Сцепленное наследование генов
Дальнейшие исследования генетиков показали, что законы Менделя о независимом наследовании признаков при дигибридном скрещивании применимы лишь тогда, когда разные гены располагаются в разных парах гомологичных хромосом. В том случае, если два гена находятся в одной паре гомологичных хромосом, расщепление в потомстве гибридов будет другим. У любого организма генов значительно больше, чем хромосом. Например, у человека имеется около миллиона генов, а хромосом всего 23 пары. Следовательно, в одной хромосоме размещается в среднем несколько тысяч генов. Гены, расположенные в одной хромосоме, называют сцепленными. Все гены этой хромосомы образуют группу сцепления, которая при мейозе обычно попадает в одну гамету.
Значит, гены, входящие в одну группу сцепления, не подчиняются закону независимого наследования, а при дигибридном скрещивании вместо ожидаемого расщепления по фенотипу в соотношении 9:3:3:1 дают соотношение 3:1, как при моногибридном скрещивании.
 Закономерности сцепленного наследования были установлены американским биологом Томасом Морганом (1866-1945). В качестве объекта он использовал плодовую муху дрозофилу. У дрозофилы окраску тела и длину крыльев определяют следующие пары аллелей: А - серое тело, а - черное тело, В - длинные крылья, b - зачаточные крылья. Гены, отвечающие за окраску тела и длину крыльев, находятся в одной паре гомологичных хромосом и наследуются сцепленно.
Закономерности сцепленного наследования были установлены американским биологом Томасом Морганом (1866-1945). В качестве объекта он использовал плодовую муху дрозофилу. У дрозофилы окраску тела и длину крыльев определяют следующие пары аллелей: А - серое тело, а - черное тело, В - длинные крылья, b - зачаточные крылья. Гены, отвечающие за окраску тела и длину крыльев, находятся в одной паре гомологичных хромосом и наследуются сцепленно.
При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой, имеющей черное тело и зачаточные крылья, все гибриды первого поколения имели серую окраску тела и длинные крылья .
При дальнейшем скрещивании между собой гибридных мух первого поколения в F2 не произошло ожидаемого расщепления по фенотипу 9:3:3:1. Вместо этого в F2 были получены мухи с родительскими фенотипами в соотношении примерно 3:1. Появление в F2 двух фенотипов вместо четырех позволило сделать вывод, что гены окраски тела и длины крыльев дрозофил находятся в одной хромосоме. Так был установлен закон Т.Моргана: гены, расположенные в одной хромосоме, наследуются совместно - сцепленно, то есть наследуются преимущественно вместе.
Однако при дигибридном скрещивании при сцепленном наследовании признаков не всегда появляются особи только двух фенотипов. Иногда появляются особи еще двух фенотипов с перекомбинацией (новым сочетанием) родительских признаков: серое тело - зачаточные крылья, черное тело - длинные крылья. (Особей с такими фенотипами немного - около 8,5% каждого типа.) Почему же нарушается сцепление генов и появляются особи с новыми фенотипами? Было установлено, что сцепление генов может быть полным и неполным.
 Полное сцепление наблюдается в том случае, если скрещиваются серый самец с длинными крыльями и самка с черным телом и зачаточными крыльями. Расщепление по фенотипу в этом случае будет 1:1, то есть наблюдается полное сцепление генов в одной хромосоме.
Полное сцепление наблюдается в том случае, если скрещиваются серый самец с длинными крыльями и самка с черным телом и зачаточными крыльями. Расщепление по фенотипу в этом случае будет 1:1, то есть наблюдается полное сцепление генов в одной хромосоме.
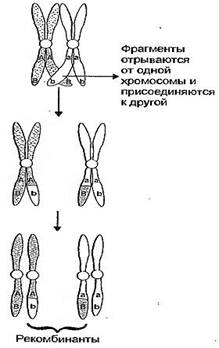
При скрещивании серой длиннокрылой самки с самцом, имеющим черное тело и зачаточные крылья, расщепление по фенотипу будет примерно 41,5:41,5:8,5:8,5, что характеризует неполное сцепление. Причина нарушения сцепления заключается в том, что в ходе мейоза происходит кроссинговер и гомологичные хромосомы обмениваются своими участками. В результате гены, расположенные в одной из гомологичных хромосом, оказываются в другой хромосоме. Возникают новые сочетания признаков. Вероятность кроссинговера зависит от расстояния между генами. Расстояние между генами измеряют в морганидах. 1 морганида равна 1% перекомбинированных признаков в потомстве.
У самцов дрозофил в мейозе кроссинговер не происходит, поэтому при скрещивании серого длиннокрылого самца дрозофилы с рецессивной самкой с черным телом и зачаточными крыльями сцепление будет полным. Неполное сцепление наблюдается в том случае, если самка гетерозиготна, а самец гомозиготен. В данном примере кроссинговер происходит примерно у 17% самок.
Таким образом, если не происходит перекреста хромосом и обмена генами, то наблюдается полное сцепление генов. При наличии кроссинговера сцепление генов бывает неполным. Благодаря перекресту хромосом возникают новые сочетания генов и признаков. Чем дальше друг от друга расположены гены в хромосоме, тем больше вероятность перекреста между ними и обмена участками хромосом.
Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности:
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
3. Гены расположены в хромосомах в определенной линейной последовательности;
4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом;
6. Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);
7. Каждый вид имеет характерный только для него набор хромосом - кариотип.