рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Биология
- /
- N I II C
Реферат Курсовая Конспект
N I II C
N I II C - раздел Биология, Эукариотические ДНК-полимеразы и ДНК-полимеразы археев Рис. 2.8. Схема Организации Белка Dnac E. Coli. I – Область В...
Рис. 2.8. Схема организации белка DnaC E. coli.
I – область взаимодействия с белком DnaB,
II – мотивы связывания АТФ
Количество белка DnaС на клетку E. coli равно ~20, т.е. совпадает с числом молекул DnaB. Гексамер геликазы DnaB стехиометрически связывает 6 мономеров DnaС в форме со связанным АТФ. Молекулы DnaС располагаются на поверхности одного из оснований кольца DnaB и, вероятно, ассоциируются с С-концевой половиной DnaB. Такое взаимодействие, стабилизируемое АТФ, “замораживает” гексамер DnaB в треугольной конформации с симметрией С3 и закрывает центральный канал DnaB на стороне, противоположной сайтам связывания DnaС. В результате через этот канал не может пройти даже онДНК.
Образование комплекса DnaB-DnaС изменяет свойства обоих партнеров. Белок DnaB утрачивает все свои каталитические активности, включая НТФазную и геликазную, а у белка DnaС активируется его “скрытая” (cryptic) способность связываться с онДНК. В результате комплекс DnaB-DnaС может ассоциироваться с голой онДНК, но не с ДНК, покрытой белком SSB. Этот механизм предотвращает неразборчивую погрузку геликазы DnaB на участки хромосомы, временно ставшие однонитевыми (например, в результате эксцизионной репарации ДНК) и покрытые SSB. Во время инициации репликации хромосомы E. coli в области oriC участки голой онДНК создаются белком-инициатором DnaА (гл.3), за счет взаимодействия с которым белка DnaB на них вербуется комплекс DnaB6-(DnaС-АТФ). После связывания с этими участками белок DnaС каким-то образом размыкает кольцо DnaB и пропускает внутрь него нить онДНК. Вероятно, это происходит так же, как при погрузке зажима ДНК-полимераз g-комплексом погрузчика (см. 1.00). Контакт белка DnaС с онДНК и DnaB запускает гидролиз АТФ, связанного с DnaС, после чего субъединицы DnaС-АДФ покидают комплекс с гексамером DnaB, и он приобретает ДНК-геликазную активность.
Cопряжение гидролиза НТФ с транслокацией по онДНК
Гидролиз НТФ ДНК-геликазами используется ими как источник энергии для перемещения по онДНК и для расплетания днДНК. Анализ равновесного связывания нуклеотидов показал, что у многих гексамерных геликаз из 6 субъединиц только три обладают высоким сродством к нуклеотидам, а остальные три имеют низкое сродство и не участвуют в связывании НТФ и в катализе гидролиза. С другой стороны, изучение предстационарной кинетики гидролиза НТФ позволило предположить, что в любой момент времени только одна связанная с гексамером молекула НТФ гидролизуется с высокой скоростью, т.е. три потенциальных каталитических центра в гексамерной геликазе участвуют в гидролизе НТФ не одновременно, а последовательно друг за другом. В этом отношении гексамерные геликазы напоминают другой, хорошо изученный ранее гексамерный фермент – F1-АТФазу мембранных протонных насосов. Последняя состоит из 3 неактивных структурных a-субъединиц и 3 каталитически активных b-субъединиц, которые работают не одновременно. В последовательном механизме действия этой АТФазы реакцию гидролиза АТФ можно разбить на 3 парциальные стадии: связывания АТФ, гидролиза АТФ и освобождения продуктов (АДФ и неорганического фосфата). В любой данный момент времени каждая из этих стадий осуществляется только какой-то одной из a-субъединиц: одна связывает АТФ, вторая гидролизует его, а третья освобождает продукты. В дальнейшем эти субъединицы, согласованно претерпевая последовательные изменения конформации, меняются друг с другом ролями.
Рассмотрим последовательную 3-сайтовую модель действия гексамерных ДНК-геликаз, основанную на аналогии с F1-АТФазой и модифицированную с учетом взаимодействия геликаз с ДНК. Предполагается, что в любой момент времени 3 активные субъединицы геликазы находятся в 3 разных конформационных состояниях. В состоянии Т субъединица связывает НТФ и одновременно имеет высокое сродство к онДНК. В состоянии D она связывает НДФ (продукт гидролиза НТФ) и проявляет более низкое сродство к ДНК, а в «пустом» состоянии Е субъединица свободна от нуклеотидов и онДНК. В первый момент в Т-состоянии находится субъединица 1, в D-состоянии субъединица 2 и в Е-состоянии субъединица 3 (рис. 2.9). Гидролиз НТФ субъединицей 1 вызывает её переход в D-состояние и вызывает одновременное изменение конформации двух остальных субединиц: субъединица 2 освобождает продукты гидролиза и становится «пустой» (переход в Е-состояние), а субъединица 3 связывает НТФ и оказывается в Т-состоянии. Реакция на каждой из субъединиц зависит от реакций, проходящих на 2 остальных субъединицах. Это обеспечивает последовательное протекание 3 стадий катализа (связывания НТФ, гидролиза НТФ и освобождения продуктов) на 3 активных сайтах геликазы. Такие циклы повторяются периодически, и после 3 циклов каждая субъединица геликазы возвращается в исходное состояние.
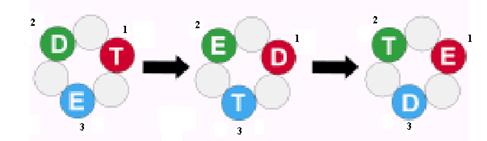
Рис. 2.9. Схема последовательных изменений состояния индивидуальных субъединиц (1, 2 и 3) в гексамерной ДНК-геликазе.
Т – сайт со связанным НТФ, D – сайт со связанным НДФ, Е – «пустой» сайт
Т.к. изменения конформационного состояния субъединиц приводят к изменению их сродства к онДНК, каждая из субъединиц должна последовательно прочно связываться с ДНК в состоянии Т, ослаблять свою ассоциацию с ДНК после гидролиза НТФ и перехода в состояние D , освобожаться от контакта ДНК при передоде в состояние Е и вновь связываться с ДНК, но уже в новом месте, после повторного связывания НТФ и возврата с состояние Т (рис. 2.10, А).
Такая последовательность событий в каждом из 3 сайтов геликазы может обеспечить перемещение ДНК-гелиеказы вдоль ДНК (рис. 2.10, В). В начальный момент времени субъединица 1, находящаяся в состоянии Т и прочно связанная с онДНК, претерпевает изменение конформации, инициирующее движение геликазы. Соседняя субъединица 2, слабо связанная с ДНК в состоянии D, освобождается из контатка с ДНК, а «пустая» субъединица 3 связывает НТФ и прочно связывается с ДНК, но уже в другомсайте. Хотя ДНК освобождается от геликазы в одном месте, в любой момент она остается связанной с двумя субъединицами геликазы. Повторение таких циклов изменения контактов субъединиц ДНК-геликазы с участками ДНК должно привести к однонаправленному процессивному перемещению ДНК-геликазы вдоль онДНК (механизм «активного вращения» - active rolling)

Рис. 2.10. Гипотетическая 3-сайтовая модель транслокации гексамерной ДНК-геликазы по онДНК, сопряженной с НТФазной активностью.
А. Последовательные изменения конформации индивидуальной субъединицы геликазы (I – гидролиз НТФ и ослабление связывания с ДНК, II – диссоциация НДФ и отрыв от ДНК, III – связывание НТФ и прочная ассоциация с новым сайтом в ДНК.
В. Последовательные стадии транслокации геликазы (1, 2 и 3 – номера индивидуальных субъединиц).
Обозначения различных состояний субъединиц – как на рис. 2.9
Расплетание днДНК
Этот аспект работы ДНК-геликаз наиболее труден для изучения, и все предложенные механизмы расплетания днДНК остаются гипотетическими. Их можно классифицировать как активные и пассивные, в зависимости от того, участвует ли геликаза в самом акте расплетания или просто стабилизирует участки онДНК. В пассивном механизме ДНК-геликаза косвенно облегчает расплетание, связываясь с онДНК, которая становится доступной в результате временного плавления двойной спирали, вызванного тепловыми флуктуациями на стыке между онДНК и ДНК. В этой модели ДНК-геликаза высупает как разновидность связывающих онДНК и дестабилизирующих дуплекс белков. В пассивном механизме ДНК-геликаза должна связываться с онДНК и однонаправленно перемещаться вдоль неё в направлении днДНК. Транслоцирующаяся ДНК-геликаза улавливает сегменты онДНК длиной в один или несколько нуклеотидов, спонтанно появляющиеся на стыке онДНК-днДНК. Пассивная модель не нашла экспериментального подтверждения. Ей противоречит и способность некоторых ДНК-геликаз связываться не только с онДНК, но и с днДНК.
Активные механизмы расплетания днДНК можно подразделить на три класса. Первые две модели не требуют прочного связывания ДНК-геликазы с днДНК. Модель клина (рис. 2.10, А) предполагает, что одна из расплетенных нитей дуплекса ДНК прочно связана в центральной отверстии кольца гексамерной ДНК-геликазы, а вторая расположена вне кольца и не взаимодействует с белком. При однонаправленном движении геликазы по нити ДНК, проходящей через центральный канал, энергия гидролиза НТФ порождают движущую силу, достаточную не только для перемещения по онДНК, но и для дестабилизации нескольких пар нуклеотидов в днДНК, примыкающей к онДНК. Движущаяся ДНК-геликаза, подобно клину, механически раздвигает эти спаренные основания. Во второй, торсионной модели (рис. 2.10, В) обе разделенные нити ДНК прочно связываются с ДНК-геликазой: одна в центральном канале, а вторая на внешней поверхности кольца. Эти сильные взаимодействия вызывают при транслокации ДНК-геликазы вращение двух нитей ДНК друг относительно друга и генерируют крутящий момент, который раскручивает две нити дуплекса на участке, примыкающем к уже расплетенным нитям. Третья модель активного действия ДНК-геликаз (модель дестабилизации дуплекса) предполагает, что геликаза взаимодействует в центральном канале или на поверхности гексамера не только с онДНК, но и со смежным сегментом дуплекса. Изменения конформации белка, обусловленные гидролизом НТФ, по неустановленному механизму дестабилизируют спираль днДНК в активном центре ДНК-геликазы и вызывают в этой области контакта с днДНК плавление нескольких п.н. После частичного расплетания дуплекса транслоцирующаяся ДНК-геликаза улавливает разошедшиеся нити ДНК. Эта модель похожа на пассивный механизм, но предполагает, что первичное разделение нитей днДНК вызвано не тепловыми флуктуациями, а изменениями конформации ДНК-геликазы.

– Конец работы –
Эта тема принадлежит разделу:
Эукариотические ДНК-полимеразы и ДНК-полимеразы археев
На сайте allrefs.net читайте: Эукариотические ДНК-полимеразы и ДНК-полимеразы археев...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: N I II C
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.064 сек.
Новости и инфо для студентов