Стандартная свободная энергия гидролиза органических фосфатов
| Соединение | Стандартная энергия гидролиза (∆G0) | |
| кДж/ моль | ккал/моль | |
Фосфоенолпируват
Карбамоилфосфат
1,3-Дифосфоглицерат
Креатинфосфат
 АТФ АДФ + Рi АТФ АДФ + Рi
 АДФ АМФ + Рi
Пирофосфат
Глюкозо-1-фосфат
Фруктозо-6-фосфат
АМФ
Глюкозо-6-фосфат
Глицерол-3-фосфат АДФ АМФ + Рi
Пирофосфат
Глюкозо-1-фосфат
Фруктозо-6-фосфат
АМФ
Глюкозо-6-фосфат
Глицерол-3-фосфат
| -61, 9 -51,4 -49,3 -43,1 -30,45 -27,6 -27,6 -20,9 -15,9 -14,2 -13,8 -9,2 | -14,8 -12,3 -11,8 -10,3 -7,3 -6,6 -6,6 -5,0 -3,8 -3,4 -3,3 -2,2 |
Самой многочисленной группой макроэргов являются фосфаты, хотя имеются представители и других классов:
Органические фосфаты:
а) нуклеозидполифосфаты: АТФ, УТФ, ГТФ и др.
б) ацилфосфаты: в) амидинфосфаты: г) енолфосфаты:
|
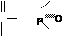
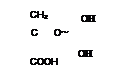
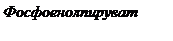
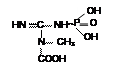
|
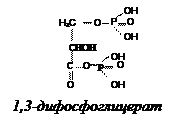
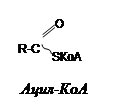
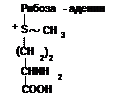
S-аденозилметионин
|
клетках данный макроэрг выступает в качестве депо для кратного хранения и переноса химической энергии на молекулзультате гидролиза АТФ или ее образование в ходе фосфорилирования (синтез АТФ). Однако такой подход односторонен. Для полного понимания необходимо рассмотреть энергетику двух процессов. Следует иметь в виду, что АТФ гораздо менее стабильная молекула, чем аденозиндифосфат (АДФ). Другими словами, АТФ более высокоэнергетическое соединение, чем АДФ.

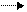 Превращение АТФ АДФ + Фн сопровождается освобождением энергии, а обратная реакция АДФ + Фн АТФ – требует её затраты. Аналогичные соображения применимы для описания гидролиза АТФ до АМФ и пирофосфата (ФФн).
Превращение АТФ АДФ + Фн сопровождается освобождением энергии, а обратная реакция АДФ + Фн АТФ – требует её затраты. Аналогичные соображения применимы для описания гидролиза АТФ до АМФ и пирофосфата (ФФн).
 Обратимый процесс АТФ АДФ + Фн (или АМФ + ФФн) составляет молекулярную основу переноса химической энергии внутри всех живых клеток.
Обратимый процесс АТФ АДФ + Фн (или АМФ + ФФн) составляет молекулярную основу переноса химической энергии внутри всех живых клеток.
Когда разрушается высокоэнергетический компонент пищи, например, углевод, освобождающаяся энергия используется для образования АТФ. Если клетка нуждается в энергии (для таких процессов, как биосинтез, сокращение мышцы или транспорт соединений через мембраны), АТФ гидролизуется с её выделением (рис. 3).

Рис. 3. Пути использования энергии макроэргов