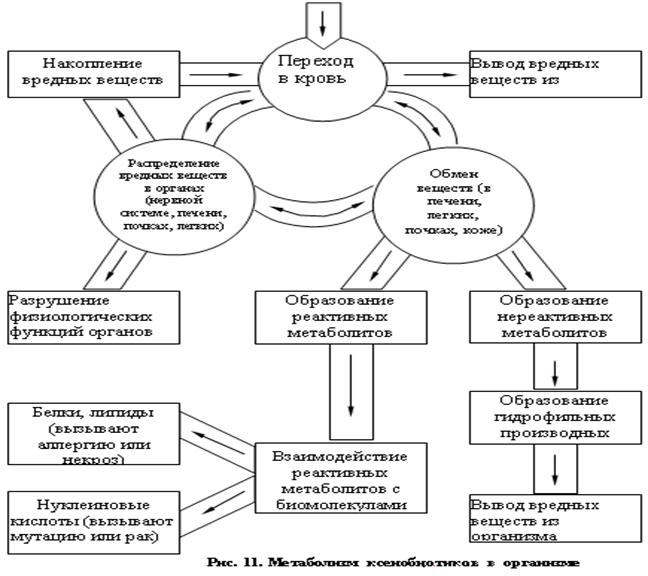
Превращение токсичных веществ в организме
Проникающие в организм ксенобиотики могут подвергаться различным биохимическим превращениям – биотрансформации. При этом может происходить образование нереактивных и реактивных метаболитов. Нереактивные метаболиты превращаются в гидрофильные производные, которые выводятся из организма. Реактивные метаболиты способны вступать в реакции с биомолекулами, например с белками или липидами, и быть причиной аллергии или некроза, взаимодействовать с нуклеиновыми кислотами, вызывая мутации или развитие злокачественных опухолей, и т.п. Схема метаболизма вредных веществ в организме приведена на рис. 11 [16].
В основе биотрансформации токсичных веществ с образованием нереактивных метаболитов лежит несколько типов химических реакций, в результате которых происходит присоединение или отщепление метильных (-CH3), ацетильных (CH3CO-), карбоксильных (-COOH), гидроксильных (-OH) радикалов (групп). В основном метаболизм ксенобиотиков идет по пути ферментативного окисления (часто сопровождающегося гидролитическим расщеплением) и восстановления, неферментативного окисления, восстановления и гидролиза, реакций синтеза и конъюгации.
Окислению микросомальными энзимами подвергаются разнообразные по строению органические липоидорастворимые соединения. В основе этих реакций обычно лежит гидроксилирование. Реакции микросомального гидроксилирования протекают по следующим схемам [22]:
гидроксилирование ароматического кольца
C6H5R ® HOC6H4R
гидроксилирование боковой цепи (ациклическое)
RCH3 ® RCH2OH
 N-дезалкилирование
N-дезалкилирование
R-NH-CH3 ® [RNHCH2OH] ® RNH2 + HC
нестойкое

O-дезалкилирование
R-O-CH3 ® [ROCH2OH] ® R-OH + HC
нестойкое
дезаминирование
R-CH(NH2)CH3 ® [RCOH(NH2)CH3] ® RCOCH3 + NH3
нестойкое
образование сульфоксида
R-S-CH3 ® [R-S-CH2OH] ® R-SOCH3
Микросомальному восстановлению подвергаются ароматические нитро- и азосоединения и алифатические галогенсодержащие соединения. Предполагаются следующие этапы восстановления, включающие и неферментативную фазу: микросомальный ферментативный комплекс НАДФ×Н2–цитохром–с–редуктаза или НАД×Н2 (никотинамидадениннуклеотид)–цитохром–в–редуктаза восстанавливает ФАД (флавинадениннуклеотид) в ФАД×Н2. Последний неферментативно восстанавливает ксенобиотик
3 ФАД×Н2 + RNO2 ® 3 ФАД + RNH2 + 2 H2O
 Немикросомальные реакции окисления, восстановления и гидролиза. Существуют многие ферментные системы, катализирующие превращения как эндогенных, так и экзогенных субстратов. Например, в растворимой фракции гомогенатов печени, почек и легких содержится алкогольдегидрогеназа, которая быстро окисляет многие первичные спирты в соответствующие альдегиды (медленно этиленгликоль, но не диэтиленгликоль). Необходимым коферментом этих реакций является НАД или НАДФ и участие цитохрома Р-450:
Немикросомальные реакции окисления, восстановления и гидролиза. Существуют многие ферментные системы, катализирующие превращения как эндогенных, так и экзогенных субстратов. Например, в растворимой фракции гомогенатов печени, почек и легких содержится алкогольдегидрогеназа, которая быстро окисляет многие первичные спирты в соответствующие альдегиды (медленно этиленгликоль, но не диэтиленгликоль). Необходимым коферментом этих реакций является НАД или НАДФ и участие цитохрома Р-450:
 |
CH3CH2OH + НАД CH3C + НАД×Н2
Окисление многих алифатических и ароматических альдегидов в соответствующие карбоновые кислоты выполняют такие ферменты, как альдегидоксидазы и ксантиноксидазы:
|

 C6H5-C + H2O + НАД C6H5COOH + НАД×Н2
C6H5-C + H2O + НАД C6H5COOH + НАД×Н2
Гидролитическому расщеплению подвергаются сложные эфиры и амиды кислот. В этом процессе участвуют ферменты (эстеразы, амидазы), находящиеся в печени и в плазме крови:
RCOOR’ + H2O 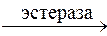 RCOOH + R’OH
RCOOH + R’OH
Сложный эфир Кислота Спирт
RCONH2 + H2O 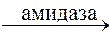 RCOOH + NH3
RCOOH + NH3
Амид кислоты Кислота
Биотрансформация галогенсодержащих соединений может происходить также путем гидролитического дегалогенизирования в печени и почках с образованием свободных хлор- и бром-ионов и соответствующих продуктов гидролиза:
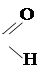
CH2ClBr 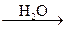 HC + Cl– + Br –
HC + Cl– + Br –
Реакции синтеза и конъюгации. В результате первичных реакций биотрансформации ксенобиотики могут приобретать химически активные группы (-OH, -COOH, -NH2, -SH и др.), способствующие дальнейшей реакции конъюгации с легкодоступными эндогенными субстратами: глюкуроновой кислотой, сульфатом, уксусной кислотой, некоторыми аминокислотами. Конъюгирование приводит к образованию более полярной молекулы, легко выделяющейся из организма почками.
Образование конъюгатов – сложный биохимический процесс, в основе которого лежит активирование эндогенного субстрата при участии специфических в каждом случае ферментов.
Глюкуроновая конъюгация. Конъюгация с глюкуроновой кислотой является наиболее универсальной реакцией, характерной для связывания различных токсичных соединений практически у всех видов млекопитающих (за малым исключением).
Источником глюкуроновой кислоты является глюкоза или исходные для ее получения сахара. Глюкоза активируется при участии АТФ (аденозинтрифосфата). Специфическим коферментом при образовании глюкуроновой кислоты служит уридиндифосфат (УДФ).
Сульфатная конъюгация является общей реакцией для большинства млекопитающих. С сульфатами реагируют фенолы, первичные алифатические спирты, аминосоединения. Конъюгат имеет структуру сложного эфира. Первой фазой этих реакций является активация сульфата, протекающая с закрытой энергией при участии АТФ и ряда ферментов. Образующийся 3-фосфоаденозин-5-фосфосульфат (ФАФС) реагирует непосредственно с ксенобиотиками. Катализируют эту реакцию ферменты сульфотрансферазы (сульфокиназы), отличающиеся субстратной специфичностью.
Метилирование. Основным источником метильных групп служит метионин, превращающийся при участии АТФ в кофермент S-аденозилметионин. Последний под влиянием фермента метилтрансферазы отдает метильные группы соответствующему ксенобиотику. Эта реакция имеет ограниченное значение в метаболизме ядов. Она более характерна для процессов обмена веществ.
Ацетилирование. Реакции ацетилирования ксенобиотиков возможны при наличии в них аминогруппы. В токсикологии ацетилированию подвергаются нефизиологические аминокислоты и в основном ароматические амины.
Непосредственным источником ацетильных групп в организме служит ацетил KoA (KoA-S-COCH3).
Синтез меркаптуровых кислот. Меркаптуровые кислоты образуются в организме при биотрансформации некоторых ароматических углеводородов и галоген- или нитропроизводных алифатических и ароматических углеводородов. Меркаптуровые кислоты являются S-арил- или S-алкил-N-ацетилцистеинами общей формулы
R-S-CH-CH2-COOH
½
NH-COCH3
Образование их многоступенчато. Предполагается реакция ароматического соединения или его эпоксида с глютатионом, превращение этого конъюгата в l-цистеиновое производное и ацетилирование последнего. В результате этой цепи ферментативных реакций образуются меркаптуровые кислоты, выделяющиеся почками.
Непосредственное образование в организме самой меркаптуровой кислоты имеет место при замещении лабильного атома галоида или нитрогруппы углеводорода. В этих случаях l-ацетилцистеиновая часть замещает лабильную группу в молекуле ксенобиотика.
Возможно образование и других типов меркаптуровых кислот при биотрансформации некоторых серосодержащих ароматических соединений, а также бромалканов.
Металлы и их соединения, в отличие от многих органических соединений, попадая в организм, многократно могут менять свою форму. В результате взаимодействия с окислительно-восстановительными буферными системами клетки, при котором осуществляется перенос электронов, степень окисления металлов меняется. При этом переход в состояние низшей степени окисления для большинства переходных металлов обычно связывается с уменьшением их токсичности.
Соединения одного и того же металла, но разной валентности проявляют неодинаковую степень токсичности вследствие разной способности образовывать малорастворимые соединения с биокомплексами и неодинакового сродства к белкам и ферментам. Зависимость токсического действия от степени окисления металла весьма четко проявляется при поступлении оксидов металлов в организм в виде пыли ингаляционным путем. Оксиды большинства переходных металлов (например, оксиды марганца, молибдена, ванадия и др.) в высшей степени окисления обладают более выраженным раздражающим действием на слизистую оболочку органов дыхания и на легочную ткань, вызывая бронхопневмонию.