Репликация линейных молекул ДНК с внутренней инициацией и внесением разрыва в цепь
Лекция 2
II.Репликация линейных молекул ДНК с внутренней инициацией и внесением разрыва в цепь.
Репликация вирусов с линейными одноцепочечными ДНК
Parvaviridae (parva - мелкий). Поражают насекомых и животных, в т.ч. и человека. Не имеют липопротеидной… Парвавирусы животных можно разделить на две группы:II.
| |||||||
 | |||||||
 | |||||||
 | |||||||
 |
|
|
|
|
|
|
Одноцепочечная ДНК далее может вновь вступить в репликативный цикл или, при накоплении белков может упаковываться в вирусные частицы.
Ферменты и вспомогательные белки, участвующие в репликации парвавирусов:
DNApolδ мультисубъединичная высокпроцессивная ДНК-полимераза эукариот, RPA (аналог SSB прокариот) – белок связывающийся с одноцепочечными участками ДНК и стабилизирующий расплетенное состояние дуплекса ДНК, RF-С – белковый комплекс, загружающий фактор процессивности ДНК-полимеразы (аналог γ-комплекса прокариот), PCNA – гетеротримерный фактор процессивности ДНК-полимеразы (аналог β-субъединицы ДНК-полимеразы прокариот). Заметьте, что праймаза не участвует в репликации парвавирусов.
Кольцевые формы:
|
 | |||||
 | |||||
 |
Шпилечную структуру может узнать Rep-белок и внести разрыв. Проблема в том, что шпилька есть только у одного конца, а на другом конце инвертированного повтора нет. Как достроить 3’ конец молекулы? Решение проблемы заложено в структуре ДНК: после синтеза дочерней цепи вытесненная цепь образует кольцо и 3’-конец достраивается по 5’-концу. Образуется полноценная геномная ДНК.
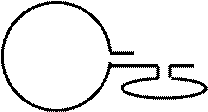 |  | ||||
 | |||||
|
|
Геном аденоассоциированных вирусов состоит из двух генов. Однако при экспрессии этих генов за счет альтернативной инициации транскрипции и альтернативного сплайсинга образуются группы Rep-белков и белков оболочки.
Rep белок, как уже было упомянуто, является транскрипционным фактором, и последовательность, которую он узнает, находится внутри генома вируса. Недалеко есть сайт, подобный TRS, в который Rep-белок может случайно внести разрыв. Если такой разрыв будет внесен в кольцевую двуцепочечную форму, то ее репликация будет осуществляться по схеме, не отличающейся от схемы разматывающегося рулона.

Благодаря инверсии, возникающей при переносе шпильки, возможно образование нескольких геномов с различной ориентацией концевых участков. В случае (+) цепей таких вариантов существует 4, еще 4 для (–) цепей, итого 8.
|
Аденоассоциированные вирусы могут встраиваться в геном животных. Например, в геноме человека существует около 200000 тысяч возможных сайтов, но встраивание вируса происходит на определенном участке XIX хромосомы человека, потому что необходимо наличие в определенных положениях поледовательностей, способных служить TRS и RBE сайтами.
Для вырезания провируса необходима транскрипция вирусной ДНК, однако Rep-белок является репрессором транскрипции. Превратить его в активатор транскрипции могут белки аденовирусов, поэтому переход от лизогении к литическому циклу у аденоассоциированных вирусов возможен лишь при коинфекции аденовирусами.
Участки связывания Rep-белка можно изменить с помощью генноинженерных методов таким образом, что становится возможным вырезание провируса при участии клеточных резольваз, узнающих крестообразные структуры.
 | |
 |
Аденоассоциированные вирусы пытаются использовать в генной инженерии.
Автозатравочный механизм репликации автономных парвавирусов
В вирионы включаются преимущественно (-) цепи ДНК. В отличие от аденоассоциированных вирусов концы различаются по структуре. На 5’-конце находится…Репликация вирусов с линейными двуцепочечными ДНК
Poxviridae
Неполная комплементраность цепей и наличие инверсии концов позволяют предположить, что репликация поксвирусов происходит по схеме, описанной для…III. Терминальная инициация репликации при помощи нуклеотид-белковой затравки