рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Биология
- /
- Топологический подход
Реферат Курсовая Конспект
Топологический подход
Топологический подход - раздел Биология, СИНЕРГЕТИКА ДЛЯ БИОЛОГОВ Биологи Традиционно И Повсеместно Используют Язык Геометрии Э...
Биологи традиционно и повсеместно используют язык геометрии Эвклида для описания структур и динамики формообразования на субклеточном, клеточном, тканевом уровнях, в индивидуальном развитии организмов и в эволюционных перестройках. Однако для такого рода описаний применим и язык топологии, поскольку именно топология рассматривает наиболее общие пространственные свойства объектов. Необходимость применения топологического языка в биологии неоднократно декларировалась такими выдающимися математиками и биологами как Р. Том и К. Уоддингтон. Плодотворность топологического анализа и моделирования уже осознана; разработана методология, позволяющая описать топологические паттерны морфогенетических процессов на всех уровнях исследования живых организмов, поставлен и решается вопрос о топологической обусловленности и топологических ограничениях биологического морфогенеза (Преснов, Исаева, 1985; Исаева, Преснов, 1990).
К настоящему времени топологический подход наиболее эффективно использован в молекулярной биологии для описания и анализа форм ДНК; топологический анализ структуры ДНК впервые был предпринят Ф. Криком (Crick, 1976) и с тех пор был существенно продвинут. Топология привлекалась также для анализа пространственной организации мембранных систем клетки и их динамики как на молекулярном, так и на субклеточном уровнях. Поле же исследований топологии клеток и организмов в онтогенезе и эволюции, примыкающее к междисциплинарной области исследований хаоса и самоорганизации, остается в мировой науке почти не занятым.
Для топологического описания не существенны ни геометрическая форма (линейные и угловые размеры, кривизна линий и поверхностей), ни частные морфологические детали организации объекта; принимаются во внимание лишь топологически инвариантные характеристики. При описании внешней формы организмов, их тканевых и органных систем такой топологической характеристикой, определяемой корректно и однозначно, может служить род поверхности (p), который визуализируется посредством простейших канонических поверхностей – шара, тора и различных “кренделей” (рис. 37).
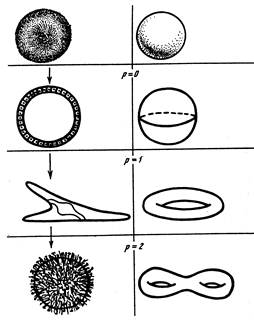
|
Рис. 37. Перестройки рода поверхности в онтогенезе морского ежа
(Преснов, Исаева, 1991)
Поскольку при таком подходе внешняя форма биологических объектов моделируется гладкими замкнутыми поверхностями, морфогенез многоклеточных животных может быть представлен как последовательность топологических перестроек их эпителизованных поверхностей. При этом к анализу пространственной организации эпителиальных слоев применяется теорема элементарной топологии, согласно которой любая замкнутая ориентируемая поверхность в трехмерном пространстве гомеоморфна (т.е. топологически эквивалентна) сфере с определенным числом (p) ручек (см. Милнор, Уоллес, 1972; Матвеев, Фоменко, 1991). У биологических объектов топологические ручки обычно представлены сквозными каналами (пищеварительным и т.д.). Любой покрытый эпителием сквозной канал или сквозное отверстие рассматривается в качестве эквивалента топологической ручки; топологические перестройки поверхности биологических объектов реализуются путем локальных разрушений и последующих срастаний эпителиальных слоев; преобразуемые без разрыва объекты топологически эквиваленты.
У низших Bilateria род поверхности p = 0. Дальнейшие преобразования привели к появлению сквозного кишечника с родом поверхности p = 1. Тип тора - формы со сквозным кишечником - оказался устойчивой топологической структурой. Новые топологические паттерны Bilateria обычно возникают на базе других систем - целомической, дыхательной и половой (Преснов, Исаева, 1985; Исаева, Преснов, 1990; Чернышев и др., 2001). Тем не менее в отдельных группах могут встречаться топологические усложнения переднего отдела пищеварительного тракта. У асцидий, ведущих прикрепленный фильтрующий образ жизни, род поверхности может быть неопределенно велик (до нескольких тысяч), а число щелей в глотке варьирует даже у разных особей одной колонии, т.е. наблюдаются элементы топологического хаоса. Напротив, у активных хордовых происходит уменьшение и стабилизация числа жаберных щелей и, соответственно, рода поверхности.
Общая схема эволюции рода поверхности многоклеточных представлена на рис. 38.

|
Рис. 38. Схема эволюционных преобразований поверхности организма многоклеточных животных (Преснов, Исаева, 1985)
Топологические усложнения развиваются на базе систем, функционально тесно связанных с внешней средой и фактически распределяющих ее в организме с увеличением площади поверхности раздела наружной среды и внутренней среды организма. К таким системам относятся, например, амбулакральная система иглокожих и трахейная система наземных членистоногих. Род поверхности иглокожих можно выразить как p = 1 + n, где n - род поверхности амбулакральной системы (обычно равен 1, реже доходит до 5).
Усложненные топологические паттерны можно условно разделить на две группы - упорядоченные и неупорядоченные. Формы с упорядоченными топологическими паттернами имеют стабильный род поверхности. К таким формам относится большинство Bilateria, а также многие гидромедузы и гребневики. Формы с неупорядоченными топологическими паттернами имеют большой, нестабильный и неопределенный род поверхности. Полностью неупорядоченные паттерны свойственны лишь губкам. Топологическая неупорядоченность всегда развивается на основе фракталоподобных систем эпителиальных каналов. Хаотическая динамика процессов морфогенеза, проявляющаяся у высших животных в хаотизации лишь конечных этапов морфогенеза квазифрактальных структур, более выражена у низших многоклеточных.
Топологическая обусловленность увеличения рода поверхности у многоклеточных животных, возможно, заключается в неизбежности существование особенностей векторного поля на поверхности зародыша или многоклеточного сферического (с топологической точки зрения) организма (Преснов, Исаева, 1990, 1991). Эти особенности векторного поля на сфере – сингулярные точки – могут определять локализацию впячивания (инвагинации) поверхности организма. Во время встречи сопряженных точек происходит взаимное уничтожение топологического «заряда» (топологического индекса) и изменение топологии поверхности. Таким образом, изменение рода поверхности является следствием неизбежной неоднородности векторного морфогенетического поля сферической поверхности организма, что открывает реальные возможности для более строгого топологического анализа этого процесса.
Топологические ограничения биологического морфогенеза наиболее ясно могут быть выявлены в модельных системах in vitro. Например, в культуре миогенных клеток наблюдается фазовый переход клеточной системы - от отдельных беспорядочно расположенных миобластов к формированию клеточных потоков, что обусловлено взаимодействиями клетка-клетка и клетка-субстрат, создающих локальный порядок расположения клеток. При слиянии миобластов в миосимпласты с формированием миотуб вдоль траекторий полей направлений клеток происходит стабилизация возникшего паттерна с отчетливой визуализацией топологических сингулярностей полей направлений (рис. 39).
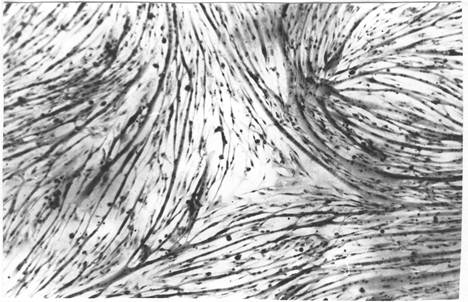
|
Рис. 39. Топологические сингулярности полей направлений в миогенной культуре (Исаева, 1994)
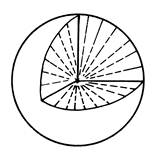
Путем контактной ориентации клетка-клетка и клетка-субстрат осуществляется трансляция локального, ближнего порядка клеточных взаимодействий в дальний, глобальный порядок клеточных полей. Ограниченное число топологических особенностей полей направлений, возможных на плоскости, диктует и соответствующие ограничения планарного морфогенеза - в однослойной культуре (рис. 40).

|
Рис. 40. Сингулярности полей направлений на плоскости
(Исаева, Преснов, 1990; по: Минеев, 1982)
Поля направлений осей удлиненных клеток на субклеточном уровне детерминируются полями направлений структурных элементов цитоскелета, определяющих осевую ориентацию поляризованных клеток. Фибриллярные или тубулярные структуры цитоскелета (цитоматрикса) образованы удлиненными молекулярными комплексами, состоящими главным образом из полимеров актина, тубулина, белков промежуточных филаментов, обладающих жидкокристаллическими свойствами. Система топологических сингулярностей, или дефектов жидкокристаллической структуры – основа морфофункциональной организации цитоматрикса, выполняющего опорную, двигательную и интегрирующую клетку функции. К дефектам такого рода применимо высказывание Ларошфуко, приведенное в книге П. Де Жена о жидких кристаллах (1977, с. 145): «Есть недостатки, которые на деле блистают ярче, чем сами добродетели».
В свою очередь, сингулярности структуры цитоматрикса в определенной мере обусловлены самоорганизацией молекул в жидких кристаллах. Для примера рассмотрим самый простой случай так называемого нематического жидкого кристалла, упорядоченность которого определяется тенденцией удлиненных молекул выстраиваться параллельно друг другу (рис. 41). Преимущественное направление длинных осей молекул описывается единичным вектором; распределение таких отрезков создает поле направлений. В двумерном нематике, как и в однослойной клеточной культуре, возможно существование лишь ограниченного числа топологических сингулярностей (рис. 40).
В капле нематического жидкого кристалла влияние граничных условий и поверхностного натяжения на расположение молекул приводит к возникновению точечной трехмерной сингулярности (рис. 42), «ежа» (либо двух особенностей на поверхности).

|
Рис. 41. Ориентация молекул нематического жидкого кристалла
Таким образом, в жидких кристаллах можно наблюдать как структурную самоорганизацию молекул, так и топологические ограничения морфогенеза в этой системе. Сходство точечной трехмерной сингулярности в капле нематика (рис. 42) с центром организации микротрубочек эукариотических клеток не случайно: структуры цитоскелета обладают свойствами жидких кристаллов.
|
Рис. 42. Объемная точечная сингулярность в капле нематика
(Исаева, Преснов, 1990; по: Рожков, 1986)
К цитоматриксу (рис. 43) применимы также представления о перколяции – образовании связной сети, связного кластера с изменением свойств системы, фазовым переходом (Де Жен, 1982; Исаева, Преснов, 1990; рис. 44). Итак, определенные топологические ограничения биологического морфогенеза неизбежны и неустранимы.

|
Рис. 43. Организация цитоматрикса
(Исаева, Преснов, 1990; по: Porter, 1987)

|
Рис. 44. Узлы решетки ниже порога перколяции (слева) и вблизи него (Шредер, 2001)
– Конец работы –
Эта тема принадлежит разделу:
СИНЕРГЕТИКА ДЛЯ БИОЛОГОВ
На сайте allrefs.net читайте: Дальневосточный государственный университет...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Топологический подход
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.02 сек.
Новости и инфо для студентов