рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Биология
- /
- Строение и типы синапсов.
Реферат Курсовая Конспект
Строение и типы синапсов.
Строение и типы синапсов. - раздел Биология, Реферат по теме: Формирование черепномозговых и спинномозговых нервов и закономерности их ветвления. Строение рефлекторной дуги. Строение и типы синапсов. Виды нейромедиаторов. Переход Возбуждения С Нервного Волокна На Иннервируемую Им Клетку- Нервную, М...
Переход возбуждения с нервного волокна на иннервируемую им клетку- нервную, мышечную, секреторную- осуществляется при участии синапсов.
Синапсы- (от греч. synapsis- соединение, связь)- особый тип прерывистых контактов между клетками, приспособленных для односторонней передачи возбуждения или торможения от одного элемента к другому. Делят их в зависимости от локализации (центральные и периферические), функции (возбуждающие и тормозные),способа передачи возбуждения (химические, электрические, смешанные), природы действующего агента (холинергические или адренергические).
Синапсы могут быть между двумя нейронами (межнейронные), между нейроном и мышечным волокном (нервно-мышечные), между рецепторными образованиями и отростками чувствительных нейронов(рецепторно-нейронные), между отростком нейрона и другими клетками (железистыми, ресничными)
Основными компонентами синапса являются:пресинаптическая часть (обычно утолщенное окончание пресинаптического аксона), постсинаптическая часть (участок клетки, к которому подходит пресинаптическое окончание) и разделяющая их синаптическая щель (в синапсах с электрической передачей она отсутствует)
В простейшем типе синапса клетка иннервируется только одним волокном (аксоном). Так, в нервно-мышечном синапсе каждое мышечное волокно иннервируется аксоном одного двигательного нейрона. В сложных синапсах, например у клеток головного мозга, количество оканчивающихся аксонов может исчисляться несколькими тысячами.
Скелетные мышцы иннервируются волокнами соматической нервной системы, т.е. отростками нервных клеток (мотонейронов). расположенных в рогах спинного мозга или ядрах черепных нервов. Каждое двигательное волокно в мышце ветвится и иннервирует группу мышечных волокон. Концевые веточки нервных волокон (диаметром 1-1,5 мкм) лишены миелиновой оболочки, покрыты аксоплазматической мембраной с утолщениями и имеют расширенную колбовидную форму. Пресинаптическое окончание содержит митохондрии (поставщики АТФ), а также множество субмикроскопических образований – синаптических пузырьков (везикул) диаметром около 50 нм. Пузырьки более многочисленны в области утолщений пресинаптической мембраны.
Пресинаптические окончания аксона образуют синаптические соединения со специализированной областью мышечной мембраны (см. рис. 18). Последняя формирует углубления, складки, увеличивающие площадь поверхности постсинаптической мембраны и соответствующие утолщениям пресинаптической мембраны. Ширина синаптической щели составляет 50-100нм.
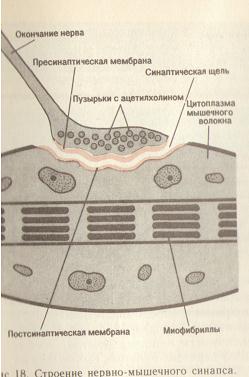
Область мышечного волокна, участвующую в образовании синапса, т.е. постсинаптическую часть контакта, называют концевой двигательной пластинкой или обозначают весь нервно-мышечный синапс.
Описанная электронно-микроскопическая картина является типичной для синапсов химической природы. Передатчиком возбуждения здесь служит посредник (медиатор)- ацетилхолин. Когда под действием нервного импульса (потенциала действия) происходит деполяризация мембраны нервного окончания, синаптические пузырьки вплотную сливаются с ней и их содержимое выбрасывается в синаптическую щель. Этому способствует повышение внутри окончания концентрации ионов кальция, поступающих извне по электровозбудимым кальциевым каналам.
Ацетилхолин выбрасывается порциями (квантами) по 4*10 молекул, что соответствует содержимому нескольких пузырьков. Один нервный импульс вызывает синхронное выделение 100-200 порций медиатора менее чем за 1 мс. Всего же запасов ацетил холина в окончании хватает на 2500-5000 импульсов. Таим образом, основное назначение пресинаптической части контакта состоит в регулируемом нервным импульсом выбросе медиатора ацетилхолина в синаптическую щель. Нервно-мышечный синапс является, холинэнергическим. Токсин ботулизма в следовых количествах блокирует освобождение ацетилхолина в синапсах и вызывает мышечный паралич.
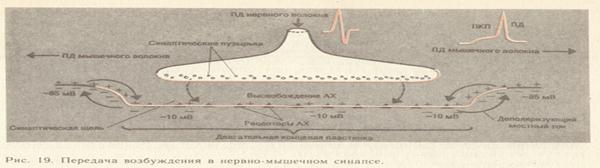
Молекулы ацетилхолина диффундируют через щель и достигают внешней стороны постсинаптической мембраны, где связываются со специфическими рецепторами- молекулами липопротеиновой природы. Число рецепторов составляет примерно 13000 на 1 мкм ;они отсутствуют в других участках мышечной мембраны. Взаимодействие медиатора с рецепторным белком (двух молекул ацетилхолина с одной молекулой рецептора) вызывает изменение конформации последнего и "открытие ворот" хемовозбудимых ионных каналов. В результате происходит перемещение ионов и деполяризация постсинаптической мембраны от -75до-10 мВ. Возникает потенциал концевой пластинки (ПКП), или возбуждающий постсинаптический потенциал (ВПСП). Последний термин применим ко всем типам химических синапсов, в том числе межнейронным.

Время от момента появления нервного импульса в пресинаптическом окончании до возникновения ПКП называется синаптической задержкой. Она составляет 0,2-0,5 мс.
Поскольку хемовозбудимые каналы не обладают электровозбудимостью, "запальная" деполяризация мембраны не вызывает дальнейшего увеличения числа активируемых каналов, как это имеет место в аксоплазматической мембране. Величина ПКП зависит от числа молекул ацетилхолина, связанных постсинаптической мембраной, т.е. в отличие от потенциала действия ПКП градуален. Амплитуда его зависит и от сопротивления мышечной мембраны (тонкие мышечные волокна имеют более высокий ПКП). Некоторые вещества, например яд кураре, связываясь с рецепторными белками, препятствуют действию ацетилхолина и подавляют ПКП. Известно, что на каждый импульс от мотонейрона в мышце всегда возникает потанцеал действия. Это обусловлено тем, что пресинаптическое окончание выделяет определенное количество квантов медиатора и ПКП всегда достигает пороговой величины.
Между деполяризованной ацетилхолином постсинаптической мембраной и граничащей с ней мембраной скелетного мышечного волокна возникают местные токи, вызывающие потенциалы действия, распространяющиеся по всему мышечному волокну. Последовательность событий, ведущих к возникновению потенциала действия, изображена на рисунке 19. Для восстановления возбудимости постсинаптической мембраны необходимо исключение деполяризующего агента- ацетилхолина. Эту функцию выполняет локализованный в синаптической щели фермент ацетилхолинэстераза, которая гидролизует ацетилхолин до ацетата и холина. Проницаемость мембраны возвращается к исходному уровню и мембрана реполяризуется. Этот процесс идет очень быстро: весь выделившийся в щель ацетилхолин расщепляется за 20 мс.
Некоторые фармакологические или токсические агенты (алкалоид физостигмин, органические фторфосфаты), ингибируя ацетилхолинэстеразу, удлиняют период ПКП, что вызывает "залпы" потенциалов действия и спастические сокращения мышцы в ответ на одиночные импульсы мотонейронов.
Образовавшиеся продукты расщепления- ацетат и холин- большей частью транспортируются обратно в пресинаптические окончания, где используются в синтезе ацетилхолина при участии фермента холин-ацетилтрансферазы (рис. 20).
Типы синапсов:
Электрические синапсы. В настоящее время признают, что в ЦНС имеются электрические синапсы. С точки зрения морфологии электрический синапс представляет собой щелевидное образование (размеры щели до 2 нм) с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно перескакивают через такой щелевидный контакт и возбуждают, т.е. индуцируют генерацию ПД второй клетки. В целом, такие синапсы (они называются эфапсами) обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т. к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя заставить эффекторную клетку (клетку, которая управляется через данный синапс) тормозить свою активность. Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
Химические синапсы. По строению химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы), которая заполнена химическим веществом - медиатором. В синапсе различают пресинаптический элемент, который ограничен пресинаптической мембраной, а также внесинаптическую область и синаптическую щель, величина которой составляет в среднем 50 нм. В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка - это синапс между нейронами, концевая пластинка - это постсинаптическая мембрана мионеврального синапса, моторная бляшка - это пресинаптичсское окончание аксона на мышечном волокне.
– Конец работы –
Эта тема принадлежит разделу:
Реферат по теме: Формирование черепномозговых и спинномозговых нервов и закономерности их ветвления. Строение рефлекторной дуги. Строение и типы синапсов. Виды нейромедиаторов.
Министерство сельского хозяйства Российской Федерации... Красноярский Государственный Аграрный университет... Кафедра quot физиологии и этологии животных quot...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Строение и типы синапсов.
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.018 сек.
Новости и инфо для студентов