рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Биология
- /
- Палеонтологические свидетельства эволюции.
Реферат Курсовая Конспект
Палеонтологические свидетельства эволюции.
Палеонтологические свидетельства эволюции. - раздел Биология, Возникновение и развитие эволюционной биологии. Данные Палеонтологии, Биогеографии, Сравнительной Анатомии, Эмбриологии И Мол...
Данные палеонтологии, биогеографии, сравнительной анатомии, эмбриологии и молекулярной биологии содержат убедительные и, что очень важно, взаимодополняющие и непротиворечивые свидетельства эволюции.Последовательное появление новых видов животных и растений в палеонтологической летописи Земли привело натуралистов XVIII-XIX к идее эволюции. В нижних слоях геологических отложений обнаруживались более примитивные организмы, в верхних, - все более и более сходные с современными. Дарвин писал «геология дает нам ясное доказательство, что все виды претерпели изменения, и притом эти изменения протекали медленно и постепенно. Мы это ясно усматриваем из того факта, что ископаемые остатки последовательных формаций неизменно гораздо более сходны друг с другом, чем ископаемые из формаций, далеко одна от другой отстоящих».
Археоптерикс и микрораптор гуи– переходные звенья в цепи эволюции от рептилий к птицам. В их строении сочетались признаки характерные для рептилий (наличие зубов, пальцев на передних конечностях, длинный хвостовой отдел позвоночника) и птиц (оперение).
Во времена Дарвина палеонтологическая летопись начиналась с Кембрийского периода (530 млн. лет назад) и в ней наблюдалось много пробелов. Сейчас эти пробелы постепенно заполняются. Палеонтологи нашли и охарактеризовали множество окаменевших останков животных, растений и микроорганизмов, живших на Земле в отдаленные эпохи. Современные методы физики и химии позволили очень точно датировать окаменелости по соотношению изотопов различных химических элементов в самих останках и окружающих их породах. Данные палеонтологии свидетельствуют, что микроорганизмы обитали на Земле 3,8 млрд. лет назад. Самые древние эукариотические клетки были обнаружены в породах, возраст которых превышает 2 млрд. лет. Чем ближе к нашему времени, тем меньше пробелов остается в палеонтологической летописи, тем более подробной она становится. Мы видим последовательное и постепенное появление новых форм. Более того, мы видим, как из прежних форм возникали новые. Между рыбами и амфибиями, амфибиями и рептилиями, рептилиями и млекопитающими найдено так много переходных форм, что зачастую трудно сказать, к какому классу позвоночных их следует относить. Рассмотрим некоторые примеры. Впервые ископаемые останки первоптицы - археоптерикса был найдены в Германии в 1861 году. Он имел перья и крылья, следовательно, относился к птицам. Но его скелет отличался рядом признаков, типичных для хищных динозавров: костистый хвост, зубы, пальцы с когтями на передних конечностях. Позднее в Испании и Китае были найдены на 30-40 млн. лет более молодые останки археоптериксов. Они еще больше похожи на птиц и меньше на динозавров. Их костистые хвосты стали короче, пальцы на крыльях редуцировались. Археоптерикс не исторический курьез, а одно из звеньев в цепи эволюции. Совсем недавно был найден далекий предок археоптерикса– микрораптор гуи. Он имел маховые перья не только на передних, но и на задних конечностях. Таким образом, у него было четыре крыла, которые он использовал для парящего полета. До него в палеонтологической летописи находятся динозавры, которые становились с течением времени все более и более похожими на птиц, а после него – древние птицы, которые все более и более утрачивали признаки динозавров. В этой длинной цепи от динозавров к птицам, которая протянулась более чем на 140 млн. лет, не было разрывов. Не было момента, когда птица вдруг вылетела из яйца динозавра. Каждый предок имел потомка, в основном похожего на себя, и в то же время отличного в деталях.
Следует помнить, однако, что многие ископаемые останки, принадлежат боковым ветвям филогенетического древа. На Земле нет сейчас живых потомков тиранозавров. Эта ветвь оказалась эволюционным тупиком. Нет на Земле и четырехкрылых птиц, хотя такие родственники археоптериксов существовали на Земле десятки миллионов лет назад.
Открытия 1990-х годов позволили восстановить интереснейшую историю постепенного превращения мелких (размером с белку) наземных копытных млекопитающих в гигантов моря – китов. Ископаемые останки свидетельствуют, что наиболее дальний предок современных китов жил на суше 65 млн. лет назад и кормился рыбой и трупами животных. Пакицетус, который жил около 50 млн. лет назад, уже перешел к полуводному образу жизни. Более молодой представитель этой линии амбулоцетус – ходячий кит - (45 млн. лет назад) был ростом с морского льва, имел довольно длинные конечности, хорошо приспособленные как для плавания, так и для хождения по дну, и длинный хвост, тоже полезный для плавания, хотя пока еще очень мало похожий на мощный хвостовой плавник кита. Родоцетус (43 млн. лет назад) сделал следующий и уже необратимый шаг в море. Его конечности сильно уменьшились, а тазовый пояс практически отделился от позвоночника. И, наконец, 40 млн. лет назад появился базилозаврус. Длина его тела достигала 15 метров. Конечности его уже полностью преобразовались в мощные плавники. Это был уже почти настоящий кит. Он еще сохранил явное сходство со своими сухопутными предками. Его ноздри уже сместились далеко назад, но им предстояло пройти еще полпути до того места на черепе, где сейчас располагается дыхальце настоящих китов. Он еще сохранил тазовый пояс, но на его рудиментарных ножках уже было не пять, а всего три крохотных пальчика. Таким образом, палеонтологическая летопись дает нам свидетельства постепенного преобразования наземных млекопитающих в обитателей моря. Несмотря на ее явную неполноту и фрагментарность, мы видим в ней серию переходных форм. По существу, все ископаемые останки представляют собой переходные формы – все они является потомками ранее живших форм, а некоторые – предками ныне живущих форм. Таким образом, палеонтологическая летопись является летописью эволюции - наследственного изменения свойств живых организмов в ряду поколений.
Молекулярные свидетельства эволюции.
Благодаря современным достижениям молекулярной биологии и генетики удалось расшифровать геном человека, мыши, дрозофилы, малярийного комара, риса и многих других видов животных, растений, грибов и микроорганизмов. Мы можем сравнивать гены разных организмов и анализировать сходство и различия между ними. Мы можем изучать гены не только ныне живущих организмов, но и давно вымерших видов, используя следы ДНК в ископаемых останках. Мы знаем, что все живые организмы имеют одинаковые механизмы записи, передачи и считывания наследственной информации. Эта информация записана в последовательности нуклеотидов ДНК (РНК у некоторых вирусов), она передается из поколения в поколение благодаря одному и тому же механизму репликации, она транскрибируется в последовательность нуклеотидов и-РНК и затем транслируется в последовательность аминокислот в белках с использованием одного и того же генетического кода. Это однозначно свидетельствует о том, что все ныне живущие на Земле организмы происходят от одного общего предка. Геном каждого вида представляет собой генетическую летопись его эволюции. Каждый организм получил свои гены от предков, а те в свою очередь от своих предков, и так до самого первого предка всех живых организмов. В большинстве случае гены передавались без изменений, но изредка возникали случайные изменения – мутации. Некоторые мутации снижали приспособленность организмов, которые их наследовали. Такие мутации отметались естественным отбором и не передавались дальше. Большинство никак не сказывались на приспособленности организмов, и, следовательно, на вероятности передачи этих измененных генов из поколения в поколение. Наконец, были такие мутации, которые давали их носителям преимущества в борьбе за жизнь. Они распространялись в популяциях и закреплялись в генофонде вида, внутри которого они возникли, и передавались дочерним видам. Таким образом, в геномах ныне живущих видов записана все история этих видов. Сравнение генов разных видов дает ключ к построению единой родословной всего живого на Земле.Многие мутации представляют собой замены одних нуклеотидов на другие. Мутации возникают, как правило, во время репликации ДНК. Отсюда следует, что, чем больше поколений прошло со времени дивергенции двух видов от общего предка, тем больше случайных замен нуклеотидов должно было накопиться в геномах дочерних видов. Соответственно, тем больше аминокислот отличают один и тот же белок у этих видов. Общий предок человека и мартышки существовал более 30 млн. лет назад, а общий предок человека и кошки — более 80 млн. лет назад. Когда мы сравниваем аминокислотные последовательности гемоглобина, мы видим, что различий между белками (и следовательно, генами) человека и мартышки гораздо меньше, чем между белками человека (или шимпанзе) и кошки.
Количественная оценка различий между видами по множеству генов позволяет построить генеалогическое древо, показывающее родство различных таксонов (видов, отрядов, семейств, классов), и определить относительное время их дивергенции. В основном это древо совпадает с теми, что были построены на основе морфологических, эмбриологических и палеонтологических данных.
Филогенетическое древо китопарнокопытных, построенное на основе сравнения генов. Ближайшим родственником китов и дельфинов является бегемот.
Ученые уже давно обнаруживали странное сходство между современными китами и копытными. Палеонтологические данные подтвердили, что предки китов по ряду признаков были похожи на предков копытных. Данные молекулярно-генетического анализа позволили точно определить положение китов на едином древе жизни. Ближайшим родственником китов оказался гиппопотам, который относится к отряду парнокопытных. Это значит, что общий предок китов и парнокопытных (гиппопотамов, коров, свиней) существовал в более близкое к нашим дням время, чем общий предок парно- и непарнокопытных (лошадей, тапиров, носорогов). В современной систематике киты и парнокопытные объединяются в один отряд китопарнокопытных
Молекулярные данные предоставляют нам самые убедительные свидетельства эволюции. Опираясь на эти данные, мы можем реконструировать историю жизни на Земле, показать, что все современные виды связаны неразрывными нитями родства, вся жизнь на Земле представляют собой единое филогенетическое древо.
Мутации – источник генетической изменчивости популяций.
Частота возникновения мутаций. Эволюция была бы невозможной, если бы генетические программы воспроизводились абсолютно точно. Как вы знаете, копирование генетических программ – репликация ДНК – происходит с высочайшей, но не абсолютной точностью. Изредка возникают ошибки – мутации. Частота мутаций не одинакова для разных генов, для разных организмов. Она возрастает, иногда очень резко, в ответ на воздействие внешних факторов, таких как ионизирующая радиация, некоторые химические соединения, вирусы и при изменениях внутреннего состояния организма (старение, стресс и т.п.).
Средняя частота мутаций у бактерий оценивается как 10-9 на ген на клетку за поколение. У человека и других многоклеточных она выше и составляет 10-5 на ген на гамету за поколение. Иными словами только в одной из 100 тысяч гамет ген оказывается измененным. Казалось бы, это ничтожно малая величина. Следует помнить, однако, что генов в каждой гамете очень много. По современным оценкам геном человека содержит около 30 тысяч генов. Следовательно, в каждом поколении около трети человеческих гамет несут новые мутации по какому-нибудь гену.
Таким образом, несмотря на чрезвычайную редкость каждой отдельной мутации, в каждом поколении появляется огромное количество носителей мутантных генов. Благодаря мутационному процессу генотипы всех организмов, населяющих Землю, постоянно меняются; появляются все новые и новые варианты генов (аллели), создается огромное генетическое разнообразие, которое служит материалом для эволюции.
Вредные, полезные и нейтральные мутации. Мутации различаются по своим фенотипическим эффектам. Большинство мутаций, по-видимому, вовсе никак не сказываются на фенотипе. Их называет нейтральными мутациями. Большой класс нейтральных мутаций обусловлен заменами нуклеотидов, которые не меняют смысла кодонов. Такие замены называют синонимическими. Например, аминокислота аланин кодируются триплетами ГЦУ, ГЦЦ, ГЦА и ГЦГ. Если в результате мутации ГЦУ превращается в ГЦЦ, то белок, синтезированный по измененной программе, остается тем же самым. Если мутация изменяет смысл кодона (несинонимическая мутация) и одна аминокислота заменяется другой, это может привести к изменению свойств белка.
Большинство несинонимических мутаций оказывается вредными. Они нарушают скоординированное в ходе предшествующей эволюции взаимодействие генетических программ в развивающимся организме, и приводят либо к его гибели, либо к тем или иным отклонениям в развитии. Только очень малая доля вновь возникающих мутаций может оказаться полезной.
Следует помнить, однако, насколько условна эта классификация. Полезность, вредность, или нейтральность мутации зависит от условий, в которых живет организм. Мутация нейтральная или даже вредная для данного организма и данных условиях, может оказаться полезной для другого организма и в других условиях, и наоборот. Жуки и комары не могли знать заранее, что люди изобретут ДДТ и другие инсектициды и подготовить мутации защиты. Тем не менее, эти мутации возникали – они обнаруживаются даже в тех популяциях насекомых, которые с инсектицидами не встречались. В то время, когда насекомые не сталкивались с инсектицидами, эти мутации были нейтральными. Но как только люди стали применять инсектициды – эти мутации стали не просто полезными, они стали ключевыми для выживания. Те особи, которым по наследству досталась такая мутация, вовсе не нужная их родителям, жившим в доинсектицидную эру, приобрели колоссальное преимущество перед теми, кто такой мутации не имел.
Естественный отбор «оценивает» вредность и полезность мутаций по их эффектам на выживание и размножение мутантных организмов в конкретных экологических условиях. При этом вредность мутации, как правило, обнаруживается немедленно, а ее полезность часто определяется задним числом: мы называем полезными те мутации, которые позволяют популяциям адаптироваться к изменяющимся условиям среды.
Чем сильнее фенотипический эффект мутации, тем вреднее такая мутация, тем выше вероятность того, что такая мутация будет отбракована отбором. Как правильно отметил Ч.Дарвин, природа не делает скачков. Ни одна сложная структура не может возникнуть в результате мутации с сильным фенотипическим эффектом. Новые признаки не возникают мгновенно, они формируется медленно и постепенно путем естественного отбора случайных мутаций со слабыми фенотипическими эффектами, которые чуть-чуть изменяют старые признаки.
Мутации случайны и не направлены. Принципиальным положением мутационной теории является утверждение, что мутации случайны и не направлены. Под этим подразумевается, что мутации изначально не адаптивны. Применение инсектицидов не ведет к направленному возникновению мутаций устойчивости к ним у насекомых. Инсектициды могут приводить к общему повышению частоты мутаций, в том числе и мутаций в генах устойчивости к ним, в том числе и таких мутаций, которые эту устойчивость повышают. Но на одну такую «адаптивную» мутацию в «нужном» гене возникают десятки тысяч любых других – нейтральных и вредных - мутаций в генах, которые не имеют никакого отношения к устойчивости к инсектицидам.
Организм не может знать, какие мутации будут полезны в следующем поколении. Нет и не может быть механизма, который бы обеспечивал направленное появление полезных для организма мутаций. Это утверждение следует из всего того, что мы знаем о принципах кодирования, реализации и передачи генетической информации. Мы уже говорили о том, что ДНК – это не чертеж, а рецепт создания организма. Говорят, что генотип определяет фенотип. Не следует понимать эту фразу буквально. Генотип определяет не сам фенотип, а последовательности биохимических и морфогенетических реакций, которые, взаимодействуя друг с другом, определяют развитие фенотипических признаков. Изменения генотипа влекут за собой изменения фенотипа, но не наоборот. Как бы не менялся фенотип организма в ответ на воздействия внешней среды – его изменения не могут привести к изменению генов, которые этот организм передаст следующему поколению.
Роль хромосомных и геномных мутаций в эволюции. Все перечисленные выше характеристики верны для всех типов мутаций – генных, хромосомных и геномных. Однако, такие геномные и хромосомные мутации как полиплоидия (кратное увеличение количества хромосом) и дупликации (удвоения определенных участков хромосом) играют особую роль в эволюции. Это связано с тем, что они увеличивают количество генетического материала и тем самым открывают возможность возникновения новых генов с новыми свойствами.
Расшифровка генома человека и других организмов показала, что многие гены и участки хромосом представлены в нескольких копиях. К ним относятся множество генов, отвечающих за синтез рибосомной РНК, гистонов (белков, участвующих в упаковке ДНК в хромосомах) и многих других. Таких генов нужно много для того, чтобы обеспечить высокий уровень синтеза, контролируемых ими продуктов. Следует ли из этого, что множественные копии этих генов возникли для этого? Конечно же, нет. Удвоение всего генома или его отдельных участков происходило случайно. При этом удваивались не только эти гены, но и многие другие. Естественный отбор, однако, «поступал» с этим лишними копиями по-разному. Некоторые копии оказались полезными, и естественный отбор поддерживал их в популяциях. Другие оказались вредными, поскольку «больше - не всегда лучше». В этом случае отбор или отбраковывал носителей таких копий, или способствовал размножению таких особой, у которых излишние копии генов терялись в результате других хромосомных мутаций – делеций. Были, наконец, и нейтральные копии, присутствие которых никак не сказывалось на приспособленности их носителей.
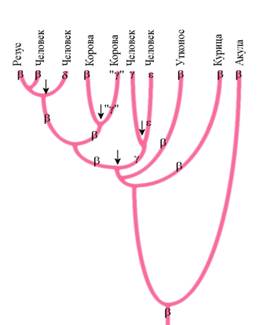
|
| Филогенетическое древо глобиновых генов. Ген глобина в ходе эволюции несколько раз дуплицировался (отмечено стрелками). Его добавочные копии затем приобретали новые свойства и функции. Из гена бета-глобина общего предка возникли гены гамма-, дельта-, эпсилон-глобинов – белков, которые выполняют иные функции, чем бета-глобин. |
Эти лишние копии становились резервом эволюции. Мутации в таких «резервных генах» не так строго отбрасывались отбором, как мутации в основных, уникальных генах. Резервным генам было «позволено» меняться в более широких пределах. Со временем они могли приобретать новые функции и становиться все более и более уникальными. Ярким примером последствий такого процесса является многочисленное и разнообразное семейство генов глобинов млекопитающих. Анализ последовательности нуклеотидов в этих генах показывается, что все они произошли в результате серии последовательных удвоений одного-единственного гена. За каждым удвоением следовало накопление случайных мутаций и постепенное изменение их функций, синтезируемых ими белков.
Когда мы сравниваем кариотипы разных видов млекопитающих, мы обнаруживаем, что в ходе эволюции этих видов происходили и закреплялись и другие хромосомные мутации, такие как транслокации и инверсии. Кариотип человека отличается от шимпанзе и других антропоидов одной транслокацией и несколькими инверсиями. За десятки миллионов лет независимой эволюции в кариотипах человека и землеройки возникли и закрепились десятки различных транслокаций и инверсий. Эти хромосомные перестройки не могли бы закрепиться, если бы они резко нарушали жизнеспособность или плодовитость их носителей.
В результате транслокаций и инверсий меняется взаимное расположение генов и, следовательно, характер их взаимодействия. В настоящее время хорошо известно, какую важную роль в проявлении генов играют их регуляторные элементы. Эти элементы, как правило, находятся в тех же хромосомах, что и контролируемые гены, но часто на большом расстоянии от них. Отрыв гена от его регуляторного элемента, обусловленный инверсией или транслокаций, или соединение этого гена с чужим регуляторным элементом может приводить к значительным изменениям в функции гена – времени его проявления в развитии, типе клеток, в которых этот ген активен, в количестве синтезируемого белка. К таким же последствиям может приводить и перемещение мобильных генетических элементов, которые могут захватывать и переносить с места на место регуляторные элементы.
В геноме обнаружены участки, где довольно часто происходят разрывы хромосом, ведущие к образованию хромосомных перестроек. Найдены и участки преимущественной локализации мобильных генетических элементов. Интересно, что во многих случаях это одни и те же участки. Таким образом, мы можем говорить о неслучайном распределении этих участков по геному. Однако, и как все остальные мутации, хромосомные перестройки и перемещения мобильных элементов случайны. Они случайно меняют функции генов, находящихся вблизи точек разрывов, они случайно распределяют гены по геному. Они приводят к тому, что возникает множество новых «коалиций» генов, а приспособительная ценность этих «коалиций» оценивается отбором.
Мутационный процесс является важнейшим фактором эволюции. Мутационный процесс изменяет гены и порядок их расположения в хромосомах и тем самым увеличивает генетическое разнообразие популяций. Он создает избыточные копии генов и тем самым открывает возможность усложнения организмов. Мутации возникают случайно и не направленно. Адаптивная ценность каждой мутации не постоянна. Она определяется взаимодействием мутантного аллеля с другими генами организма и с условиями среды, в которой развивается и живет мутантный организм.
Мутационный процесс, создавая новые аллели генов, изменяет частоты аллелей в популяциях. Пусть частота спонтанного мутирования аллеля B (например, черной окраски лисиц) в аллель b ( красной окраски) равна 10-5 на гамету за поколение.Тогда частота аллеля b в популяции будет медленно, но неуклонно возрастать в каждом поколении на одну десятитысячную, если этому возрастанию не будут препятствовать или способствовать другие факторы эволюции. В принципе, благодаря только мутационному процессу новый аллель может практически полностью вытеснить старый аллель из популяции. Однако, это будет происходить крайне медленно. Для того чтобы снизить частоту аллеля B от 0,50 до 0,49 нужно 2000 поколений. Но дальше процесс пойдет еще медленнее. На снижение от 0,10 до 0,09 – уйдет 10000 поколений. Чем ниже частота аллеля, тем медленнее она будет изменяться за счет мутаций. Если бы мутационный процесс был единственным фактором эволюции, то сама эволюция происходила гораздо медленнее, чем на самом деле. Частоты генов в популяциях меняются не только и не столько за счет мутационного процесса, но благодаря действию других факторов эволюции.
– Конец работы –
Эта тема принадлежит разделу:
Возникновение и развитие эволюционной биологии.
На сайте allrefs.net читайте: Возникновение и развитие эволюционной биологии....
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Палеонтологические свидетельства эволюции.
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.211 сек.
Новости и инфо для студентов