Новый взгляд на происхождение билатерий
Новый взгляд на происхождение билатерий
Владимир Васильевич Малахов, член-корр. РАН, зав. каф. зоологии беспозвоночных МГУ им.М.В.Ломоносова. Одна из загадок развития жизни на Земле - “кембрийский взрыв”. Так… Кишечнополостные по сравнению с билатериями, действительно, устроены гораздо проще, у них нет ни сквозного кишечника…Сравнительная анатомия
За последние 150 лет специалисты по сравнительной анатомии разработали несколько десятков гипотез о происхождении билатерально-симметричных животных. Все эти гипотезы можно разделить на три группы: планулоидно-турбеллярные, архицеломатные и метамерные.
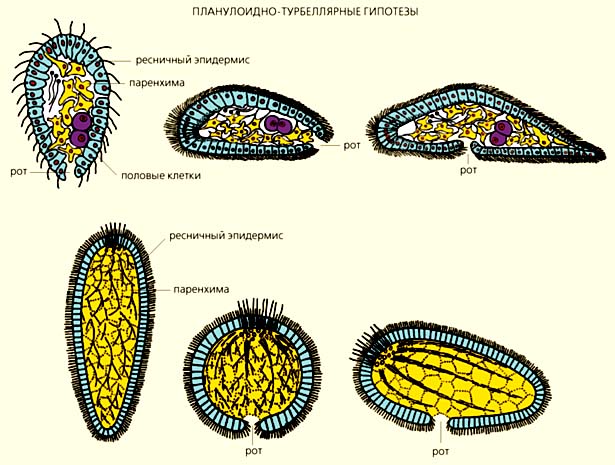
Схемы, отражающие различные взгляды сравнительных анатомов на происхождение двусторонне-симметричных животных. По мнению сторонников планулоидно-турбеллярных гипотез (вверху), предками билатерий были организмы, напоминающие личинок современных кишечнополостных животных (планул). По одной из версий (верхний ряд), брюшная сторона первичных билатерий образовалась из бокового сектора планулоидного предка [4], по другой, - из ротовой поверхности [5].
Согласно архицеломатным гипотезам (внизу), билатерии произошли от четырехлучевых коралловых полипов, кишечная полость которых разделена на четыре камеры [7], а метамерным (ниже всех) - от многолучевых кораллов (вид сверху и сбоку [10], а также со стороны ротовой поверхности [9]).

Согласно планулоидно-турбеллярным гипотезам, все или, по крайней мере, некоторые билатерально-симметричные животные произошли от организмов, напоминавших личинок современных кишечнополостных - планул. На заднем конце таких предковых форм находился рот, а на противоположном (аборальном органе) - скопление ресничных чувствительных клеток. Сначала планулоидные организмы плавали в толще воды, затем опустились на дно и перешли к ползанию по субстрату, что и способствовало формированию билатеральной симметрии. По мнению сторонников этих гипотез, первичные билатерии были очень простыми организмами, они не имели сквозного кишечника и целома. Их потомками стали ресничные черви (Turbellaria), от которых происходят другие двусторонне-симметричные животные.
В понимании, как именно возникла брюшная сторона у билатерий, разные авторы были не столь единодушны. Например, отечественные зоологи (В.Н.Беклемишев, А.В.Иванов и Ю.В.Мамкаев и др.) убеждены, что у ресничных червей она гомологична боковому сектору (антимере) планулоидного предка [3, 4]. Американская исследовательница Л.Гаймен считала, что предки билатерально-симметричных животных стали ползать на ротовой поверхности, которая и преобразовалась в брюшную сторону [5].
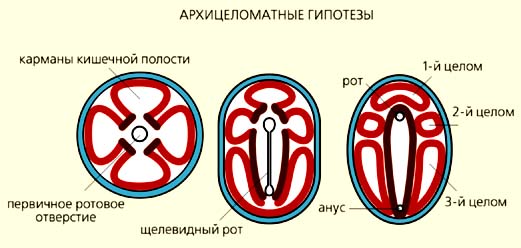
Приверженцы архицеломатных гипотез предполагали, что предками билатерий были четырехлучевые полипы, кишечная полость которых разделена перегородками на четыре камеры [6-8]. Такие полипы опустились на грунт и начали перемещаться на ротовой поверхности, которая впоследствии превратилась в брюшную сторону. Первичный рот вытянулся и стал щелевидным (как у современных коралловых полипов), а затем сомкнулся посредине так, что от него остались только два отверстия: одно - на переднем конце тела и стало ртом, а другое - на заднем и преобразовалось в анус. Кишечные карманы отделились от центральной части кишечника и дали начало пяти камерам вторичной полости тела - целому, расчлененному на три отдела. По мнению сторонников архицеломатных гипотез, все нецеломические билатерально-симметричные организмы (например, плоские и круглые черви) утратили вторичную полость тела.

Ранние стадии эмбрионального развития некоторых групп билатерально-симметричных животных. Красным цветом выделен бластопор и его превращения у иглокожих, ресничных и кольчатых червей в брюшную сторону, а у хордовых - в спинную. Пояснения в тексте.
Сторонники метамерных гипотез также выводили билатерально-симметричных животных от кораллов, но не от четырехлучевых полипов, а многолучевых, у которых кишечная полость поделена на множество камер многочисленными перегородками-септами, например, как у шестилучевого коралла цериантарии [9-11]. Как и в архицеломатных гипотезах, предполагалось, что такие кораллы стали ползать на ротовой стороне, щелевидный рот замкнулся посредине, а камеры кишечной полости отделились от центральной части кишечника и дали начало многочисленным целомическим камерам. Только в этом случае круговое расположение камер кишечной полости (цикломерия) у радиально-симметричных форм превратилось в метамерию первичных билатерий, а расположенные по кругу щупальца - в боковые метамерные придатки (параподии), служащие органами движения. Таким образом, первичные билатерии были не только целомическими, но и сегментированными организмами с метамерными придатками - зачатками конечностей.
Классическая эмбриология
У примитивных билатерально-симметричных животных (морских беспозвоночных, которые сохранили наружное оплодотворение, и плавающих в толще воды ресничных личинок), как уже говорилось, ранние стадии эмбрионального развития - дробления, бластулы и гаструлы - подчиняются радиальной симметрии. Происходит это путем разрастания одного из секторов гаструлы - именно того, который впоследствии станет спинной стороной личинки. В результате меняется взаимное расположение первичного нервного центра (аборального органа) и отверстия первичной кишечной полости (бластопора). Аборальный орган смещается вперед, а бластопоральная сторона становится брюшной стороной будущей личинки. Такой процесс характерен для подавляющего большинства групп билатерально-симметричных животных. Единственное исключение - хордовые, у которых бластопоральная сторона превращается в спинную часть организма. Тому есть свое объяснение: хордовые - это перевернутые билатерии [12].
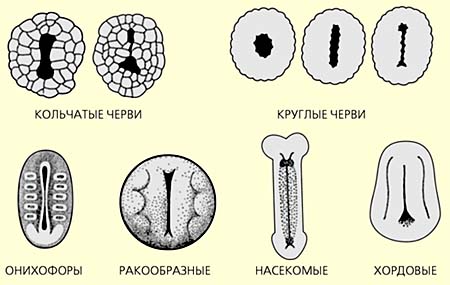
Разновидности щелевидного бластопора у эмбрионов билатерально-симметричных животных.
Таким образом, данные эмбриологии помогли выяснить, во-первых, что брюшная сторона билатерально- симметричных животных представляет собой…Палеонтология
Трудно сказать, почему вендские многоклеточные животные были бесскелетными. Возможно, они обитали в холодноводных бассейнах вне контролируемого… Формы с радиальной или осевой симметрией очень многочисленны в венде и во…Прорисовки отпечатков вендских билатерий.
Вендские билатерально-симметричные организмы. Реконструкции М.А.Федонкина.
Как у современных кишечнополостных, их аборальная (т.е. противоположная рту) сторона была одета уплотненным защитным покровом - кутикулой. Правда, у вендских билатерий этот покров был органическим, а у современных кишечнополостных может быть как органическим, так и известковым. В отличие от современных кишечнополостных, которые прикреплены к субстрату, вендские двусторонне-симметричные кишечнополостные были подвижными организмами, ползавшими на ротовой поверхности.
Молекулярная биология
Схема распределения зон экспрессии генов, содержащих гомеобоксы, у кишечнополостных (слева) и эмбрионов билатерий.
Эта система выявлена у всех многоклеточных животных, в том числе и у современных кишечнополостных [17-19]. У них найдены, в частности, гомологи генов Brachiury, goosecoid и fork head. У эмбрионов билатерально-симметричных животных эти гены экспрессируются в процессе гаструляции вдоль щелевидного бластопора на переднем и заднем краях или вокруг продуктов его разделения - рта и ануса. Они играют важную роль в формировании переднего и заднего мезодермальных зачатков у позвоночных и беспозвоночных животных. У кишечнополостных гомологи этих генов экспрессируются в кольцевой области вокруг ротового отверстия [20-22]. Очевидно, эмбриональные организаторы головных и туловищно-хвостовых структур, которые у билатерий связаны с передним и задним концами щелевидного бластопора, произошли из единого кольцевого организатора кишечнополостных. Интересно, что некоторые гены, например fork head, задействованы в переднем и заднем организаторах (иногда еще и в серии участков вдоль линии замыкания бластопора), тогда как другие, например goosecoid и Brachiury, сегрегированы: goosecoid в переднем, Brachiury - в заднем. Такое расщепление первоначально единого организатора могло возникнуть только в результате удлинения первичного ротового отверстия, смыкания его посредине и превращения отверстий на его концах в дефинитивные рот и анус билатерий, как это предполагалось в построениях классической сравнительной анатомии.
Наиболее вероятный путь происхождения билатерий
В вендском периоде существовала обширная фауна радиально-симметричных кишечнополостных животных, часть из которых перешла к ползанию по субстрату на…Происхождение билатерально-симметричных животных. Красным цветом выделен аборальный нервный центр.
Не исключено, что вендские двусторонне-симметричные кишечнополостные были предками не только трехслойных билатерий, но и фанерозойских…Некоторые представители кембрийских билатерально-симметричных животных.
Большая морфологическая сложность вендских предков билатерий объясняет так называемый кембрийский взрыв, т.е. почти одновременное появление в…Литература
1. Федонкин М.А. // Итоги науки и техники. Сер. Стратиграфия и палеонтология. 1983. Т.12. С.3-127.
2. Соколов Б.С. Очерки становления венда. М., 1997.
3. Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. М., 1952.
4. Иванов А.В., Мамкаев Ю.В. Ресничные черви (Turbellaria), их происхождение и эволюция. Филогенетические очерки. Л., 1973.
5. Hyman L.H. Plathyhelminthes and Rhynchocoela. The acoelomate Bilateria // The Invertebrates. N.Y.; Toronto; London, 1951. V.2.
6. Masterman A.T. // Quart. J. Microsc. Sci. 1897. V.40. P.59-71.
7. Remane A. // Verh. Deutsch. Zool. Ges. 1950 (1949). S.16-23.
8. Siewing R. // Zool. Jb. Abt. Anat. 1980. B.103. S.439-482.
9. Sedgwick A. // Quart. J. Microsc. Sci. 1884. V.24. P.43-82.
10. Beneden E. van. // Arch. Biol. Paris. 1891. T.11. P.114-146.
11. Lameere A. Precis de zoologie. Liege. Desoer. 1932. T.2.
12. Малахов В.В. // Журн. общ. биологии. 1977. Т.38. ?4. С.485-499.
13. Федонкин М.А. Беломорская биота венда (Докембрийская бесскелетная фауна севера Русской платформы) // Тр. ГИН АН СССР. 1981. Вып.342. С.3-100.
14. Федонкин М.А. Бесскелетная фауна венда и ее место в эволюции Metazoa. М., 1987.
15. Федонкин М.А. // Природа. 2000. №9. С.3-11.
16. Иванцов А.Ю., Федонкин М.А. Следы самостоятельного передвижения - финальное доказательство животной природы эдиакарских организмов // Эволюция жизни на Земле: Материалы II междунар. симпозиума. Томск, 2001. С.133-137.
17. Finnerty J.R., Martindale M.Q. // Biol. Bull. 1997. V.193. P.62-76.
18. Broun M., Bode H.R. // Debelopment. 2002. V.129. P.875-884.
19. Scholz C.B., Technau U. // Dev. Genes Evol. 2003. V.212. P.563-570.
20. Wiegel D., Jurgens G., Kuttner F. et al. // Cell. 1989. V.57. P.645-658.
21. Technau U. // Bioessays. 2001. V.23. P.788-794.
22. Lartillot N., Le Gour M., Adoutte A. // Dev. Gen. Evol. 2002. V.212. P.551-561.