дХТТСГМШИ ОНРЕМЖХЮК ΔφД.
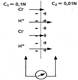
дКЪ ЕЦН БНГМХЙМНБЕґМХЪ МЕНАУНДХЛ ЙНМРЮЙР ЩКЕЙґРПНКХРНБ Я ПЮГКХВМНИ ЙНМЖЕМРПЮЖХЕИ Х ПЮГКХВґМНИ ОНДБХФМНЯРЭЧ ЮМХНМНБ Х ЙЮРХНМНБ (UH+ ╩ UCl- ). цПЮМХЖЮ ПЮГДЕКЮ НДХМЮЙНБН ОПНМХґЖЮЕЛЮ ДКЪ НАНХУ ХНМНБ. оЕПЕУНД ХНМНБ м+ Х я1~ АСДЕР НЯСЫЕґЯРБКЪРЭЯЪ Б ЯРНПНМС ЛЕМЭЬЕИ ЙНМЖЕМРПЮґЖХХ. б ЯХКС АНКЭґЬЕИ ОНДБХФМНЯРХ ХНМНБ БНДНґПНДЮ, АСДЕР ЯНГДЮБЮРЭЯЪ ХУ АНКЭЬЮЪ ЙНМЖЕМРПЮґЖХЪ Я ОПЮґБНИ ЯРНПНМШ НР ЦПЮМХЖШ ПЮГґДЕКЮ ЩКЕЙґРПНКХРНБ, БНГМХЙМЕР ПЮГМНЯРЭ ОНРЕМґЖХЮКНБ ΔφД. йНКХВЕЯРБЕММН БЕКХВХМЮ ΔφД НОПЕДЕКЪЕРЯЪ СПЮБМЕМХЕЛ цЕМДЕПЯНМЮ:
ΔφД = ((U+ - U- * RT)/(U+ = U- *nF))ln C1/C2
2. пЮБМНБЕЯМШИ ЛЕЛАПЮМґМШИ ОНРЕМЖХЮК ΔφЛ(П).
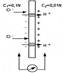 нМ БНГМХЙЮЕР МЮ ЦПЮМХЖЕ ПЮГґДЕКЮ ДБСУ ЩКЕЙРПНКХґРНБ, ПЮГДЕКЕММШУ ЛЕЛґАПЮМНИ, ХГАХПЮґРЕКЭМН ОПНМХЖЮЕЛНИ ДКЪ НДМНЦН ХГ ХНМНБ. щРН ВЮЯРМШИ ЯКСВЮИ ДХТТСГМНЦН ОНґРЕМЖХЮКЮ, ЙНЦДЮ ОНДБХФґМНЯРЭ НДМНЦН ХГ ХНМНБ ПЮБМЮ МСКЧ. оПХ ОНДБХФґМНЯРХ ХНМНБ яl- ПЮБМНИ МСКЧ, ХНМШ Cl- НЯРЮЧРЯЪ ОН НДМС ЯРНПНМС ЛЕЛґАПЮМШ, Ю ХНМШ м+ ОПНУНДЪР ВЕПЕГ ЛЕЛАПЮМС, НАґПЮГСЪ МЮ ДПСЦНИ ЯРНПНМЕ ОНКНФХРЕКЭМШИ ГЮПЪД. бНГМХЙЮЧЫХИ ОНРЕМЖХЮК НОХЯШБЮЕРЯЪ СПЮБМЕґМХЕЛ мЕПМЯРЮ, ВЮЯРМШИ ЯКСВЮИ СПЮБМЕМХЪ цЕМґДЕПЯНМЮ, ЙНЦДЮ UCl- = 0:
нМ БНГМХЙЮЕР МЮ ЦПЮМХЖЕ ПЮГґДЕКЮ ДБСУ ЩКЕЙРПНКХґРНБ, ПЮГДЕКЕММШУ ЛЕЛґАПЮМНИ, ХГАХПЮґРЕКЭМН ОПНМХЖЮЕЛНИ ДКЪ НДМНЦН ХГ ХНМНБ. щРН ВЮЯРМШИ ЯКСВЮИ ДХТТСГМНЦН ОНґРЕМЖХЮКЮ, ЙНЦДЮ ОНДБХФґМНЯРЭ НДМНЦН ХГ ХНМНБ ПЮБМЮ МСКЧ. оПХ ОНДБХФґМНЯРХ ХНМНБ яl- ПЮБМНИ МСКЧ, ХНМШ Cl- НЯРЮЧРЯЪ ОН НДМС ЯРНПНМС ЛЕЛґАПЮМШ, Ю ХНМШ м+ ОПНУНДЪР ВЕПЕГ ЛЕЛАПЮМС, НАґПЮГСЪ МЮ ДПСЦНИ ЯРНПНМЕ ОНКНФХРЕКЭМШИ ГЮПЪД. бНГМХЙЮЧЫХИ ОНРЕМЖХЮК НОХЯШБЮЕРЯЪ СПЮБМЕґМХЕЛ мЕПМЯРЮ, ВЮЯРМШИ ЯКСВЮИ СПЮБМЕМХЪ цЕМґДЕПЯНМЮ, ЙНЦДЮ UCl- = 0:
ΔφЛ(П) = (RT)/(nF) ln(C1/C2)
оН СПЮБМЕМХЧ мЕПМЯРЮ АШКХ ПЮЯЯВХРЮМШ ЛЕЛґАПЮММШЕ ОНРЕМґЖХЮКШ ЙКЕРЙХ, ЯНГДЮБЮЕЛШЕ ХНґМЮЛХ Na+, K+, C1ґґґґґ- Б НРДЕКЭМНЯРХ. нДґМЮЙН, ЯСЛґЛЮПМШИ ОНРЕМЖХЮК ЩРХУ ХНМНБ ГМЮВХРЕКЭМН НРКХґВЮЕРЯЪ НР ЩЙЯОЕПХЛЕМРЮКЭМН ХГґЛЕПЕММНЦН МЮ ПЮГґКХВМШУ ЙКЕРЙЮУ. бНГМХЙКЮ МЕНАУНґДХЛНЯРЭ ЯНГДЮґМХЪ АНКЕЕ СМХБЕПЯЮКЭМНИ РЕНПХХ, НАЗЪЯґМЪЧЫЕИ БНГМХЙМНґБЕМХЕ АХНОНРЕМЖХЮКНБ ОНЙНЪ ЙКЕРЙХ.
3. яРЮЖХНМЮПМШИ ЛЕЛАПЮММШИ ОНРЕМЖХЮК ΔφЛ(c). уНДФЙХМ Х йЮРЖ ОПЕДОНКНФХґКХ, ВРН ОНґРЕМЖХЮК ОНЙНЪ
ЙКЕРЙХ ЪБКЪЕРЯЪ МЕ ПЮБМНБЕЯМШЛ, Ю ЯРЮЖХНМЮПґМШЛ, Р.Е. НМ НАСЯґКНБКЕМ ОНДБХФМШЛ ПЮБМНБЕЯХЕЛ ОНРНЙНБ ХНМНБ: Na+, K+, я1-.