мембран-
ными структурами клеток ткани-индуктора и реагирующей ткани;
Г — взаимодействие через
межклеточные соединения
Пространственные закономерности взаимодействия индуктор-компетентная ткань проявляются в строгом соответствии положения ткани индуктора и реагирующей системы друг относительно друга.
Временные закономерности взаимодействия индуктор-компетентная ткань проявляются в величине времени контакта тканей организатора и реагирующей системы для достижения положительного эффекта индукции.
Для регионализации основное значение имеет не продолжительность контакта, а строго пространственно-временное соответствие физиологически активного состояния индуктора и реагирующей системы. Нарушение этого соответствия ведет к нарушению эффекта индукции.
К первому морфогенетическому взаимодействию в ходе развития амфибий относят открытое выдающимся голландским эмбриологом П.Ньюкоопом взаимодействие между анимальной и вегетативной частями зародыша на стадии морулы. В результате этого воздействия в экваториальной зоне зародыша возникает зачаток мезодермы, а остальные клетки анимальной половины детерминируются в эктодермальном направлении. Разные зоны вегетативного полюса индуцируют в эктодерме мезодермальные территории (дорсальную и вентральную) с различной судьбой.
Источником последующих индукционных влияний является зачаток мезодермы. В процессе развития он разделяется на две зоны – дорсальную и вентральную. В период гаструляции-нейруляции различные участки мезодермы приходят в контакт с другими тканями зародыша и оказывают на них индуцирующее влияние. В результате к концу нейруляции в пределах каждого из трех зародышевых листков (эктодерма, мезодерма, энтодерма) имеются определенные компартменты, содержащие клетки-предшественники соответствующих линий дифференцировки. Первым молекулярным сигналом в цепи индуктивных событий в ходе эмбриогенеза амфибий является активин.
Можно выделить несколько особенностей функционирования генетических систем, контролирующих развитие.
«Опережающее» функционирование генов в ходе онтогенеза. Многие продукты синтезируются в развивающемся зародыше «заранее», часто задолго до того, как они будут востребованы. Это, в частности, вещества, которые участвуют в «разметке» плана строения организма (продукты генов сегментации, гомеозисных генов), в осуществлении эмбриональной индукции (индуцирующие вещества и их ингибиторы). Некоторые молекулы, например, гемоглобин, образуются еще в яйцеклетке, задолго до того, как начнут выполнять свои функции в клетках эритроидного ряда.
Автономия частей при единстве целого. Это качество функциональной динамики генома отчетливо проявляется в раннем эмбриогенезе при созревании индуктора (хордомезодермы) и компетентной ткани (нейроэктодермы). Очевидно, генетические системы, регулирующие созревание индуцрующих свойств хордомезодермы и способность компетентной ткани реагировать на воздействие индуктора, функционируют в автономном режиме, независимо от того, находится ли данная развивающаяся эмбриональная закладка в составе целого зародыша или вне его. Целостность же развивающейся системы обеспечивается за счет того, что в норме сроки созревания двух взаимодействующих тканей строго «подогнаны» один к другому, как бы «синхронизированы» в результате чего достигается нормальное течение онтогенетического процесса.
Можно выделить три автономно функционирующих генетических системы, контролирующих три соответственно автономных процесса:
§ формообразовательные события;
§ дифференцировка специфических морфологических типов клеток;
§ химическая спецификация этих клеток.
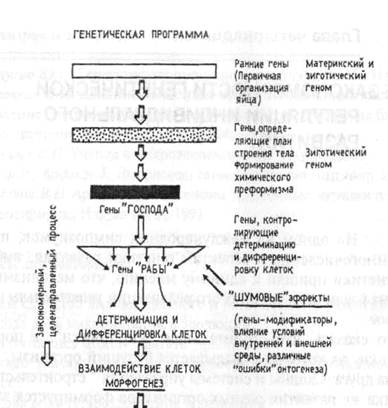
Схема генетического контроля индивидуального развития на разных этапах
– Конец работы –
Используемые теги: нцип, дифференциальной, активности, генов, основа, гетерогенизации, регионализации, развивающ, организма0.123
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Принцип дифференциальной активности генов как основа гетерогенизации (регионализации) развивающегося организма
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.201 сек.
Новости и инфо для студентов