рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Биология
- /
- ТЕОРИЯ ЭВОЛЮЦИИ
Реферат Курсовая Конспект
ТЕОРИЯ ЭВОЛЮЦИИ
ТЕОРИЯ ЭВОЛЮЦИИ - раздел Биология, Теория Эволюции ...
ТЕОРИЯ ЭВОЛЮЦИИ
Представления об эволюции до Чарлза Дарвина
Эволюция протекает на всех уровнях организации живой материи и на каждом уровне характеризуется новообразованием структур и появлением новых… Проблемы происхождения и эволюции жизни на Земле принадлежали и принадлежат к… В основе современной теории эволюции лежит теория Ч. Дарвина. Но эволюционизм существовал и до Ч. Дарвина. Поэтому,…Ч. Дарвин и его теория эволюции
Творцом первой подлинно научной теории эволюции стал великий английский ученый Чарлз Роберт Дарвин (1809-1882). Главным трудом Ч. Дарвина является… Эволюционное учение Дарвина состоит из трех разделов, а именно: совокупность… Доводы в пользу того, что эволюция действительно имеет место, Ч. Дарвин черпал из разных наук. Наиболее убедительные…Современные представления о происхождении жизни
В современной теории эволюции значительное место занимают вопросы происхождения жизни на Земле, ибо в рамках теории эволюции происхождение жизни… Общепризнанной теорией происхождения жизни является теория, впервые… Предполагают, что Земля сформировалась 4,5-5 млрд лет назад из облаков космической пыли. Она увеличивалась по массе в…Ход, главные направления и доказательства эволюции
Поскольку эволюция представляет собой процесс, протекающий во времени, то узловым вопросом является определение этого времени. Главным путем… Историю Земли подразделяют на несколько очень крупных последовательных… Конец одной эпохи и начало другой сопровождались существенным преобразованием лика Земли, менялось соотношение между…Геохронологическое подразделение истории Земли
| Эра | Период | Эпоха | Продолжи- тельность (в млн. лет) | Время от начала периода до наших дней (в млн. лет) | Преобладающие формы жизни | Климат | Геологические условия |
| Кайнозой (новая) | Четвертичный | Совре- менная | 0,02 | Травянистые растения, млекопитающие. Эпоха человека | теплый | Конец последнего ледникового периода | |
| Поейсто-цен | 1,7 | Вымирание многих видов растений и крупных млекопитающих. Появление человека | Холодный | Повторные оледенения (четыре ледниковых периода) | |||
| Третичный | Плиоцен | Цветковые растения. Млекопитающие (появления лонов, лошадей, верблюдов, сходных с современными) | Прохладный | Поднятие отдельных гор. Остепенение суши. Вулканическая активность | |||
| Миоцен | Появление первых человекообразных обезьян | Прохладный | Образование гор. Вулканическая активность | ||||
| Олигоцен | Леса. Однодольные цветковые растения. Предшественники многих современных родов млекопитающих (включая человекообразных обезьян и людей) | теплый | Материки низменные | ||||
| Эоцен | Пацентарные млекопитающие | теплый | Горы размыты. Внутриконтинентальных морей нет | ||||
| Палеоцен | Примитивные млекопитающие. Угасание царства динозавров | ||||||
| Мезозой | Мел | Покрытосесенные однодольные и двудолные растения. Первые цветки. Динозавры. Примитивные млекопитающие | Образуются горы, отложения мела. Сланцев | ||||
| Юра | Цикадофиты и хвойные. Динозавры. Древнейшие птицы, насекомоядные, сумчатые | Материки довольно возвышены. Мелководные моря | |||||
| Триас | Голосеменные, динозавры | Теплый, усиливается засушливость | Материки приподняты над уровнем моря |
Продолжение табл.32.
| Эра | Период | Эпоха | Продолжи- тельность (в млн. лет) | Время от начала периода до наших дней (в млн. лет) | Преобладающие формы жизни | Климат | Геологические условия |
| Герцинское горообразование | |||||||
| Палеозой (древний) | Пермь | Голосеменные, насекомые, пресмыкающиеся. Появление предков динозавров | Аридный | Материки приподняты, периодическое оледенение | |||
| Карбон (каменноугольный) | Папоротникообразные, плауны, голосеменные, первые настоящие семена. Земноводные | Теплый, влажный | Болота, поднятие суши, материки низменнные | ||||
| Девон | Первые леса (папоротники). Первые настоящие листья. Рыбы. | Аридный | Небольшие внутриконтинентальные моря. Оледенение | ||||
| Силур | Водоросли, хвощи, плауны. Морские паукообразные (скорпионы). Начался выход животных на сушу | Засушливый | Обширные внутриконтинентальные моря. Поднятие суши | ||||
| Ордовик | Морские водоросли. Наутилоидные. Трилобиты, моллюски | Теплый, включая Арктику | Заметное погружение суши | ||||
| Кембрий | Морские водоросли, «взрыв» морских многоклеточныех. Трилобиты и плеченогие, губки и иглокожие | Умеренный | Материки низменные | ||||
| Второе великое горообразование | |||||||
| Протерозой | Примитивные водоросли и водные грибы. Морские простейшие. В конце – моллюски, черви и др. Морские беспозвоночные | Интенсивное осадкообразование. Вулканическая активность, обширная эрозхия и многократные обледенения | |||||
| Первое великое горообразование | |||||||
| Архей | Ископаемые не обнаружены (обнаружено органическое вещество в скальных породах) | Значительная вулканическая активность. Слабое осадкообразование. Обширная эрозия |
Копытные в ходе эволюции разошлись на группу с четным числом пальцев (коровы, овцы, верблюды, олени, жирафы, свиньи и гиппопотамы) и группу с нечетным числом пальцев (лошади, зебры, тапиры, носороги). В эоцене существовали китообразные формы, давшие начало китам и дельфинам. В олигоцене широко были распространены также предковые формы человекообразных обезьян и человека.
В миоцене и плиоцене древесная растительность вытесняется травянистыми формами. Наступает расцвет покрытосеменных растений, членистоногих и млекопитающих. Конец третичного периода датируется появлением всех современных семейств животных и растений.
Следующий период (четвертичный) этой эры, длящийся последние 1,5-1,7 млн лет, состоит из плейстоценовой и современной эпох. Для плейстоцена характерны четыре последовательных оледенения. Растения и животные, которые были распространены в плейстоцене, сходны с современными растениями и животными. Однако в плейстоцене вымерли многие растения (особенно лесные) и млекопитающие (мамонты, саблезубые тигры). Появились многие травянистые формы растений, но, самое главное, появился человек. Современная эпоха началась около 11 000 лет назад, когда отступил последний ледник.
Хотя в палеонтологической летописи существуют многие пробелы, тем не менее она полностью раскрывает происхождение многих существующих в настоящее время видов животных и растений. Например, очень хорошо прослежено происхождение лошадей, слонов, парнокопытных, хищников.
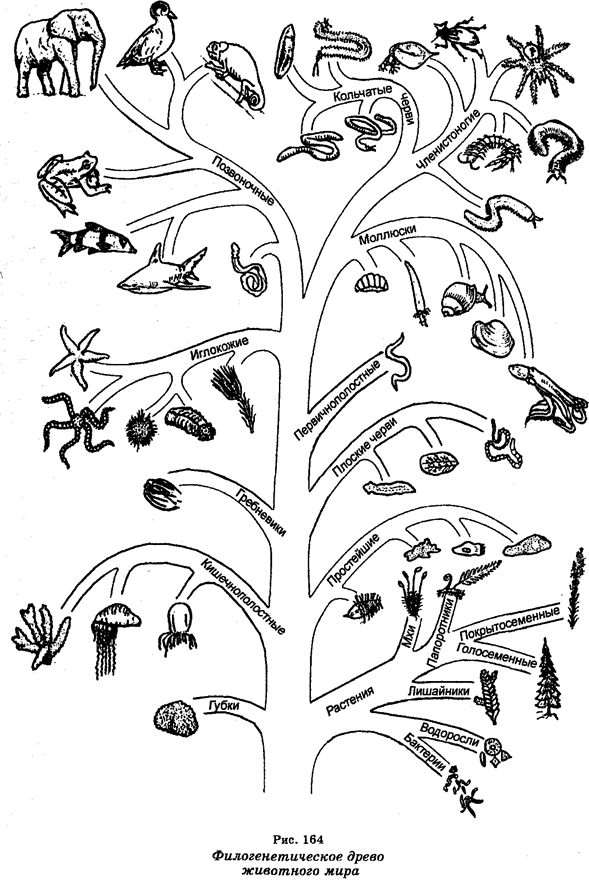
Эволюцию на уровне крупных таксонов называют макроэволюцией. Основополагающие представления о макроэволюции в целом сводятся к тому, что все современные живые существа являются измененными потомками одной или нескольких форм, существовавших ранее. Более молодые по происхождению формы животных и растений произошли от предков, менее отличавшихся друг от друга, а те, в свою очередь, развились от общего единого источника (рис. 164).
Считают, что первыми возникли архебактерии. Примерно 3,5 млрд лет назад. Затем возникли истинные бактерии, а от них ответвились вирусы, цианобактерии, красные водоросли и зеленые жгутиковые. От последних ответвились водоросли (от которых позднее произошли, в свою очередь, высшие растения) и бесцветные ризофлагелляты, из которых развились диатомовые водоросли, плесневые грибы, губки и простейшие. Часть простейших стала паразитической.
Вторым крупнейшим этапом эволюции была трансформация одноклеточных организмов в многоклеточные. Считают, что первые эукариоты появились 1,5 млрд лет назад. Их появление есть крупнейший эволюционный переход, ибо в основе появления эукариотов лежит формирование митоза и развитие клеточных структур. Из цилиарных простейших (цилиофор) развились многоклеточные (синтициальные) плоские черви. Последние разветвились на пять линий, одна из которых повела к иглокожим и хордовым. Остальные линии повели к большинству других типов животного мира.
Первыми позвоночными были остракодермы, которые развились из беспозвоночных, а из остракодерм развились миноги и панцирные акулы. Хрящевые рыбы (акулы и скатовые) и костные рыбы развились из панцирных акул, а из костных рыб развились кроссоптериги, которые являлись примитивными рыбами и которые затем разошлись на две эволюционные ветви, одна из которых привела к дипловертебронам (ранним земноводным), называемым стегоцефалами. Дипловертеброны были предшественниками современных амфибий и пресмыкающихся. Первые пресмыкающиеся (котилозавры) разветвились на ветви, давшие начало динозаврам и морским пресмыкающимся, которые затем вымерли, а также черепахам, крокодилам, ящерицам, змеям, птицам и млекопитающим.
Эволюционная линия, ведущая к человеку, начинается от тупай и ведет к ранним приматам, разветвляясь затем на обезьян Нового света, обезьян Старого света, понгид и гоминид. От понгид (настоящих обезьян) произошли гориллы, орангутанги и шимпанзе, а от гоминид произошел непосредственный предшественник современного человека (см. гл. XVI).
Главные направления эволюции сформулированы А. Н. Север-цовым (1866-1936), который впервые четко определил различия между прогрессивным и регрессивным развитием, обосновал понятия о морфофизиологическом и биологическом прогрессе, а также о морфофизиологическом и биологической регрессе.
Морфофизиологический прогресс — это изменения структуры и общей жизнедеятельности на пути эволюции от простых форм к сложным, от одноклеточных организмов к многоклеточным. Напротив, Морфофизиологический регресс — это упрощение организации и жизнедеятельности, выражающееся в редукции органов (например, хорды у оболочников) на пути эволюции.
Следствием морофофизиологического прогресса является биологический прогресс. Он характерен для организмов, которые характеризуются возрастанием приспособленности к окружающей среде, что ведет к увеличению их численности и расширению их ареалов. Однако биологический прогресс может быть результатом не только морфофизиологического прогресса. Иногда от является и результатом морфофизиологического регресса
В конечном итоге биологический прогресс ведет к процветанию вида.
В современную эпоху процветающими являются покрытосеменные растения, насекомые, костные рыбы, птицы, млекопитающие.
Значение биологического прогресса заключается в том, что он играет важную роль в эволюции и обеспечивает главные направления эволюции в виде ароморфозов, идиоадаптаций и общей дегенерации.
Ароморфозы — это крупные изменения организмов, которые поднимают на новый более высокий уровень их морфофизиологическую организацию и жизнедеятельность. Кроме того, они открывают организмам путь к проникновению в новую среду обитания. Ароморфозы возникают скачками. Примерами ароморфозов А. Н. Северцов называл развитие легких и четырехкамерного сердца, превращение парных плавников рыб в парные конечности земноводных. По А. Н. Се-верцову, ароморфозы обозначили возникновение организмов новых классов и типов организмов, завоевание организмами новых просторов. У млекопитающих ароморфозами были: появление живорождения, шерстного покрова, постоянной температуры тела и др.
Идиоадаптация — это изменения, которые не приводят к повышению уровня организации организмов, но сопровождаются приданием им большей приспособленности к существующим условиям жизни, больших возможностей прогрессивного развития. Примером идиоадаптаций являются приспособления насекомых к жизни в разных средах (в воде, почве, воздухе), к разным типам питания. Другим примером служит разнообразие форм у насекомоядных (землеройка, крот, еж, выхухоль и др.), что позволило занять им разные экологические ниши.
Общая дегенерация — это изменения, которые заключаются в снижении уровня организации живых существ и в снижении активности отдельных или ряда органов. Примерами общей дегенерации А. Н. Северцов называл биологию саккулины из группы паразитических ракообразных (крабов), а также биологию оболочников. Выживаемость этих организмов обеспечивается чрезвычайным разнообразием их личиночных форм.
Следствием морфофизиологического регресса является биологический регресс. Биологический регресс — это снижение приспособленности организмов к среде, сокращение их численности и ареалов. Конечным итогом биологического регресса является вымирание организмов.
Учение А. Н. Северцова о главных направлениях эволюционного процесса было развито дальше в трудах И. И. Шмальгаузена, который рассматривал организм как целое в историческом и индивидуальной развитии. Среди главных направлений эволюционного процесса И. И. Шмальгаузен большое внимание уделял различным формам ароморфозов.
Доказательства эволюции получены в разных науках. Классические доказательства эволюции получены, прежде всего, в палеонтологии в результате изучения ископаемых организмов, живших в прошлые эпохи. Предполагают, что в ходе эволюции вымерло около 200 000 видов животных. В более глубоких слоях Земли обнаруживаются остатки более древних форм жизни, тогда как в поверхностных слоях находят остатки более поздних форм. Можно сказать, что история жизни на Земле написана на языке ископаемых остатков. Палеонтологический материал дает также основания судить о темпах и направлениях эволюции.
Доказательства эволюции получены в биогеографии, которая является наукой о распространении растений и животных. В биогеографии различают шесть биогеографических областей. Каждая из этих областей характеризуется специфическими обитателями (растениями и животными), называемыми эндемиками, под которыми понимают организмы видов, родов и таксонов, ограниченных в своем распространении определенными территориями.
В Палеоарктической области (Европа, африканский север от Са-хары, часть Азии к северу от Гималаев, Азорские острова и острова Зеленого мыса) эндемичными, если говорить о животных, являются кроты, олени, быки, овцы, козы, скворцы и сороки.
В Неоарктической области (Гренландия и Северная Америка) эндемичными являются горные козлы, луговые собачки, опоссумы, скунсы, еноты, сойки и американские грифы. Кроме того здесь встречаются формы, андемичные для палеоарктической области.
В Неотропической области (Южная и центральная Америка, юг Мексики и острова Вест-Индии) обитают альпаки, ламы, цепко-хвостые обезьяны, тапиры, ленивцы, вампиры, муравьеды и многие виды птиц, не встречающиеся в других частях земного Шара.
В Эфиопской области (Африка к югу от Сахары, Мадагаскар) обитают шимпанзе, гориллы, зебры, носороги, трубкозубы, бегемоты, жирафы, многие виды птиц, пресмыкающихся и рыб, не обнаруживаемые в других областях.
В Восточной области (Индия, Цейлон, Индокитай, юг Китая, Малайский полуостров и отдельные острова Малайского архипелага) обитают орангутанги, черные пантеры, индийские слоны, гиббоны и долгопяты. В Австралийской области (Австралия, Новая Зеландия, Новая Гвинея и др. острова Малайского архипелага) эндемичными являются утконосы, кенгуру, вомбаты, коала и другие сумчатые животные. Эндемичными являются бескрылые птицы эму и казуар, а также птица-лира и какаду.
Географические закономерности, характерные для фауны, присущи и флоре этих биогеографических областей.
Одно из основных положений биогеографии заключается в том, что каждый вид растений и животных возникал только однажды и только в одном месте (центре происхождения), откуда он расселялся до тех пор, пока не встречал какую-нибудь преграду, например, географическую, климатическую, пищевую и т. д. Географические ареалы близких видов, как правило, не совпадают, но они и не очень отдалены один от другого.
Особенности географического распространения животных и растений являются отражением специфики эволюции каждого вида. Например, сходство фауны Неоарктической и Неотропической областей обусловлено относительно недавним геологическим разделением Северной Америки и Евразии в результате образования Берингова пролива. Наличие в Австралии однопроходных и сумчатых, которых нет ни в одной из других областей, является результатом того, что этот континент давно (в мезозое) был изолирован от других частей суши. Примитивным млекопитающим — аборигенам Австралии не довелось конкурировать с более приспособленными плацентарными млекопитающими, поэтому они дали начало формам, легко приспособившимся к местным условиям.
Далее, сейчас аллигаторы встречаются только в реках юго-востока США и в р. Янцзы (КНР), тогда как магнолии произрастают только в восточной части США, в КНР и Японии. В начале кайнозоя Северная Америка соединялась с Азией (на месте теперешнего Берингова пролива). При тогдашнем теплом климате аллигаторы и магнолии на этой территории были распространены повсеместно. В дальнейшем из-за поднятия Скалистых гор и похолодания (оледенения) аллигаторы и магнолии погибли повсеместно, кроме юго-восточной части США и восточной части КНР, где не было оледенения. Поскольку виды магнолий и аллигаторов были разделены в отдаленных частях земного шара на протяжении многих миллионов лет и эволюционировали обособленно, то в современную эпоху, оставаясь близкородственными видами, они несколько различаются между собой.
Еще Ч. Дарвин отмечал различия островных и материковых флор и фаун. Например, на Галапагосских островах, возраст которых составляет всего лишь 3—5 млн лет и которые никогда не были связаны с материком, отсутствуют лягушки и жабы, ибо ни они, ни их яйца не смогли бы выжить в морской воде, с которой неизбежно пришлось бы столкнуться мигрирующим на этот остров животным. На этих островах нет также наземных млекопитающих, но на одном из этих островов встречаются гигантские ящерицы и черепахи, которые очень близки некоторым формам, обитающим на побережье Эквадора. Это указывает на эволюцию растений и животных после заселения этих островов.
В систематике главное доказательство эволюции заключается в том, что все живые существа можно расположить в иерархическую систему таксономических единиц — виды, роды, семейства, отряды, классы и типы. Это означает, что все организмы связаны между собой филогенетически в результате существующих между ними эволюционных взаимоотношений. Принадлежность организмов к тем или иным систематическим группам свидетельствует о том, что большинство промежуточных форм, существовавших в прошлом, вымерло. Если бы виды всех существовавших в прошлом организмов жили до настоящего времени, то классифицировать живой мир на таксономические группы было бы невозможно. Будучи генетически различными, виды представляют собой независимо эволюционирующие и репродуктивно изолированные единицы. Поскольку можно предполагать, что у генетически сходных видов общий предок существовал в менее отдаленном прошлом по сравнению с генетически различными видами, то степень генетических различий является, по существу, мерой, на основе которой сейчас совершенствуют филогенетическое древо,
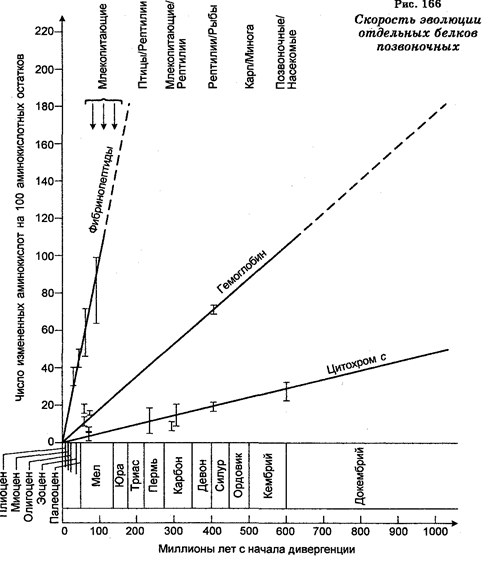
Степень генетических различий между видами определяют либо прямо путем изменений последовательностей нуклеотидов в генах, либо косвенно путем изменений последовательностей нуклеотидов в рРНК, или последовательностей аминокислот в белках. Результаты сравнения последовательностей ДНК разных организмов позволяют определить количество пар нуклеотидов, в которых в ходе эволюции имели место замены азотистых оснований (табл. 33), тогда как сравнение белков от разных организмов позволяет определить различия в аминокислотных последовательностях, т. е. судить о близости организмов (рис. 165) и о связи последовательностей со скоростью эволюции (табл. 34, рис. 166). На основе данных о филогении отдельных белков строят филогенетическое древо, которое, как показано для цитохрома С, совпадает с филогенетическим древом, построенным по ископаемым останкам. На основе реконструкции филогении и определения степени генетических различий по аминокислотным последовательностям ряда белков считают, что гены, кодирующие эти белки у животных, происходят от общего предка.
Таблица 33 - Количества нуклеотидных замен в генах, кодирующих цитохромы С у разных организмов
| Организм | ||||||||||||||||||||
| 1. Человек | — | |||||||||||||||||||
| 2. Макак -резус | — | IS | ||||||||||||||||||
| 3. Собака | — | |||||||||||||||||||
| 4. Лошадь | — | |||||||||||||||||||
| 5. Осел | — | |||||||||||||||||||
| 6. Свинья | — | |||||||||||||||||||
| 7. Кролик | — | |||||||||||||||||||
| 8. Кенгуру | - | |||||||||||||||||||
| 9. Утка | — | |||||||||||||||||||
| 10. Голубь | — | |||||||||||||||||||
| 11. Курица | — | |||||||||||||||||||
| 12. Пингвин | — | |||||||||||||||||||
| 13. Черепаха | — | |||||||||||||||||||
| 14.Гремучая змея | — | |||||||||||||||||||
| 15. Тунец | — | |||||||||||||||||||
| 16.Муха комнатная | — |
С целью определения степени сходства белков используют также иммунологическое и электрофоретическое сравнение белков. Степень иммунологического сходства белков выражают в иммуно-логическом расстоянии (табл. 35), которое приближенно можно связать с различиями по аминокислотным последовательностям. Электрофоретические исследования позволяют выявить электрофоретическое сходство белков и на основе этих данных также определить генетические расстояния между видами.

Таблица 34 - Связь между аминокислотными различиями в и-и р-глобинах со скоростью эволюционной дивергенции
| Животные | a-цепь | b-цепь | a+b (среднее) | Миллионы лет с начала дивергенции |
| Плацентарные в сравнении между собой | 16,1 | 16,7 | 16,4 | |
| Кенгуру в сравнении с плацентарными млекопитающими | 21,7 | 26,9 | 24,3 | |
| Куры в сравнении с млекопитающими (сумчатыми и плацентарными) | 29,6 | 31,7 | 30,6 | |
| Змеи в сравнении с теплокровными | 39,2 | |||
| Земноводные в сравнении с наземными животными | 46,7 | 48,9 | 47,8 | |
| Костные рыбы в сравнении с четвероногими (рептилиями, амфибиями и млекопитающими) | 49,3 | 49,6 | 49,5 |
Таблица 35 - Иммунологические различия между приматами (по альбуминам)
| Вид приматов | Антисыворотка в альбуминам | ||
| Человека | Шимпанзе | Гиббона | |
| Человек | О | 3,7 | 14,4 |
| Шимпанзе | 5,7 | 14,6 | |
| Горилла | 3,7 | 6,8 | 11,7 |
| Орангутан | 8,6 | 9,3 | 11,7 |
| Гиббон | 10,7 | 9,7 |
Эволюция подтверждается данными сравнительной морфологии, эмбриологии, физиологии, биохимии и генетики.
Результаты сравнительного изучения строения животных в пределах систематических групп показывают, что оно имеет общий план (рис. 167). Особенно это проявляется в случае гомологичных органов, которые характеризуются общим строением, сходством эмбрионального развития, иннервации, кровоснабжения и отношения к другим органам. Например, гомологичными являются передний ласт тюленя, крыло летучей мыши, передняя нога собаки и рука человека (рис. 168) и другие. Эти органы состоят из почти одинакового количества костей и мышц, их сосуды и нервы имеют сходную топографию. Гомологичные органы с несомненностью указывают на общность происхождения животных, у которых есть эти органы.
Помимо гомологичных органов морфологическим свидетельством в пользу эволюции являются данные о рудиментарных органах, являющихся «остатками» развитых органов, имевшихся в прошлом у предковой формы. Например, у человека имеется свыше 100 разных рудиментов, типичными примерами которых являются червеобразный отросток слепой кишки (аппендикс), копчик (слившиеся хвостовые позвонки), волосяной покров туловища и конечностей, третье веко. Червеобразный отросток слепой кишки является результатом перехода человека на диету с большим содержанием мяса и меньшим количеством клетчатки. Слепой вырост в таких условиях уменьшился и стал рудиментом. Напротив, у травоядных животных он представляет собой активно функционирующий орган. Кости задних конечностей китов в связи с переходом их к водному образу жизни стали рудиментами, располагающимися в толще брюшных мышц.
На эмбриологические доказательства эволюции обращал внимание еще Ч. Дарвин. В 1866 г. Э. Геккель сформулировал биогенетический закон, в соответствии с которым онтогенез есть повторение филогенеза (краткое и быстрое резюме филогенеза по терминологии Э. Геккеля), т. е. зародыши в процессе развития как бы сокращенно повторяют эволюционный путь своих предков. Как оказалось позднее, Э. Геккель был прав в принципе, но не в деталях, поскольку зародыши высших животных сходны лишь с зародышами низших животных, но не со взрослыми особями низших форм, как это считал Э. Геккель.
Эмбриологические данные в пользу эволюции сводятся к тому, что на ранних стадиях развития между зародышами млекопитающих, рыб, земноводных и пресмыкающихся существует много сходных признаков. В то же время эти организмы обладают рядом собственных признаков, присущих только им и обеспечивающих приспособленность к развитию в утробе материнского организма, но не под скорлупой яйца.
 Данные эмбриологии свидетельствуют о том, что после гаструляции развитие зародышей проходит в одном из двух направлений. С одной стороны, бластопор превращается либо в анальное отверстие, либо занимает положение около этого отверстия, как это имеет место в случае иглокожих и хордовых. С другой стороны, бластопор развивается в ротовое отверстие, либо занимает положение около этого отверстия, как это имеет место в случае кольчатых червей, моллюсков, членистоногих и др. При развитии как в одном направлении, так и другом между эктодермой и энтодермой развивается мезодерма, которая у иглокожих и позвоночных закладывается частично (карманами или выпячиваниями первичной кишки), а у кольчатых червей и других организмов закладывается из специальных клеток, дифференцирующихся на ранних этапах развития.
Данные эмбриологии свидетельствуют о том, что после гаструляции развитие зародышей проходит в одном из двух направлений. С одной стороны, бластопор превращается либо в анальное отверстие, либо занимает положение около этого отверстия, как это имеет место в случае иглокожих и хордовых. С другой стороны, бластопор развивается в ротовое отверстие, либо занимает положение около этого отверстия, как это имеет место в случае кольчатых червей, моллюсков, членистоногих и др. При развитии как в одном направлении, так и другом между эктодермой и энтодермой развивается мезодерма, которая у иглокожих и позвоночных закладывается частично (карманами или выпячиваниями первичной кишки), а у кольчатых червей и других организмов закладывается из специальных клеток, дифференцирующихся на ранних этапах развития.
Вслед за появлением мезодермы у всех хордовых развивается дорзальный полый нервный тяж, а также нотохорд (основа скелета тела) и жаберные щели (отверстия в глотке). На ранних стадиях развития у зародышей, например человека, как и у зародышей рыб, имеются жаберные щели, дуги аорты, пересекающие жаберные перегородки, сердце с одним предсердием и одним желудочком, примитивная почка (пронефрос) и хвост с мышцами. Таким образом, на ранних стадиях развития зародыши человека напоминают зародыши рыб.
На более поздних стадиях развития зародыши человека приобретают сходство с зародышами пресмыкающихся, в частности, зарастают жаберные щели, сливаются кости, из которых состоят позвонки, исчезает старая почка (пронефрос) и образуется новая почка (мезонефрос), разделяется предсердие на правую и левую части. Еще позднее у зародышей человека развивается четырехкамерное сердце и метанефрос (совершенно новая почка), исчезает нотохорд. На седьмом месяце внутриутробного развития плод человека покрыт волосами и имеет «обезьянье соотношение» в размерах тела и конечностей.
Явление рекапитуляции признаков у человека объясняют действием сохранившихся «рыбьих», «земноводных» и «пресмыкающихся» генов, существовавших у предковых форм, вслед за которыми действуют гены млекопитающих, затем приматов. Следовательно, на начальных стадиях развития человека действуют гены далеких предков и проявляются признаки, сходные с признаками далеких предков, на поздних же стадиях действуют гены, характерные для млекопитающих, в частности приматов, в результате чего проявляются признаки млекопитающих и приматов. Закономерность здесь такова, что у зародышей человека раньше проявляются признаки, присущие типам и классам, а позднее признаки, присущие родам и видам.
Например, у человека и свиньи процессы развития дивергиру-ют раньше, чем у человека и человекообразных обезьян, поскольку предки человека разошлись с предками свиней раньше, чем произошло расхождение человека и человекообразных обезьян, эволюционировавших от одного предка.
Основные физиологические доказательства эволюции заключаются в сходстве физиологических процессов — дыхания, пищеварения, кровообращения, выделения, реакций на раздражения у организмов многих систематических групп.
Данные биохимии в пользу эволюции очень значительны. Во-первых, химический состав всех живых организмов является сходным. В элементарном составе организмов преобладают три элемента — С, О и Н, в атомарном составе 50-60% приходится на С (в земной коре он составляет 0,35%), что указывает на естественное родство всего органического мира. Далее, у всех организмов белки построены из 20 аминокислот. Между белками животных и растений имеется сходство. Например, между гемоглобином позвоночных, гемоцианином беспозвоночных и хлорофиллом растений существует принципиальное сходство, что указывает на единство происхождения всех организмов. Кроме того, имеется иммуноло-гическое сходство между белками крови разных животных. Например, ближайшими родственниками человека являются человекообразные обезьяны, затем обезьяны Старого света, цепкохвостые обезьяны Нового света и, наконец, долгопяты. Кошки, собаки и медведи образуют одну родственную группу, тогда как коровы, овцы, козы олени и антилопы — другую. Тюлени и морские львы стоят ближе к хищникам.
Отмечается сходство ферментов и биосинтетических путей у различных организмов. У всех животных и растений в качестве универсального звена в энергетических реакциях используется АТФ. Налицо также сходство продуктов выделения. У человека и других приматов конечным продуктом обмена пуринов является мочевая кислота, у других млекопитающих — аллантоин, у земноводных и у большинства беспозвоночных — мочевина, а у некоторых беспозвоночных — аммиак. Эволюция позвоночных сопровождалась постепенной потерей способности синтезировать ферменты, необходимые для постепенного распада мочевой кислоты.
Основное доказательство эволюции, представляемое генетикой, заключается в установлении универсального характера ядерного генетического кода. Универсальностью обладают также митохон-дриальный и хлоропластный генетические коды. Можно сказать, что ядерный и экстраядерный генетические коды — это важнейшее доказательство дарвиновской идеи единства происхождения всех форм жизни, т. е. происхождения всех живых форм из одного корня.
Наконец, в пользу эволюции свидетельствует практика сельского хозяйства, на что неоднократно указывал сам Ч. Дарвин, приводя множественные примеры происхождения культурных растений и домашних животных от диких предков. Например, все сорта белокочанной, листовой, цветной и брюссельской капусты, а также кольраби и брокколи происходят от одного вида дикой капусты, а кукуруза берет начало от травянистого растения (теосинта), произрастающего в Андах и в Мексике. Можно сказать, что среди культивируемых сельскохозяйственных растений нет ни одного вида, который был бы в «диком» состоянии. Это же заключение относится и к домашним животным. Все существующие ныне породы крупных и мелких домашних животных также происходят от диких предков.
Учение о микроэволюции и видообразование
Эволюция на уровне вида (на внутривидовом уровне) получила название микроэволюции. В рамках учения о микроэволюции различают элементарную… Элементарной единицей эволюции является популяция. Популяция (от фр.… Для популяций характерны такие показатели, как ареал, численность особей, размеры по числу особей в пространстве,…Генотипическая изменчивость в природных популяциях
Животных и растении
Как отмечено в начале этого раздела, эволюция протекает на всех уровнях…Гипотеза нейтральности молекулярной эволюции
Обобщение результатов молекулярно-генетических исследований эволюции привело японского ученого М. Кимуру (1968) и других исследователей к… При одинаковой способности альтернативных аллелей придавать организмам… По гипотезе нейтральности молекулярной эволюции большинство мутаций любых генов вредно для организмов и мутанты…Антидарвиновские концепции эволюции
С самого начала теория эволюции оказалась ареной острой борьбы идей, в которой дарвинизму с самого начала стали противопоставлять неоламаркизм или… В неоламаркизме различают психоламаркизм и механоламар-кизм. Психоламаркисты в… В XIX в. возник и перешел в XX в. социал-дарвинизм, основателем которого был Э. Геккель, много сделавший для защиты и…Вопросы для обсуждения
1. В чем заключается эволюционизм? 2. На чем основываются современные доводы в пользу эволюции? 3. Почему современную теорию эволюции называют синтетической и в чем состоит ее отличие от классического дарвинизма? …Литература
Аиала Ф. Введение в популяционную и эволюционную генетику. М.: Мир. 1984. 227 стр.
Дубинин Н. П. Генетика. Кишинев: Штиинца. 1985. 533 стр.
Майр Э. Популяции, виды и эволюция. М.: Мир. 1974. 460 стр.
Опарин А. И. Жизнь, ее природа, происхождение и развитие. М.: Наука. 1968.
Руттен М. Происхождение жизни. М.: Мир. 1973. 411 стр.
Северцов А. Н. Главные направления эволюционного процесса. М.: Виомедгиз. 1934. 150 стр.
Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. В. Краткий очерк теории эволюции. М.: Наука. 1969. 407 стр.
Яблоков А. В.. Юсуфов А. Г. Эволюционное учение. М.: Высшая школа. 1989. 335 стр.
Czifto G. Without Miracles Universal Selection Theory and the Second Darwinian Revolution. NIT Press. 1957. 385 pp.
Rosemweig M. L. Species Diversity in Space and Time. Cambridge University Press. 1995. 436 pp.
– Конец работы –
Используемые теги: Теория, эволюции0.048
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: ТЕОРИЯ ЭВОЛЮЦИИ
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.139 сек.
Новости и инфо для студентов