Трансформация
Феномен трансформации впервые был описан в 1928 г. Ф. Гриффитсом, обнаружившим превращение бескапсульного R-штамма пневмококков (Streptococcus pneumoniae) в штамм, образующий капсулу S-формы. Гриффитс ввел мышам одновременно небольшое количество авирулентных R-клеток и убитых нагреванием S-клеток. R-клетки были получены от штамма, капсульное вещество которого принадлежало к типу S II, а убитые нагреванием S-штаммы - к типу S III. Из крови погибших мышей были выделены вирулентные пневмококки с капсулой S III.
В 1944 г. О. Эвери, К. Мак-Леод, М. Мак-Карти установили природу трансформирующего фактора, показав, что ДНК, экстрагированная из инкапсулированных пневмококков, может трансформировать некапсулированные пневмококки в инкапсулированную форму. Таким образом, было доказано, что именно ДНК является носителем генетической информации.
Процесс трансформации может самопроизвольно происходить в природе у некоторых видов бактерий, B. subtilis, H. influenzae, S. pneumoniae, когда ДНК, выделенная из погибших клеток, захватывается реципиентными клетками. Процесс трансформации зависит от компетентности клетки-реципиента и состояния донорской трансформирующей ДНК.Компетентность - это спо-
собность бактериальной клетки поглощать ДНК. Она зависит от присутствия особых белков в клеточной мембране, обладающих специфическим аффинитетом к ДНК. Состояние компетентности у грамположительных бактерий связано с определенными фазами кривой роста. Состояние компетенции у грамотрицательных бактерий приходится создавать искусственным путем, подвергая бактерии температурному или электрошоку.
Трансформирующей активностью обладает только двунитевая высокоспирализованная молекула ДНК. Это связано с тем, что в клетку-реципиент проникает только одна нить ДНК, тогда как другая - на клеточной мембране - подвергается деградации с высвобождением энергии, которая необходима для проникновения в клетку сохранившейся нити. Высокая молекулярная масса трансформирующей ДНК увеличивает шанс рекомбинации, так как внутри клетки трансформирующая нить ДНК подвергается воздействию эндонуклеаз. Интеграция с хромосомой требует наличия гомологичных с ней участков у трансформирующей ДНК. Рекомбинация происходит на одной нити, в результате чего образуется гетеродуплексная молекула, одна нить которой имеет генотип реципиента, а другая - рекомбинантный генотип. Рекомбинантные трансформанты формируются только после цикла репликации (рис. 5.5).
В настоящее время этот метод является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
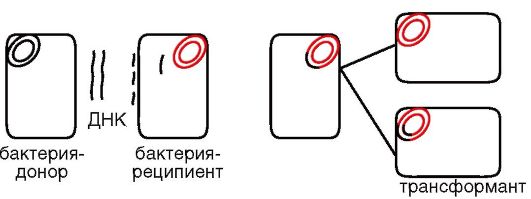 Рис. 5.5.Схема трансформации
Рис. 5.5.Схема трансформации