
Разумный мозг. Кортикальная организация и селекция групп в теории высших функций головного мозга
Разумный мозг. Кортикальная организация и селекция групп в теории высших функций головного мозга - Лекция, раздел Медицина, Текст Взят С Психологического Сай...
Текст взят с психологического сайта http://www.myword.ru
На данный момент в библиотеке MyWord.ru опубликовано более 2000 книг по психологии. Библиотека постоянно пополняется. Учитесь учиться. Удачи! Да и пребудет с Вами.... :)Разумный
МОЗГ
КОРТИКАЛЬНАЯ ОРГАНИЗАЦИЯ
И СЕЛЕКЦИЯ ГРУПП В ТЕОРИИ ВЫСШИХ ФУНКЦИЙ
ГОЛОВНОГО МОЗГА
Перевод с английского
канд. биол, наук, Н. Ю. Алексееико
под редакцией
д-ра биол. наук
Е. Н. Соколова
Издательство
«Мир»
Москва
ББК 28.903 Э19
УДК 612+577.3
Эделмен Дж., Маунткасл В.
Книга посвящена одной из основных проблем современной нейрофизиологии — нейронной организации активности головного мозга Широко известные… Предназначена для нейрофизиологов, биофизиков, специалистов по бионике и…Э50300—123 123—81, ч. 1 2007020000 ББК 28.903
Редакция литературы по биологии © 1978 by The Massachusetts Institute of Toilinology © Перевод на русский язык, «Мпр», 1981И
1975), возможное значение которых послужило темой одной из работ (Schmitt, Dev, Smith, 1976) и большинства выступлений на Конференции 1977 г.
В разных отделах коры форма колонок различна в зависимости от числа и характера упаковки составляющих их мини-колонок. Именно колонки описываются как основные локальные элементы. Про новую кору сказано, что вся она гораздо однороднее, чем думали раньше; стремительное увеличение ее в процессе филогенеза произошло путем умножения колонок, одинаковых в своей основе, а не путем развития новых типов нейронов или разных способов внутренней организации. Связи внутри колонок и между ними организованы очень точно, но они составляют распределенные системы, которые v служат распределительным функциям. Внутренняя структура коры везде одинакова и состоит из повторяющихся элементов.
Новая кора, с афферентными входами внутрикорко-вых колонок, имеет выходы почти ко всем главным образованиям ЦНС, причем значительную их часть составляют массивные возвратные системы с многими входными и выходными точками.
Неискушенного читателя могут удивить два момента в статье Маунткасла: во-первых, подсчеты Пауэлла и сотр. (Rockel, Hiorns, Powell, 1974) говорят о видимой точности, с какой могут быть определены размеры и число нейронов, содержащихся в тонком кортикальном цилиндре (около 110 нейронов у всех исследованных млекопитающих, за исключением 260 в стриарной коре приматов); во-вторых, 600 млн. мини-колонок Коры человека состоят приблизительно из 50 млрд. нервных клеток. Всего лишь около десяти лет прошло с тех пор, как почти во всех учебниках было написано, что общее число нейронов во всем головном мозгу составляет 10 млрд.
Об этом огромном числе Маунткасл говорит в своей статье не мимоходом; оно играет важную роль и в рассуждениях Эделмена, который указывает па тот факт, что число нейронных групп — величина того же порядка, что и число возможных вариантов макромолекул гамма-глобулина, участвующих в явлениях иммунитета. Это согласуется с его представлением, по которому субстраты высшей
Ф. Шмитт
функции мозга, как и иммунологические процессы, взаимодействуют по принципу отбора (селекции).
Помимо поразительного множества нейронов, участвующих в деятельности мозга, и повторного характера их организации в виде локальпых групп (колонок и составляющих их мини-колопок) Маунткасл особенно подчеркивает еще один факт, который играет ваншую роль также и в концепции Эделмена, а именно что каждая из основных элементарных подгрупп, составляющих новую кору, соединяется системой связей с такими же подгруппами колонок в других участках головного мозга, образуя распределенные системы и системы повторного входа (reentrant). Это хороню согласуется с гипотезой Эделмена, по которой фазная цикличность внутренней активации, генерируемой при такой организации, может облегчить постоянное обновление воспринимаемого образа собственного «я».
Самой удивительной чертой теории Эделмена является ее жесткий и последовательный селекционизм; поступающие сенсорные сшналы не играют роли в образовании основных анатомических связей, составляющих первичный ассортимент (набор). Селекциопные операциональные элементы представляют собой группы (каждая из них состоит из сотен или тысяч нейронов), которые составляют локальные единицы («первичпый ассортимент») и в больших количествах входят в состав коры. Колонки и мини-колонки Маунткасла прекрасно подходят под это определение. Локальные группы нейронов образуют первичный ассортимент, аналогичный, но не гомологичный пуклеотидным кодонам генетического кода. Внутренние и внешние нейронные сети в единичных группах и между ними определены генетически и онтогенетически. Набор (ассортимент) нейронных групп — колонок, или модулей — делает возможным вырожденное соответствие групп ассортимента сенсорным сигналам; это значит, что ассортимент способен распознавать данные входные сигналы несколькими способами. Понятие вырожденности отлично от понятия избыточности, которое характеризует идентичные структуры.
«Опознание» единичных групп осуществляется электрическим, ультраструктурным путем и/или по связям И
Введение
отличается точностью. Множественная сигнализация группам первичного ассортимента ведет к ассоциативному распознаванию и образованию вторичного ассортимента нейронных групп с большей вероятностью ответа, чем группы клеток первичного ассортимента.
В результате опыта и образования вторичных ассортиментов формируются структуры, которые различают собственные входные сигналы и внешние.
Согласно развиваемой гипотезе, сознание может возникать на основе повторновходной сигнализации, при которой возникают ассоциации между текущей сенсорной входной активностью и заложенными в память комбинациями (pattern) нейронных групп. В деталях определены те этапы, по которым повторновходные сигналы обрабатываются относительно реакций в первичных и вторичных ассортиментах.
Важно подчеркнуть, что концепция повторного входа является критической. Ввиду вырожденной природы постулируемого селекционного процесса отсутствие повторного входа привело бы к отсутствию непрерывности в системе и к невозможности образования координированных абстрактных отображений внешних сигналов. Иными словами, повторный вход гарантирует непрерывность в распределенной селективной системе. Сознание может быть своего рода ассоциативным обновлением памяти повторным входом, которое непрерывно подтверждает или изменяет теорию личности параллельными сенсорными или моторными входами и выходами.
До какой степени работы Маунткасла и Эделмена дополняют друг друга, было ясно и тогда, когда они были доложены, но это стало особенно очевидно из полного текста их рукописи. Эти статьи, разумеется, войдут в большой том «The Neurosciences. Fourth Study Program» («Нейробиология, Четвертая исследовательская программа»), в котором будут собраны труды Конференции 1977 г. Но для того, чтобы подчеркнуть то общее, что есть в этих работах, и то, как содержащиеся в них сведения дополняют друг друга, а также сделать эти труды более доступными широкому и разнообразному кругу читателей, мы сочли нужным опубликовать их в единой книге, которая и предлагается вниманию читателя.
Ф. Шмитт
ЛИТЕРАТУРА
Rakic P., 1975. Local Circuit Neurons (Neurosci. Res. Program Bull.
13, No. 2), Cambridge Mass., The MIT Press. Rockel A. 1, Hiorns R. W., Powell T. P. S., 1974. Numbers of neurons
through full depth of neocortex, Proc. Anat. Soc. Gr. Br. Ire., 118,
371. Schmitt F. 0., Dev P., Smith В. Н., 1976. Electronic processing of
information by brain cells, Science, 193, 114—120. Schmitt F. 0., Worden F. G., eds., 1978. The Neurosciences: Fourth
Study Program, Cambridge, Mass., The MIT Press.
ОРГАНИЗУЮЩИЙ ПРИНЦИП ФУНКЦИИ МОЗГА -
ЭЛЕМЕНТАРНЫЙ МОДУЛЬ
И РАСПРЕДЕЛЕННАЯ СИСТЕМА
Введение Вряд ли можно сомневаться в господствующем влиянии дарвиновской революции… Достижения последних десятилетий требуют новых формулировок, которые включали бы иерархический принцип организации…Филогенез неокортекса
Предполагается, что современные примитивные насекомоядные очень мало изменились по сравпению со своими предками, от которых произошла также линия… Организующий принцип функции мозга использовании такого аллометрического метода встречаются некоторые необычные отношения; так, высокий эволюционный ранг…Онтогенез новой коры
Организующий принцип функции мозга рано, главным образом из вентрикулярной зоны, могут передвигаться по своим… На осповании результатов этих исследований, развития новой коры и изучения ряда препаратов мозга, взятых у плодов…Существует ли причинная связь
Между цитоархитектоническими и функциональными
Различиями разных областей коры?
Теперь эти старые споры, по-видимому, улажены. Все согласились на том, что различия между основными областями новой коры действительно существуют и… Организующий принцип функции мозга могут быть описаны объективно. Они представляют собой постоянное свойство новой коры любого вида животных, и между…Колончатая организация коры больших полушарий
1. Кортикальная колонка представляет собой обрабатывающее устройство с входом и выходом. Число других участков, передающих сигналы в традиционно… 2. Расположение в виде колонок делает возможным Организующий принцип функции мозгаЗе
В. Маунткасл
Эта общая мысль получила дальнейшее подтверждение в работе Цеки, сочетающей электрофизиологические и анатомические исследования. Цеки обследовал ряд проекций 17-го поля в полях 18-м и 19-м, а также в коре заднего края верхпей височной борозды у обезьяны. В каждой из этих зон он наблюдал дальнейшую, более тонкую обработку какого-либо свойства стимула (Zeki, 1974, 1975, 1977).
Итак, в стриарной зрительной коре проявляются некоторые общие свойства колончатой организации.
1. В ее двумерной матрице картировано несколько переменных.
2. Ее макро-колопки функционируют как такие устройства, ведущие обработку между входом и выходом, при которых параллельная обработка в каждом из них делает возможной селекцию некоторых динамических свойств стимула для формирования в ограниченных каналах, ведущих к определенным выходам (обычно эти свойства усиливаются в перерабатывающих механизмах, находящихся в областях, куда направлены эти каналы).
3. Группы ее колонок избирательно связаны с упорядоченными группами колонок в других областях коры и в модулях подкорковых структур, па которые опа проецируется.
4. Зрительная кора обезьяны подчиняется своей гени-кулостриарной проекции, и этим она сходна с представительствами кисти и стопы в соматосенсорной коре по степени изоляции входа.
5. Колопчатая организация зрительной коры совместима с принципом частично сдвинутого перекрывания в представительстве полей зрения в пространстве нервной системы.
Данные исследования слуховой коры
Давно известно, что участки улитки, и тем самым частота звуковых стимулов, представлены по порядку в «первичной» слуховой коре А1; эти сведения получены главным образом во множестве опытов с вызванными потенциалами, проведенными Вулси и его сотрудниками (см. обзор Woolsey, 1960). Такие же методы были применены для идентификации слуховой коры в юловном моз-
Организующий принцип функции мозга
гу человека на верхней поверхности височной доли в области, соответствующей поперечным височным извилинам (Celesia, 1976). Деление на колонки выражено в кониокортексе этой области более отчетливо, чем в каком-либо другом участке гетеротипической коры; радиальное расположение клеток ясно видно здесь на срезах, перпендикулярных к поверхности коры. Так, Соуса-Пинта (Sou-sa-Pinta, 1973) описал в поле А1 у кошки вертикальные цилиндры клеток в средних слоях диаметром 50—60 мкм с центрами, содержащими лишь небольшое число нейронов. Исследование физиологических свойств отдельных нейронов А1 показало, что клетки, с которыми встречается микроэлектрод, проходят вниз по такой вертикальной колонке, настроены приблизительно на одну и ту же частоту и кривые их настройки резко снижаются с обеих сторон, что, по крайней мере отчасти, объясняется латеральным торможением (Parker, 1965; Abeles, Goldstein, 1970; Merzenich, Brugge, 1973; Merzenich, Knight, Roth, 1975; Imig, Adrian, 1977). Такая же колончатая организация в отношепии предпочитаемой частоты обнаружена в передней слуховой области (Knight, 1977). Перекрывание кривых настройки нейронов в смежных равночастотных полосках хорошо объясняет частично сдвинутое перекрывание частотного представительства, наблюдаемое в опытах по картированию поверхности.
Тунтури первый высказал предположение, что другие свойства слуховых сигналов, возможно, картируются вдоль каждого равночастотного контура, или полоски, коры под прямым углом к направлению изменения частоты (Tuntu-ri, 1952; Tunturi, Dudman, 1958). Теперь доказано, что это действительно так у кошки (Imig, Adrian, 1977) и у обезьяны (Brugge, Merzenich, 1973). У кошки в высокочастотном участке области А1 свойства доминантности уха и бинаурального взаимодействия меняются вдоль по направлению равночастотной полосы. Нейроны, лежащие в такой полосе по перпендикулярной оси коры, обладают одинаковой частотной чувствительностью и одинаковым бинауральным взаимодействием. Микроэлектрод, наклонно вводимый в равночастотную полосу, часто проходит из зоны с одним видом бинаурального взаимодействия нейронов в зону с другим его видом.
В. Маунткасл
Бинауральная стимуляция может вызвать подавление ответа по сравнению с ответом па стимуляцию одного уха, и во всех таких случаях доминирует контралатераль-ное ухо. Вместе с тем бинауральная стимуляция может приводить к суммации, и в этом случае доминировать может любое ухо. Колонки подавления и суммации ответов расположены непрерывными полосами, идущими более или менее под прямым углом к равночастотным коп-турам. В каждой суммационной группе в свою очередь происходит деление на колонки, которые характеризуются параметром доминантности одного уха. Хорошо известно, что интерауральная разница в интенсивности служит параметром, по которому локализуются в пространстве высокочастотные звуки (т. е. выше 3000 Гц).
Слуховая кора обезьяны исследована в меньшей степени, чем кора кошки, но уже теперь ясно, что частота представлена в А1 обезьяны также в виде изочастотных контуров, или полос, лежащих под прямым углом к линии, по которой расположены представительства участков улитки. Эти изочастотные полосы тянутся через все слои клеток коры в соответствии с колончатой организацией (Brugge, Merzenich, 1973). В этой связи большой интерес для нас представляет одно наблюдение этих исследователей. Они обнаружили в полосах для частоты 2500 Гц и ниже, что нейроны в колонках высокочувствительны к интерауральной задержке, а именно на этом свойстве (интерауральная задержка) основана способность локализовать звуки пизкой частоты в окружающем пространстве.
Суга (Suga, 1977) описывает колончатую организацию слуховой коры летучей мыши, отмечая те преимущества, какие дает возможность картирования нескольких переменных в двух измерениях. Эта летучая мышь испускает ориеитационные звуки, которые складываются из длительного компонента с постоянной частотой, за которым следует короткий компонент с переменной частотой. Первый компонент используется для обнаружения цели и измерения скорости ее движения по компенсации доилеров-ского сдвига. Второй компонент используется для локализации и определения величины цели. Самую интенсивную часть первого компонента составляет вторая гармо-
Организующий принцип функции мозга
ника около 61 кГц, которая снижается до 51 кГц за несколько последних миллисекунд сигнала. Около 30% первичной слуховой коры заняты колонками, строго специализированными на компонент с постоянной частотой во второй гармонике ориентационных звуков и ее эхо с доплеровским сдвигом. Колонки специализированы вдоль радиальной и концентрической осей по двум параметрам — частоте и амплитуде, и это представительство занимает непропорционально большую часть слуховой коры. Итак, первичная слуховая кора организована в виде колонок, и хотя данных еще недостаточно, но параметры идентификации по осям X и Y позволяют думать, что две пересекающиеся полосы колонок картируют частоту звука в нейронном выражении тех свойств стимула, которые нужны для локализации звука в пространстве. А таковыми являются интерауральиая разница в интенсивности для высоких частот и интерауральиая разница во времени для низких частот (ниже 2500 Гц). Для уверенного построения этой общей модели нужны дополнительные данные; несомненно также, что еще и другие динамические параметры тоже картированы вдоль оси равно-частотных полос колонок, но общее сходство в устройстве со зрительной и соматосенсорной корой совершенно очевидно.
Данные исследования прецентральной моторной коры
До недавнего времени прецентральная моторная кора была наиболее интенсивно изучаемой, но наименее понятной из гетеротипических областей новой коры. Значительные успехи в понимании функциональной организации этой области и ее роли в управлении движением достигнуты за последнее десятилетие благодаря применению новых методов. Среди них выделяются регистрация электрических знаков активности одиночных нейронов в моторной коре бодрствующих обезьян, обученных многократно совершать определенное движение (Evarts, 1975); внутрикорковая стимуляция и регистрация посредством проникающего микроэлектрода (Asanuma, 1973, 1975), а также некоторые новые способы прослеживания проекций к таким областям коры и от них. В результате воз-
В. Маунткасл
никла обширная литература, и я привлеку внимание читателя только к небольшой ее части, а именно к той, которая посвящена функциональной организации прецент-ральной моторной коры.
Асанума и сотр. (Asanuma, Rosen, 1972a, b) применили метод внутрикорковой микростимуляции и получили убедительные доказательства того, что те локусы в коре, стимуляция которых слабыми токами (4 мкА) вызывает небоиыпие движения в дпсталыюм суставе, нередко производимые одной только мышцей, — образуют вертикальные колонки диаметром 0,5—1,0 мм, которые соответствуют радиальным колонкам клеток, столь характерным для этой области. Им удалось также путем регистрации через стимулирующий электрод проследить афферентный вход к моторным клеткам коры в непосредственной близости от него. Эти клетки обычно активировались стимуляцией рецепторов в глубоких тканях в суставе и вокруг сустава, приводимого в движение локальной внутрикорковой стимуляцией. Только те колонки, раздражение которых вызывало движение пальцев, содержали нейроны с кожными рецептивными полями. Обычно эти поля находились на коже кисти, лишенной волос, в таких местах, где они активировались движением, вызываемым локальной стимуляцией. Составленная таким образом петля входа — выхода, как полагают, играет роль в управляемых тактильно движениях кисти и пальцев и в инстинк-тивпых хватательных движениях. Эти наблюдения в принципе подтверждены рядом исследователей (Doetsch, Gardner, 1972; Lenon, Porter, 1976). Другие авторы подчеркивают более распределенную природу «колоний» нейронов пирамидного тракта, связанных с определенной мышцей (Phillips, 1969; Anderssen et al., 1975). Существенный факт, который обнаружили Янковская и сотр. (Jankowska, Padel, Tanaka, 1975a), заключается в том, что, хотя возбужденные локусы для данной мышцы были распределены шире, чем можно было думать на основании наблюдения Асанумы, они не были непрерывными. Таким образом, согласно одной рабочей гипотезе о функциональной организации моторной коры, радиальные колонки нейронов, перерабатывающие входные сигналы в выходные, направленные к одной совокупности мотоней-
Организующий принцип функции мозга
ронов, образуют скопления, которые перекрываются там, где утончающийся край одного скопления встречается с краями скоплений колонок, связанных с другими мышцами. По этому представлению колончатая организация моторной коры и частично сдвинутое перекрывание, которое постулировали Филлипс (Phillips, 1969) и другие, полностью совместимы. Можно думать, что такие моторные корковые констелляции, управляющие движением, возникают из динамической комбинации непрерывно образующихся и распадающихся групп активных колонок, связанных с одной, двумя или многими мышцами, участвующими в определенной фазе движения. Считается, что организующие команды по формированию этих групп движений возникают где-то за пределами прецеитральнои моторной коры как таковой. Таким образом, оказывается лишним старый и часто повторяемый вопрос: «Думает ли кора в понятиях движений или же в понятиях мышц?» Моторная кора составляет промежуточный уровень в истинном джексоновском смысле: она вообще не «думает»; и движения, и мышцы «представлены» в ней, хотя и по-разному.
Асанума и Розен (Asanuma, Rosen, 1973) продвинулись еще дальше в изучении колонок моторной коры, применив два микроэлектрода — один для стимуляции и другой для отведения. Они обнаружили, что стимуляция верхних слоев вызывает возбуждение локально, а также в нижних слоях, в колонке диаметром несколько меньше 1 мм; эта же стимуляция создавала вокруг колонки зону торможения. Стимуляция в глубоких слоях вызывала локальное возбуждение и такое же торможение вокруг колонок, возможно идентичное по своему механизму тому, какое, по описанию Стефаниса и Джаспера (Stephanis, Jasper, 1964), создается импульсами в возвратных аксон-ных коллатералях клеток, дающих начало волокнам пирамидного тракта. Как это ни странно, стимуляция глубоких слоев не вызывала возбуждения нейронов в супра-гранулярных слоях.
Итак, моторная кора разбита на колонки клеток диаметром около 1 мм, но форма и геометрия этих колонок неизвестны. Группы колоно.к, связанных с определенными совокупностями вставочных или моторных нейродои,
В. Маунткасл
образуют скопления, края которых перекрываются с соседними скоплениями, связанными с другими мышцами. Параметром, определяющим их картирование по осям X и Y, по-видимому, служит их местонахождение на карте мышц тела. Можно предсказать, что и другие, более динамичные, свойства нейронной активности картированы по тем же осям X и Y, возможно, посредством дробления крупных колонок на более мелкие, пока что описываемые только топографически. Динамическое торможение вокруг колонок производится, по-видимому, как аксонами, входящими в серое вещество, так и действием возвратных коллатералей проекционных нейронов инфра-гранулярных слоев.
Данные исследования теменной гомотопической коры
Гомотипическая кора теменной доли обезьяны построена в виде вертикальных нейронных тяжей и обладает функциональными характеристиками колончатой организации (Mountcastle et al., 1975; Lynch et al., 1977). Эти области сильно отличаются от рассмотренных .выше гете-ротипических областей, так как они получают не один только афферентный вход, как это присуще первичным сенсорным областям, и не связаны однозначно с периферическими эффекторами, как моторная кора. Определяющие параметры для групп колонок в гомотипической теменной коре надо искать в опытах с электрофизиологическими наблюдениями на животных, обученных определенным поведенческим актам, относящимся к изучаемой зоне. При таком исследовании все группы колонок полей 5 и 7 проявляют одно общее определяющее их свойство: их клетки активны в связи с действием животного на непосредственно окружающую его среду и в ней, а также в связи с пространственными взаимоотношениями тела и его частей, гравитационного поля и этой среды (Mountcastle, 1975, 1976, 1977). В поле 7 имеются разные группы колонок, клетки которых активны при следующих движениях: 1) протягивание руки к интересующему предмету; 2) манипуляции с предметом; 3) фиксация взора и, следовательно, зрительное внимание; 4) сакка-дическое движение глаз, не спонтанное, а вызванное
Организующий принцип функции мозга
зрительным стимулом; 5) медленное следящее движение глаз и 6) внезапное появление предметов на периферии поля зрния (это единственные идентифицированные до настоящего времени нейроны теменной коры, которые являются «зрительными» в обычном смысле). Обособление в колонки этих групп клеток с совершенно разными свойствами доказывается тем, что: 1) микроэлектроды, вводимые в кору в направлении, перпендикулярном к ее поверхности, и вдоль вертикально расположенных колонок клеток, с большой вероятностью встретятся с клетками одного только класса и 2) микроэлектроды, вводимые наклонно по отношению к клеточным колонкам, проходят через блоки ткани, в которых все клетки относятся к тому или иному одному типу, а не перемешаны. Пока еще мало известно о точных размерах и форме колонок в теменной коре или о динамике нейронной активности в них. Я полагаю, что группы колонок каждого класса в теменной коре соединены специфическими внешними связями с аналогичным образом обособленными группами модулей в других областях коры, а также в подкорковых ядрах; эти тесно связанные модульные группы в разных крупных отделах мозга образуют точно соединенные друг с другом распределенные системы, обслуживающие распределенные функции.
Система центрального вещества, проецирующаяся на новую кору, без колончатой организации
Общая концепция колонок не исключает возможности того, что в кору проникают и действуют там разные другие системы, в особенности такие, которые выполняют общие регуляторные функции, а не детальную обработку информации. Примером может служить адренергическая система, которая заложена в locus coeruleus и очень широко проецируется на разные отделы центральной нервной системы, в том числе на всю новую кору (см. обзор Moore, Bloom, 1977). Эта система прямых проекций на кору обнаружена у многих видов животных, включая приматов. От locus coeruleus берут начало два идентифицируемых восходящих тракта, которые доходят до коры не прерываясь. Они идут вверх через субталамус; медиальный компонент достигает коры по поясному пучку, а лате-
В. Маунткасл
ральный — по наружной капсуле. Особое свойство этой системы состоит в том, что от двух точек вхождения ее волокна идут тангенциально и попадают во все зоны и слои коры. Молливер и др. (Molliver et al., 1977) применили для прослеживания этой системы иммуногистохимиче-ский метод, и на смонтированных ими реконструкциях видно, что новая кора пронизана по всем направлениям паутиной тонких норадренергических волокон с промежутками в 30—40 мкм. Все слои, кроме IV, содержат радиальные, тангенциальные или наклонные аксоны, расположенные так, что каждый норадренергический аксон может влиять на смежные соседние корковые колонки на очень большом расстоянии. По-видимому, любая клетка locus coeruleus имеет очень обширную проекцию в головном мозгу, включая области новой коры, и создает огромное разветвленное аксонное поле. Точно еще неизвестно, как оканчиваются эти волокна; это могут быть и традиционные сипаптические окончания, и участки, выделяющие медиатор en passage («по ходу»), но ясно одно: эта система способна оказывать прямое влияние на каждую клетку новой коры.
Функциональное зна.чение этой системы тоже неясно. Ее распределение позволяет думать, что она оказывает контролирующее или регулирующее действие на новую кору и что это может осуществляться по прямой синап-тической связи, или путем выделения синаптических ме-диаторных веществ «на расстоянии», или обоими этими способами, или же посредством регуляции кровотока и проницаемости сосудов (Raichle et al., 1975). Та роль, какую эта система, как известно, играет в механизмах сна, возможно, составляет лишь одну, наиболее очевидную из ее контрольных функций. Какова бы ни была природа ее влияния на кору — быть может, даже влияния на созревание коры, — в том, как она распределена по коре, нет никаких признаков «колончатой организации».
Внутренняя организация новой коры
Орзанизующий принцип функции мозга строить такую схему, хотя в работах последних лет получено много необходимых… Общий план представляется следующим образом. Афферентные волокна, приходящие в новую кору, идут из трех главных…Внешние связи новой коры
Организующий принцип функции мозга ратном направлениях, использован аксоплазматическии транспорт идентифицируемых… Накопленное сейчас огромное количество сведений о связях новой коры нельзя суммировать вкратце, однако теперь начинают…СЕЛЕКЦИЯ ГРУПП
И ФАЗНАЯ ПОВТОРНАЯ СИГНАЛИЗАЦИЯ;
ТЕОРИЯ ВЫСШИХ ФУНКЦИЙ
ГОЛОВНОГО МОЗГА
Поразительное разнообразие нервных систем у разных видов животных и их замечательная способность к адаптивной функции одновременно интригуют и… Но на функциональном уровне в основах и методологии все еще царит путаница. Во… Селекция групп и фазная повторная сигнализация 69Селекция и ее предпосылки
Селекция групп и фазная повторная сигнализация 71 но они не могут функционировать, пока их нейронные сети не получат от… Такая инструктивная модель высших функций мозга наталкивается на ряд трудностей. Инструкция требует точного…Требования к селекции групп; вырожденность
Селекция групп и фазная повторная сигнализация 75 ассортимент можно представить себе как собрание разнообразных нейронных групп,… Первое требование состоит в том, чтобы этот ассортимент был достаточно богатым; иными словами, он должен содержать…Вырожденная селекция групп в головном мозгу
Первый важный вопрос состоит в следующем: что распознает ЦНС? То есть, каков элементарный субстрат высших функций мозга? Мы примем здесь, что выше… Хотя о существовании клеточных групп в других областях имеются лишь скудные… Дж. ЭделтенЖ
Рис. 6. Фиксация ответов в вырожденной сети посредством комбинации дискриминативного торможения и коммитирования, повышающей вероятность ответа и приводящей к вхождению группы G3 ассортимента (Я для R) во вторичный ассортимент, что отмечено рамкой. Пупктирная стрелка — плохое распознавание; сплошная стрелка — хорошее распознавание; жирная стрелка — очень хорошее распознавание; двойная линия — коммитирование.
Селекция групп и фааная повторная сигнализация 89
Ниже мы обсудим это подробно в связи с условиями, которые требуются для сознания. Здесь же краткое рассмотрение ассоциативной памяти в вырожденных системах позволит подготовить почву для такого обсуждения.
Ассоциация предполагает такую программу, при которой предъявление разных признаков объекта на входе
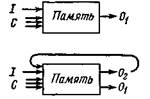
Рис. 7. Схема ассоциативной памяти, в которой входные (input, /) сигналы (например, сигналы от групп клеток) в контексте (context, С) (представляющем фон, торможение и др.) вызывают соответствующие выходные (output, О) сигналы. Такие выходные сигналы, как Ог, могут повторно поступать в такую систему.
приводит к образованию связи между этими признаками на выходе (рис. 7). Вся группа программ, находящаяся на хранении, может образовать ассоциативную память при наличии средств обращения к^ ней и считывания. В идеале предъявление на входе в надлежащем контексте любой совокупности находящихся на хранении элементов должно привести к вызову части или всей этой совокупности. При определенных условиях элементы не из этой совокупности также должны быть способны произвести вызов. Весьма вероятно, что в головном мозгу к содержанию хранения такого типа имеется адрес, и обращение происходит одновременно и параллельно.
Вырожденная совокупность клеточных групп обладает рядом черт, которые могут привести к ассоциации и породить память с такими свойствами. Некоторые группы нейронов могут распознавать данный сигнал только более или менее удовлетворительно. Кроме того, некоторые из этих групп могут распознать какой-либо другой сигнал лучше, чем тот, на какой они отвечали при определенном событии. Отсюда возникает возможность того, что два разных события вызовут реакции одной и той же группы, а
Дж. Эделтен
также одновременные реакции совершенно различных групп (рис. 8). Кроме того, если вероятность реакции всех этих групп изменится вследствие многократных предъявлений, то повысится вероятность их совместного
РдляЯ U U 1Гг U U
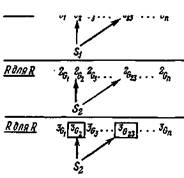
Рис. 8. Ассоциация, образуемая вырожденным распознаванием и коммитированием для двух независимых сигналов S π 52. Толстые и тонкие стрелки обозначают степень распознавания. Группы в рамках соответствуют коммитированию с переходом во вторичный
ассортимент.
возбуждения при будущих предъявлениях любого из этих сигналов. Поскольку эти группы могут иметь разные ней-роанатомические связи, ассоциация с другими группами, отвечающими на дополнительные сигналы, еще сильнее укрепится.
Предъявление вырожденному набору клеточных групп двух сходных, но не идентичных сигналов может стимулировать какую-либо общую часть групп этой совокупности, а также разные части, специфичные для каждого сигнала (рис. 9). Если общая часть связана с другими нейронными группами, возможна ассоциация реакций этих групп с реакцией общей части. Кроме того, последовательное предъявление двух связанных друг с другом сигналов может вызвать те же ответы, активируя эту часть, так что группы (R для R), считывая любую из этих частей, станут таким образом ассоциированными.
Существует дополнительная возможность того, что распознавание структуры клеточной группы потребует
Селекция групп и фазная повторная сигнализация 91
только части внутренне связанных между собой нейронов распознающей группы, оставляя вторую ее часть свободной для других распознаваний. Это особый случай, близкий к распознаванию двумя пространственно различными группами. Но благодаря локальным связям он создает
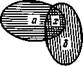
Рис. 9. Считывание с ассоциацией разных совокупностей групп, вызываемое двумя структурно сходными сигналами а и б. Совокупности групп клеток, распознающих сигналы, обозначены а ж б. Общая подгруппа реагирующих групп обозначена х. Ответы разных групп (R для R), считывающих эти сигналы, могут дать ассоциативную реакцию.
дальнейшие возможности для определенных видов ассоциации путем рекомбинации свойств внутри группы.
Рассматривая ассоциацию, мы должны также учитывать степень иерархического «вложения» (nesting), возможного при селективных распознаваниях в ассоциативных системах такого рода. Под вложением я имею в виду число последовательных распознаваний на разных уровнях организации, таких, как R и (7? для R). Хотя вложенное распознавание может иметь место, схема не имеет обязательного или необходимого иерархического ограничения. Подгруппа (R для R) может избирательно «распознать» другую подгруппу (R для R); распознавание не ограничено клеточными группами другого уровня, такими, как R. Это создает возможность считывания состояний клеточных групп, вероятность реакции которых стабильно изменена при предыдущих селекциях клеточными группами, которые не были так изменены. Ввиду вырожденного характера ассортимента это означает также, что данная структура может «храниться» в реакции нескольких или, возможно, многих изофункциональных, не обязательно изоморфных групп. Этим создается широкая возможность для ассоциативных взаимодействий групп (R для R) в рефлексивном распознавании. В то же время следует под-
Дж. Эделмен
черкнуть, что некоторые нейронные группы должны всегда оставаться в первичном ассортименте. Действительно, могут оказаться нейроны, неспособные к коммитирова-нию, и тогда группы, из которых они состоят, не оказываются фиксированными для данной формы ответа.
Еще одна важная общая черта системы вырожденной селекции групп состоит в ее дистрибутивном характере. Мало вероятно, что данное стабильное выбранное состояние или реакция на входной сигнал представлены только в одной группе клеток в одном месте. Действительно, вне специальных нейроанатомических ограничений нет необходимости в том, чтобы выбранные изофункциональные группы были смежными. Это согласуется с наблюдениями Лешли (Lashley, 1950), согласно которым перерезка и удаление областей коры не нарушают приобретенного поведения, однако это не значит, что его представление об эквипотенциальное™ правильно. В самом деле, можно думать, что в некоторых случаях специальная способность распознавания присуща лишь некоторым более крупным областям головного мозга в результате эволюционного отбора и требований эффективной коммуникации между нейронами. Это должно находить отражение в соответствии, которое обнаруживается между функциональной адаптацией и плотностью периферической иннервации, с одной стороны, и размерами и изощренностью центрального представительства этой периферии — с другой. Эта установленная картина распределенных нейронных групп согласуется с данными о грубой локализации функций, но, кроме того, находится в соответствии с распределением форм памяти и обучения. Она согласуется также с функциональным взаимодействием групп, далеко отстоящих друг от друга, при осуществлении сложных функций мозга, как показали данные о повреждении и удалении частей мозга (Лурия, 1973). Имеются данные о большой вариабельности порогов устойчивости разных групп клеток к разрушению ткани (Russel, Espir, 1961) v что опять-таки согласуется с представлением о вырожденной селекции групп.
Важно подчеркнуть, что в селективной системе вырожденность и многообразие важнее, чем абсолютная широта ассортимента. Тем не менее требование ограничения
Селекция групп и фазная повторная сигнализация 93
большого числа разнообразных групп клеток в вырожденном ассортименте (см. рис. 1) находится в согласии с тем фактом, что у наноцефалов не обнаружены типы нейронов, коренным образом отличные от нормальных (Seckel, 1950), и с тем, что высшие функции не коррелируют с суммарными данными о размерах мозга. Однако, если число клеток ниже определенного уровня и нет условий для образования определенных синаптических связей, набор либо не будет содержать достаточного числа вырожденных подгрупп, либо входные сигналы не будут эффективно его мобилизовать.
Ни дистрибутивные, ни ассоциативные свойства не являются принадлежностью исключительно селективных систем; созданы различные модели, обладающие этими свойствами (Languet-Higgins, Willshow, Berneman, 1970; Cooper, 1973). Но в модели вырожденной селекции групп внимание обращено не столько на общие свойства всей сети, сколько на характеристики клеточных групп. Этим особепно подчеркивается главная роль нейроанатомии и развития в построении отдельных первичных ассортиментов и отдельных видов связанных друг с другом клеточных групп.
Важно показать, как под влиянием представления о вырожденной селекции групп меняется понятие о памяти. Фиксация реакций клетки внутри группы или всей группы через посредство нескольких клеток может изменить селеюивные характеристики на любом уровне распознавания. В аспекте теории вырожденной селекции групп память — это не локальное свойство какой-либо отдельной области нервной системы, а общее выражение усиленного взаимодействия групп клеток, которые содержат избирательно коммитированные клетки и их синапсы. Если добавить к этому возможности связи по длинным путям, понятие об активном хранении с высокой ассоциативной способностью и представление о повторном входе сигналов, о котором речь пойдет ниже, то нет необходимости считать память свойством отдельной области или каким-то исключительным выражением некоторой высшей функции мозга. Каков бы ни был микроскопический или молекулярный механизм памяти или взаимодействия клеток (например, новые дендритные связи, метастабиль-
Дж. Эделмен
ные изменения мембраны и поверхности клетки на дендритных шипиках, молекулярные изменения синапсов), это свойство является обязательным следствием вырожденной селекции групп и поэтому должно быть свойством нейронов, когда они функционируют в составе групп. Считывание из памяти не составляет какой-либо особой проблемы; этот процесс не отличается от других форм сообщения между нейронами в вырожденной системе групп.
Что можно сказать о нейронном субстрате вырожденного ассортимента? Становится ясным, что в высших мозговых центрах, например в обонятельной луковице (Shepherd, 1972), закон динамической полярности, связанный с классической нейронной доктриной, недействителен. Взамен возникает картина весьма многообразных дендро-дендритных, аксодендритных, аксосоматических и денд-росоматических связей. Такую картину, вероятно, можно обнаружить и в других частях коры, и вместе с данными о взаимодействии нейронов типа Гольджи II и весьма значительным увеличением числа ЛНЦ в филогенезе и по мере восхождения по нервной оси (Rakic, 1975) она создает анатомическое представление, которое полиостью согласуется с вырожденной селекцией групп. Развитие новой коры, по-видимому, совершалось путем значительного увеличения числа клеточных колонок или единиц, а также их взаимосвязей. Наряду с анатомией областей коры (Chow, Leiman, 1970; Peters, Paley, Webster, 1976) строение некоторых областей лимбической и ретикулярной формаций (Isaacson, Pribram, 1975a) тоже согласуется со свойствами вырожденного ассортимента.
Поскольку распределение клеточных групп в ассортименте не случайно, имеются широкие возможности для индивидуальных вариаций в разных частях мозга и у разных индивидуумов. Тем пе менее, согласно данной теории, у разных особей мозг должен выполнять определенные распознавания одинаково эффективно, даже несмотря на то, что в их функциональной истории были отобраны разные изофункциональпые подгруппы (рис. 10). Поэтому следствием вырожденности является высокая степень многообразия и индивидуальности. Это приводит к редким или необычным флуктуациям в пейронных ассортиментах некоторых особей.
Селекция групп и фазная повторная сигнализация 95
Согласно излагаемой теории, эмбриогенез и развитие создают первый ассортимент; ранний период обучения и селективное взаимодействие ведут к образованию второго ассортимента посредством химического облегчения и процессов памяти. После нескольких селекции статистические
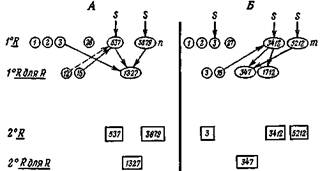
Рис. 10. Схема, иллюстрирующая индивидуальный характер следовой информации при установлении вторичного ассортимента у разных индивидуумов (А и В) в случае сходных первичных ассортиментов. Образование следов на разном нейроанатомическом фоне может изменить частоту встреч групп клеток, представленных в разном числе у разных индивидуумов. 1" R и 2° R — первичный и вторичный распознающие ассортименты; так же обозначены ассортименты (Я для Я).
отношения в первичном ассортименте меняются таким образом, что возникает вторичный ассортимент отобранных групп. Хотя тем самым меняются статистические отношения реакций ассортимента, все еще остается большое число возможностей. Группы клеток в этом вторичном ассортименте в общем с большей вероятностью будут повторно выбраны такими же входными сигналами, чем изо-фупкциональные группы первичного ассортимента. Следует, однако, подчеркнуть, что это лишь вероятностное предположение; при повторном предъявлении сигнала в зависимости от ряда условий могут быть выбраны предпочтительно группы первичного ассортимента, а не вторичного. Во всяком случае, в любой момепт группы, спо-
Дж. Эделмен
собные к специфическому распознаванию данного сигнала, в первичном ассортименте будут все же гораздо многочисленнее, чем во вторичном. Но вследствие предшествующего коммитирования пороговых событий не многие из этих групп будут так «тонко настроены» на данный сигнал.
Таким образом, строение центральной нейронной системы рассматривается как следствие двух разных селективных процессов: онтогенетического развития первого ассортимента и образования второго ассортимента путем селекции в процессе взаимодействия с окружающей средой. Соотношение переходных и критических периодов в обоих этих процессах, исследованные Блейкмором (В1а-kemore, Van Stuyters, 1974) для нейронов Хьюбела и Ви-зела и рассмотренные Пиаже (Piaget, 1950, 1954) для психологии раннего возраста и развития, заслуживают подробного изучения в свете этих представлений.
Позднее я сделаю несколько особых замечаний относительно эмбриогенеза первого ассортимента. А сейчас, пожалуй, важнее суммировать процесс образования второго ассортимента. Этот процесс ведет к образованию специфического следа селекционных событий, к повышению вероятности повторной селекции элементов второго ассортимента предпочтительно перед первым и к изменениям вероятности встреч, возможно, на уровне синапсов. В такой системе очень важен порядок развития второго ассортимента. Число клеточных групп, число клеток и синапсов в ЦНС создают массу возможностей образования второго ассортимента. Однако, независимо от числа селективных событий, все же в первом ассортименте остается большое, даже преобладающее количество некоммитиро-ванных клеточных групп.
Проведенный анализ имеет отношепие к двум ключевым положениям Маунткасла (Mauntcastle, 1976): «Центральная задача физиологии мозга состоит в том, как понимать действия крупных популяций нейронов, — действия, возможно, не полностью предсказуемые на основании свойств подгрупп», и «Центральная задача внутренней физиологии коры больших полушарий заключается в том, чтобы раскрыть природу нейронной обработки информации в проходящих через корковые слои цепочках
Селекция групп и фазная повторная сигнализация 97
взаимосвязанных клеток (в колонках)». В свете проведенного анализа эти высказывания можно перефразировать следующим образом: основная задача физиологии мозга состоит в том, чтобы понять природу построения ассортиментов из популяций клеточных групп. Разумеется, решение этой задачи требует знания передачи, мобилизации и встречи, а также усиления и стабилизации синаптических событий на молекулярном уровне. Предлагаемая теория является феноменологической в тдм смысле, что на уровне предпринятого здесь описания она не зависит от детального уточнения этих процессов; она просто утверждает, что множество разных решений совместимо с вырожденной селекцией групп.
Повторный вход селективной сигнализации и нервные субстраты сознания
Пока что наше обсуждение вырожденной селекции групп клеток не касалось конкретных цепей нейропных связей и их отношения к определенным высшим функциям мозга. Теперь я попытаюсь рассмотреть центральную проблему сознания и показать, как вырожденная селекция и сигнализация, производимая группами клеток путем повторного входа, создают условия, необходимые для объяснения этого феномена на клеточном уровне. До последнего времени проблема сознания не поддавалась экспериментальному анализу, и еще и теперь она представляет огромные трудности. Пожалуй, ни одна другая тема не вызывала к жизни столько самых различных спекуляций, сколько сознание: проблему разум — тело (Campbell, 1970), существование духов, очищение понятия «разум» посредством семантического анализа (Ryle, 1949) и возможность существования «главенствующего» нейрона (Sherrington, 1941). Общее мнение, несомненно, сходится на том, что эта функция мозга является своего рода центральной и должна быть понята для полного проникновения в обучение и другие высшие функции.
К счастью, теперь достигнуты некоторые успехи, позволяющие нам сузить предмет обсуждения. Опыты Пен-филда (Peniield, 1975), Джаспера (Jasper, 1966) и других (Isaacson, Pribram, 1975b) по роли ретикулярной форма-
Дж. Эделмен
ции в «пробуждении» (arousal) показывают, что и вне коры в головном мозгу существуют области, необходимые для сознания. С другой стороны, опыты Сперри (Sperry, 1970а) говорят об отдельной специфической локализации в полушариях разных функций мозга, связанных с сознанием. Кроме того, имеются данные о том, что система гиппокампа и лимбическая система участвуют в функции различения новизны и считывания кратковременной памяти, возможно модулируя тем самым активность на входе в сознательный мозг (Виноградова, 1975). Быть может, главным вкладом этой и других подобных работ является то, что созпание не составляет свойства всего головного мозга, а является результатом процессов, происходящих в некоторых определенных областях (предположительно обширных), например в коре больших полушарий, таламокортикальной радиации (Mountcastle, 1974) и лимбической и ретикулярной системах. Тем не менее характер взаимодействий, создающих сознание, еще не определен.
Поэтому вопрос состоит в следующем: каковы эти взаимодействия и какие свойства следует у них предполагать? Любая гипотеза, предлагаемая в качестве ответа на этот вопрос, разумеется, не должна противоречить законам термодинамики, постулировать вещи, которые не могут быть измерены, или вести к бесконечной регрессии определенных во времени проявлений распознавания. В своем положительном содержании гипотеза должна особенно выделять главную динамическую функцию головного мозга при переходе от приобретенного опыта к действию. При этом она должна объяснить обновление прошлого опыта, хранящегося в памяти, и временные параметры вызова. Возможно ли построить такую общую модель, включающую вырожденную селекцию групп, которая удовлетворяла бы этим ограничениям и учитывала бы временную последовательность состояний сознания и вызовов, необходимость обновления запаса информации (МасКау, 1970) и дифференцирование непосредственного и долговременного опыта. И в качестве предельного условия соответствовала ли бы такая модель различию между ■«я» и «не-я»?
Среди прочих условий такая модель должна специаль-
Селекция групп и фазная повторная сигнализация 99
но учитывать непрерывность восприятия, временную последовательность и обнаружение новизны. Первое решение, которое должно быть принято при построении этой модели, — это выбор между непрерывным и прерывистым временным ходом обработки информации. Есть ряд доводов в пользу прерывистого хода: 1) переработка информации о любом событии должна совершаться до осознания его (иными словами, некоторый выход должен быть подготовлен до принятия решения об исходе сенсорного входа); 2) существуют строгие временные ограничения вызова, которые требуют операций в реальном времени, и 3) как будет показано ниже, прерывистый способ значительно упрощает наше понимание того, как может кодирование пространственно-временной непрерывности происходить в вырожденной селективной системе. Эти и другие соображения, о которых будет сказано ниже, приводят к выводу, что элементарные процессы, создающие сознание, могут быть фазными, т. е. они могут требовать циклического повторения некоторой последовательности нейронных событий.
Подход к построению фазной модели показан на рис. 11. Предположим, что некоторая группа клеток в R получает сенсорную или сенсомоторную информацию и что ее действие и хранение распознаются несколькими группами клеток в (Я для R). Предположим одновременно, что тот же сенсорный входной приток в лимбические и ретикулярные центры обрабатывается и переключается таламусом в кору, чтобы стабилизировать данные группы (R для R) для нескольких немедленно следующих за этим событий. Стабилизацией, или фиксацией, я называю в этом контексте длительную импульсацию в этих группах с типичными для них последовательностями импульсов. Она составляет первую по времени фазу цикла обработки входных сигналов. Теперь предположим, что в следующей фазе этот обработанный сигнал повторно входит на более высоком уровне в проводящий путь информации S-*-R-*-(R для R) следующего цикла. Повторно вошедший сигнал и новые (R для R)-сигналы последующих входов считываются группами (R для R), которые образуют ассоциации со следами, хранящимися в (R дляR) группах клеток второго ассортимента, а также с груп-
пами в первом ассортименте, если имеется какая-либо новизна. Это составляет вторую, или повторную, фазу входной активности. Такая система построена так, что сигнал, возникший внутри, входит повторно, как если бы он был внешним сигналом. Это основная черта модели,
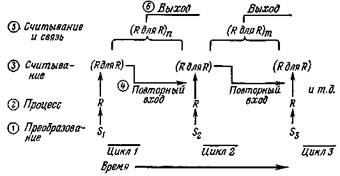
Рис. И. Повторный вход сигналов в путь, идущий от входов S к «распознавателям» (Я) и к «распознавателям для распознавателей» (Я для Я). На схеме показан повторный вход сигналов в последовательных циклах; порядок этих событий во времени в двух последовательных циклах обозначен цифрами. Ассоциативное значение повторного входа сигналов не показано (см. рис. 12). (Я для Я)п и (Я для Я)т означают ассоциативные нейронные группы высшего порядка, выходные сигналы которых могут повторно войти в дру-тих точках, активировать моторный выход, образовать ассоциации и т. п.
и она выполняет две функции: доставляет средство обработки новизны и создает соответствие и связь между внутренними состояниями и новыми сенсорными входными сигналами разных модальностей. В деталях она должна быть способна соотносить модальности и устанавливать кросс-корреляцию. Возможный способ выполнения этой задачи будет описан ниже.
Благодаря повторному входу эта модель обеспечивает непрерывность или связь между последовательными фазными входами. Поэтому не требуется никакого распознавания высшего порядка связи между состояниями пред-
Селекция групп и фазная повторная сигнализация 101
метов по мере того, как они регистрируются во времени. Надо иметь в виду следующий существенный момент: при вырожденности селективной системы и наличии адреса отсутствие повторного входа сделало бы невозможным ассоциирование последовательных свойств, если они абстрагированы во времени. Повторный вход служит гарантией того, что непрерывность в нейронном построении является обязательным следствием пространственно-временной непрерывности предметов.
Второе чрезвычайно важное свойство этой системы состоит в способности обеспечивать последовательность пли порядок ассоциированных событий (R для R) для последующего вызова. Хотя упорядоченная и точная последовательность не является обязательным условием для вызова, она должна быть принята во внимание. Мы полагаем, что порядок событий в контексте выражается в определенном порядке ассоциации, и таким образом вызов события 1 в структурах (R для R) необходим для вызова события 2 и т. д. Те же «состояния часов» системы, которые требуются для первоначальных интеграции и абстракции, затем используются для вызова этой последовательности в тех же величинах и масштабе реального времени. Ясно, однако, что эти часы создают лишь основу реального времени для данного процесса; вызов необязательно займет столько же времени, сколько первоначальные события. Истинную продолжительность цикла повторного входа в реальном времени мы рассмотрим ниже.
Новизна сигналов обрабатывается в этой системе путем сравнения повторных и новых сигналов цикла, а также благодаря наличию первичных и вторичных ассортиментов. Совершенпо новая информация должпа активировать первичный ассортимент. Соответствие и несоответствие сигнальной информации могут определяться путем различения между активацией вторичного и активацией первичного ассортиментов. Каким именно образом различаются ассортименты, не ясно, но параметров частоты или длительности латентных периодов может быть достаточно для различения нейронной активности каждого из них. Впрочем, нельзя пренебрегать еще одной возможностью, которая состоит в том, что для обнаружения новизны информация, заложенная во вторичном ассортименте, весь-
Дж. Эделмен
ма эффективно сравнивается с новыми входными сигналами.
Из-за отсутствия сведений об анатомических деталях трудно сделать выбор из разных цепей, согласующийся со схемой повторного входа сигналов. Но теория предполагает наличие некоторого минимального числа связей. Каждое место вхождения афферентации в головной мозг имеет преимущественно инвариантные связи прежде всего е ограниченным набором R, помещенным в первичной сенсорной коре. Затем происходит более или менее специализированная дивергенция на группы (R для R). В этом месте сенсорные этикетки исчезают, и ассоциативный повторный вход ведет к тому, что высшие функции выражаются многими классами групп (R для Я), которые больше не расположены последовательно. Важно, что та-ламо-кортикальные и кортико-таламические радиации содержат цепи, которые являются кандидатами на участие в схеме повторного входа (Mountcastle, 1974).
Здесь важно перечислить условия, необходимые для того, чтобы такая схема могла функционировать, и описать некоторые последствия ее работы. Примем на время, что для схемы имеются надлежащие анатомические отношения; тогда для схемы требуются следующие условия.
1. Распознавание R группами (й для R) в височной, лобной и префронтальной областях. При селективной схеме это, по-видимому, предполагает в качестве первой степени дивергенцию, а не конвергенцию нейронов.
2. Импульсная входная активность, соответствующая церебральным, таламо-кортикальным, гиппокампальным и лимбико-ретикулярным ритмам.
3. Подгруппы (R для R) во вторичном ассортименте со стабильно измененными синапсами и передаточными функциями, служащие выражением коммитированных комбинаций (pattern), которыми представлены в памяти прежние состояния.
4. Вырожденное распознавание (R для R) -групп клеток другими (R для R) -группами. На некотором уровне должна также иметь место конвергенция, чтобы осуществлялось «абстрактное суммирование» или превращение сложных комбинаций.
5. Стабилизация выбранных (R для R) -групп по мень-
Селекция групп и фазная повторцая сигнализация 103
шей мере на один цикл (согласно предлагаемой схеме, это происходит по лимбико-ретикулярным афферентам общим или специфическим образом).
6. Кратковременное хранение для удержания комбинаций (pattern) «внешних входов» (входное состояние 1) и потенциальная способность хранения повторных «внутренних входов» (входное состояние 2). Такое хранение делает возможным соответствие и ассоциацию, причем само оно является вырожденным и широко распределенным.
7. Связь выхода (R для R) с центральными состояниями и с комбинациями, хранящимися в памяти, участвующими в управлении движениями.
Эта модель не предполагает наличия бесконечного множества распознающих нейронных групп. Действительно, поскольку она зависит от сравнений между наличными входными сигналами и заложенными в память состояниями и поскольку предполагается наличие ассоциированных вырожденных групп со сходными распознающими свойствами, нет необходимости в том, чтобы в двух последовательных циклах входные сигналы распознавались одной и той же или одними и теми же группами (R для R). Вырожденность этих групп означает, что одно и то же входное состояние может быть распознано в разное время разными группами. Тем не менее важно, чтобы в пределах одного цикла использовались одни и те же (R для й)-группы, и перемена (R для й)-групп не должна прерывать выполнение двигательной выходной программы (pattern). Как будет подробнее рассмотрено ниже, предполагается, что осознание (awareness) возникает из ассортимента (R для R), имеющего доступ к R и к состояниям, заложенным в память и мультимодально порождаемым в нем самом. В свою очередь это обращение к памяти может привести к возникновению ассоциативно связанных сигналов, которые затем снова обрабатываются в тех же входных линиях, что и сигнал S; иными словами, эта система является системой повторного входа. Цикл в целом создает возможность модификации выходом (R для R) сенсорного входа и его порогов, а также возможность изменения состояния активации (arousal), намерения или внимания. Основным априорным условием
Дж. Эделмен
сознательного состояния является способность обозревать внутреннее состояние посредством непрерывного повторного поступления заложенной в память информации. Очевидно, что этот обзор модифицируется способностью распознавать новизну и состояниями активации.
До сих пор я предполагал, что сознательное состояние требует фазного распознавания вырожденными (R для R)-группами сигналов, представляющих внутреннее состояние организма и вход. Обсуждение было сосредоточено на несколько искусственном примере — на единичном сенсорном сигнале одной модальности. Но при этом не было должным образом освещено одно свойство вырожденных сетей, необходимое для их активности, связанной с сознательными состояниями: ассоциативный характер взаимодействия между вырожденными группами клеток. Многие нейронные группы, вызываемые сигналом данной структуры, лишь более или менее удовлетворительно соответствуют этой структуре. Кроме того, эти группы могут содержать нейронные конфигурации, которые тоже способны так или иначе распознавать другие, посторонние структуры сигналов; быть может, эти другие структуры будут распознаваться даже с большей вероятностью (или лучшей «пригонкой»), чем данная структура. Этим создается возможность того, что благодаря их «неиспользованной» потенциальной информации такие клеточные группы будут обладать ассоциативными свойствами, которые позволят им взаимодействовать с разнообразными сигналами, исходящими или от поступающей информации, или от заложенных в памяти (R для R). Таким образом, группа клеток может быть использована неоднократно разными сигналами или же одновременно двумя сигналами. И действительно, получены данные о мультисенсорном входе в данный корковый нейрон (Eccles, 1966b), хотя это может быть связано скорее с установлением уровня, чем с ассоциацией.
Ассоциативный характер системы повторного входа приводит к считыванию групп нейронов (R для R), которые представляют мультимодальный прошлый опыт. Согласно одному из вариантов модели (рис. 12), именно считывание этих групп в связи с текущей входной активностью имеет существенное значение для формирования
Селекция групп и фагная повторная сигналигация 105
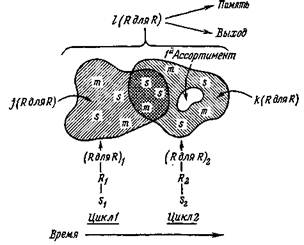
Рис. 12. На схеме показан повторный вход сигналов в двух последовательных циклах для иллюстрации ассоциативного мультимо-дального считывания совокупностей групп, содержащих заложенные в памяти следы прежних сенсорных входов (s) или сенсомо-торных событий (то). Это считывание завершается после стадий повторного входа, связывая таким образом две последовательные совокупности групп (Я для Я), отвечающих на Я. Здесь ;'(Я для R) и k(R для R) означают разное число групп, участвующих в ассоциативном ответе на (Я для Я), и (Я для Я)2. Перекрывание этих групп связано со сходством St и S2. Участок Iй Ассортимент означает ассоциативный приход к группам первичного ассортимента новой информации, содержащейся в S2. Нейроны I (R для R) могут сохранять результат, передавать его другим группам (Я для Я) или вызывать центральные программы двигательных реакций. Непрерывность зависит от сочетанного считывания (Я для fi)i и (Я для Я)2 и ассоциативных ответов в /(Я для Я) и k(R для Я). Новизна может быть обнаружена отчасти дифференциальной реакцией при считывании нейронными группами первичных и вторичных ассортиментов.
сознания. По поводу таких ассоциативных групп возникает вопрос, каким образом заложенные в память «правильные» (R для R)-группы становятся доступными для сравнения с Д-группами на выходе. Один из ответов на этот вопрос состоит в предположении, что (R для Д)-группы вторичного ассортимента распознают состояния Д
Дж. Эделмен
непосредственно. Это окажется возможным, если каждая такая группа будет обладать альтернативными конфигурациями, которые сделают возможными ответы на иные сигналы, кроме тех, для которых они были первоначально выбраны. Более специальная (и, как мне кажется, более привлекательная) альтернатива состоит в том, что (R для й)-группы, распознающие S через R, распознают также структуры других находящихся в памяти (R для R)-групп. Во всяком случае, извлечение из памяти и сравнение происходит никак не посредством случайной выборки; вернее, здесь играет роль густая сеть ассоциативных взаимодействий, которые усиливаются в вырожденной системе.
Этот ассоциативный потенциал, возможно, имеет большое значение также еще для одной характеристики ин-тактного головного мозга в связи с сознанием — одновременной параллельной переработки разнообразных входных сигналов разных модальностей. Можно представить себе, что при наличии нескольких входных сигналов Si, S2, S3... с ними со всеми произойдет то же, что с одиночным входом. Весьма вероятно, однако, что некоторые из них будут перекрываться во времени, хотя и асинхронно. Важная функция ассоциативной памяти может состоять в вызове синэстетических компонентов, связанных одновременным возникновением ассоциативных сигналов, идущих от параллельных и перекрывающихся сенсорных входов. Вследствие ассоциативного характера вырожденных сетей высока вероятность юго, что между (R для R)-группами установятся многочисленные взаимоотношения как на уровне хранения, так и на уровне считывания. Эти взаимоотношения создадут ряд выборочных ассоциаций между несколькими одновременно возникающими сигналами. Предполагается, что для состояния сознания имеет большое значение соотношение между этими мультимодальны-ми взаимодействиями и заложенными в память мультимо-дальными абстрактными структурами.
Выход к моторным системам и повторный вход из памяти непрерывно подтверждают или изменяют это соотношение. Наличие абстрактного представительства, которое, возможно, является следствием конвергенции афферент-пых путей на группах клеток в областях коры, выполняю-
Селекция групп и фазная повторная сигнализация 107
щих лингвистические функции, могло бы служить конденсации и интеграции прошлого опыта. Группы клеток, которые обслуживают такую функцию, вызываются для интеграции состояний сознания, возникающих из текущей входной активности путем ассоциативного взаимодействия. Все эти факторы должны способствовать объединению описанных ранее вероятностных, параллельных и различных по фазе процессов в богатую ассоциациями сеть взаимодействий, которая непрерывно меняется при изменениях на входе. И действительно, это должно сглаживать и связывать реакции и состояния сознания, создаваемые активностью разных (R для й)-групп. Это должно также способствовать быстро изменяющемуся обращению к большим блокам памяти с новыми ассоциациями в каждом цикле. Кроме того, как было указано выше, поскольку обращение к памяти зависит от цикла и «часа», упрощается грандиозная задача геперации памяти о продолжительности во времени данной совокупности вызванных событий: для вывоза и осознания используется одна и та же временная шкала.
Если такой анализ правилен, то субъект будет осознавать продолжительность, но не чередующиеся состояния цикла, так как состояния памяти, параллельная обработка других сигналов и асинхронность приведут к сглаживанию разных входов в течение больших периодов времени, чем единичный повторный цикл. Это подводит пас к критическому вопросу о периоде времени, отводимом для фазных состояний. Обнаружены гиппокампальные ритмы с частотами от 3 до 8 Гц, и можно представить себе, чш длительность циклов составит до 300 мс. Однако между переработкой информации и тета-ритмами не отмечается четкой корреляции (Bennett, 1975). Возможно, чго более подробные данные о взаимодействии во времени талами-ческого пейсмекера с восходящей ретикулярной системой дадут нам самую важную информацию (Mountcastle, 1974). Здесь имеет значение также и тот факт, что минимальный «период активации» для осознания околопорою-вого стимула составляет около 200—500 мс (Libet, 1966). Мне представляется, что возможен гораздо более быстрый вход; исследования по экспериментальной психологии показывают, что для полного восприятия время внутрикор-
Дж. Эделмен
новой переработки информации может не превышать 400 мс. Полезно было бы вычислить время, идущее на передачу и задержку, предполагаемые во всех частях постулированного здесь цикла, но сейчас из-за отсутствия подробных анатомических сведений такое вычисление невозможно. Впрочем, имеет смысл указать, что вслед за периодами такого порядка гиппокамп получает тормозные сигналы и происходит «стирание» (Pribram, Isaacson, 1975). Именно это и требуется в некоторых частях фазной схемы повторного входа. Кроме того, наличие такой кратковременной памяти, какую связывают с процессами в гиппокампе (Виноградова, 1975), также может иметь существенное значение для «подборки и удержания» фазной входной активности при описанной здесь обработке на высоком уровне.
При реакции на новизну головной мозг осуществляет центральную связь между приобретенным опытом и действием. Поскольку функция мозга в конечном счете состоит в том, чтобы управлять действием, то может показаться странным, что настоящее обсуждение не было сосредоточено на моторном выходе или функции. Однако понимание того факта (Evarts et al., 1971), что центральное программирование моторных действий, очевидно, преобладает над простыми рефлекторными актами, позволяет нам рассмотреть моторный репертуар почти так же, как это было сделано с сенсорным. Необходимо подчеркнуть то важное обстоятельство, что центральные состояния могут избирательно вызывать целые комбинации двигательной активности, рапее сформированные селективными процессами. Кроме того, по имеющимся данным (Vander-wolf, 1975), активность гиппокампа в состояниях внимания связана с осуществлением двигательных актов.
В сенсомоторпом взаимодействии, которое сосредоточивается на входе в селективную систему, имеется одна особенно важная черта: моторный репертуар направляет дальше, ограничивает и помогает программировать встречи в сенсорной сфере. Поэтому выполнение некоторых двигательных актов способно изменять плотность и характер входных сигналов и помогать уточнению селекции такими способами, которые иначе были бы маловероятными. Глубокий анализ влияния двигательных функций на
Селекция групп и фагная повторная сигнализация 109
парадигмы сознательного поведения можно найти у Мак-Кея (МасКау, 1966).
Подводя итог, стоит еще раз пересмотреть на примитивном уровне ту минимальную совокупность свойств, которые, согласно модели с повторным входом сигналов, абсолютно необходимы для состояния сознания. Требуются вырожденная селекция, взаимное рефлексивное распознавание нейронных (R для R)-групп, хранение (R для R), координированная обработка активации и сигналов S, ритмическая активность с фазовыми состояниями и соответствующие сети, удерживающие сигналы, что делает возможной повторную обработку икоординацию с внешним сенсорным входом хотя бы на некоторый короткий период времени. Можно ожидать, что как устранение входных сигналов от высших уровней среднего мозга, так и исключение хранения (R для R) или устранение сенсорного входа вызовет обширные расстройства сознания. Ясно, однако, что через некоторое время эта система не будет так жестко зависеть от стимула: благодаря состояниям памяти и проприоцептивным входным сигналам она будет продолжать функционировать, хотя и с нарушениями (Jasper, 1966). Надо представить себе, что (R для R), заложенные в память, находятся в состоянии непрерывной активности. Разумеется, изменение или отмена фазных возбуждающих и тормозных сигналов попеременной переработки повторных входов тоже имели бы серьезные последствия. В самом деле, одно забавное предсказание этой модели состоит в том, что сознание преобразуется в цифровую форму и отстает: во время входного состояния 1 осознание содержания сигнала еще невозможно. В этот период тестирование, если бы оно было осуществимо, не обнаружило бы осознания содержания сигнала. «Сознание» возможно только в состоянии входа 2.
Условия, достаточные для сознательного восприятия
Дж. Эделмен в лучшем случае посредством отчетов, но прямое сравнение сенсорных качеств,… Для начала стоит указать, что, поскольку сенсорные модальности передаются сигналами по «меченым линиям», в принципе не…Онтогенез и развитие первого ассортимента
Одно из главных допущений предлагаемой теории состоит в том, что развитию сознательного восприятия предшествует (и для него необходимо) образование вырожден-
Не
Дж. Эделмен
ного первичного ассортимента распознающих групп клеток. Поэтому следует задаться вопросом, как может развиться такой первый ассортимент в онтогенезе и как возникают его специфические свойства в процессе эволюции. Здесь я коротко рассмотрю проблему онтогенеза, а ниже сделаю несколько сравнительных замечаний о филогенезе селективных систем распознавания.
Проблема эмбриогенеза, в особенности гистотипических взаимодействий, в настоящее время далека от своего решения на молекулярном уровне. Во всей сложности она поставлена развитием нервных систем высшего уровня — например, в развитии такой системы, как ретинотекталь-ная проекция, проблема «схемы соединений» представляется грандиозной (Barondes, 1976). Один из способов решения этой проблемы очень прост — это постулирование для каждой пары клеток набора специфических дополнительных распознающих молекул, появление которых на соответствующем этапе развития запрограммировано генетически (Moscona, 1974). Однако все большее число данных говорит о том, что такого предварительного ассортимента фиксированных продуктов генетического процесса недостаточно. Как эмбриологические исследования (Hamburger, 1970), так и опыты по регенерации (Weiss, 1970) показывают, например, что нервно-мышечные взаимодействия не предетерминированы, а формируются с течением времени.
Полученные данные показывают, что после начального множественного образования синапсов происходит их конкурентное исключение, в результате которого отношение между аксонами и мышечными концевыми пластинками обычно становится равным 1:1. Кроме того, имеются данные, что при образовании некоторых нервных сплетений конечностей ветвление происходит до возникновения нервно-мышечного взаимодействия (Landmesser, Pilar, 1970, 1972). После частичного удаления tectum opticum в нем сохраняется возможность образования соответствующих крупных проекций (Yoon, 1975), что свидетельствует об отсутствии заранее фиксированного кода взаимодействия между отдельными клетками и нервными волокнами. Наконец, химические анализы областей адгезии клеток сетчатки (Brackenbury et al., 1977; Thiery et al., 1977) свиде-
Селекция групп и фазная повторная сигнализация 117
тельствуют о наличии одной или нескольких поверхностных белковых молекул, а не об обширном ретиноспецифи-ческом или нейронпоспецифическом ассортименте молекул.
Поэтому остается "в силе вопрос: как формируется первичный ассортимент? Я предлагаю здесь элементы предварительной гипотезы о развитии первичного ассортимента, свойства которого согласуются с последующей вырожденной селекцией групп для формирования вторичного ассортимента.
1. На ранних этапах онтогенеза группы клеток, или поликлоны, детерминированы генетически.
2. В эту систему встраивается высокая степень вырожденности — число клеточных групп значительно больше, чем в конечном счете используется для образования проекций и связей. Многие «неиспользуемые» клетки погибают (Prestige, 1970; Cowan, 1973).
3. Создается иерархия взаимодействий, причем на раннем этапе решения (например, ветвление нейронов в сплетениях конечностей) принимаются сравнительно редко и обусловливаются генетической программой.
4. Взаимодействия между группами клеток последовательные, избирательные и определяемые взаимным влиянием. Так, иннервация конечности тем или иным сегментом спинного мозга происходит при нервно-мышечных взаимодействиях и образовании синапсов, которое зависит
-от антероградных и ретроградных сигналов. Из нескольких первоначально сформированных синапсов обычно выживает только один. Это избирательное выживание согласуется с высказываниями Вайса (Weiss, 1970), а также Шанже и Денчина (Changeux, Danchin, 1976). Конечным результатом такого избирательного взаимодействия являются утрата синаптических связей и гибель многих нейронов ЦНС в процессе развития. Однако после такой селективной стабилизации (Changeux, Danchin, 1976) сохраняется высокая степень избыточности и вырожденности специфических изофункциональных клеточных групп.
5. Поэтому главное формирование окончательных связей на уровне синапсов рассматривается как избирательное взаимодействие, основанное на некоторых функциональных аспектах. После того как эти связи сформирова-
Дж. Эделмен
лись, они в общем не меняются. Но, несмотря на ее селективную природу, многое в этой системе все же генетически предопределено — положение ядер, путь их диффе-ренцировки и общая локализация. Так, колонки доминирования правого или левого глаза в общем предетермини-рованы, но могут быть приведены в действие в критическом периоде (Blakemore, Van Sluyters, 1974), а конкуренция правого и левого глаз заканчивается на уровне формирования синапсов с отдельными нейронами стриарной коры.
Согласно предлагаемой гипотезе, первый ассортимент, как и второй, селективно формируется на уровне своей тонкой структуры. Но даже если это и правильно, то сначала многое должно быть детерминировано генетической программой, и последовательные этапы развития являются особо важными, критическими (Bodian, 1970). Отличительный элемент гипотезы состоит в том, что вырожденность и селекция в значительной степени присущи системе уже на ранней стадии, даже еще до того, как завершено формирование первичного ассортимента. Дальнейшее развитие, особенно в критические периоды, может уменьшить эту вырожденность селективной синаптической стабилизацией (Changeux, Danchin, 1976). По мере восхождения по нервной оси можно ждать все более поздних эффектов критической селекции и все больше и больше свидетельств вырожденности. В некоторых субсистемах вырожденность не столь уж необходима и эволюционный отбор, а также формирование во время критических периодов могут устранить ее почти полностью. Но в связи с необходимостью развития второго ассортимента для высших функций мозга требуется сохранность большей части вырожденности в корковых и лимбико-ретикуляр-ных областях. С этой точки зрения для префронтальной, лобной и височной областей критический период длится непрерывно, а у некоторых счастливых индивидуумов — до самой смерти.
Эволюция вырожденных систем, коры и локальных нейронных цепей
Селекция групп и фазная повторная сигнализация 119 рожденных ассортиментах. Так, весьма вероятно, что у насекомых (Bullock,… Для некоторых видов выживание в этих условиях совершенно очевидно было связано с постепенным формированием глаза,…Некоторые предсказанияи следствия
1. Основными единицами селекции в функциях высших отделов головного мозга являются группы клеток, а не отдельные клетки. 2. Такие группы многократно повторяются, они являются вырожденными и… 3. В то же время множество входов от нейронов R и (R для R) конвергирует на одну и ту же группу клеток (R для R), чем…Резюме
Рассмотрение свойств центральной нервной системы у высших млекопитающих и особенно у человека говорит о том, что удовлетворительная теория функций головного мозга должна учитывать распределенный характер обучения, ассоциативный характер вызова из памяти, адаптивную реакцию на новизну и способность к весьма абстрактным представлениям в модели мира. Этим условиям, очевидно, удовлетворяет селективная теория функции головного мозга, в которой единицей отбора (селекции) служит группа нейронов.
Теория предполагает, что в эмбриогенезе и во время развития образуются группы нейронов с определенной структурой, содержащие до 10 000 клеток. Внутренние связи в группе и внешние связи между группами определяются генетической программой и синаптической селекцией. В результате этого процесса образуются первичные ассортименты (наборы), состоящие из групп нейронов различного строения и с различными связями, из которых более чем одна способна реагировать на данную форму (pattern) сигнала или распознавать ее. Такая реакция по принципу отношения «много к одному» означает, что все ассортименты являются вырожденными. Мобилизация таких вырожденных первичных ассортиментов сигналами приводит к ассоциативному распознаванию. Кроме того, в результате повторения сигналов, взаимодействующих с отобранными нейронными группами, создаются вторичные ассортименты групп с более высокой вероятностью реакции. Такой процесс селективен в том смысле, что сигналы не играют роли в образовании анатомических связей в группах первичного ассортимента, а лишь отбирают соответствующие группы из ранее образованного ассортимента.
Эта селективная теория высших функций мозга не требует специальных термодинамических допущений и свободна от менталистических представлений. Но поскольку предлагаемая система селективная и вырожденная, она не детерминирована механистически в том смысле, в каком детерминирована машина с часовым механизмом. На основе теории вырожденной селекции групп сделана по-
Селекция групп и фазная повторная сигнализация 125
пытка объяснить свойства сознания при помощи феноменологической теории, согласующейся с нейроанатомиче-скими и нейрофизиологическими фактами. К требующим объяснения свойствам сознания относятся: 1) способность оценивать или различать разные события; 2) способность реагировать критически на внутренние и внешние состояния и обновлять информацию; 3) способность накапливать следы в памяти и вызывать их посредством ассоциаций во временных последовательностях и 4) способность отличать «я» от «не-я» (осознание себя). Удовлетворительная гипотеза, объясняющая эти свойства, должна избегать бесконечной регрессии иерархических состояний и должна создавать возможность предварительного планирования и моторного выхода без программиста, т. е. должна снижать потребность в программировании.
Предложенная здесь гипотеза заключается в том, что состояние сознания возникает в результате фазного повторного входа сигналов, происходящего в параллельных процессах, которые включают в себя ассоциации между заложенными в память формами (patterns) сигналов и текущей сенсорной или внутренней входной сигнализацией. Каждый фазный процесс делится на две стадии. Первая стадия состоит из входа обработанных входных сигналов и удержания возникших в результате и ассоциированных сигналов для повторного входа на второй стадии. На этой стадии последующий обработанный входной сигнал соединяется с повторными входными сигналами и ассоциируется с реакциями групп как в первичном, так и во вторичном ассортиментах. Этот цикл завершается в течение миллисекунд. Предполагается, что осознание возникает в результате обращения групп нейронов высшего порядка к богатым мультимодальным ассоциативным структурам, заложенным в долговремепную память в результате прошлого опыта.
В этой теории сделано допущение, что временная основа повторного входа и временная основа вызова одни и те же. В сочетании с последовательным маркированием заложенных в памяти событий это дает возможность вызова по надлежащей временной шкале и в надлежащем порядке. Благодаря селективному, вырожденно-групповому и фазному характеру циклов повторного входа и способ-
Дж. Эделмен
ности нейронов высшего порядка к абстракции последовательности событий может быть создана непрерывная сдвинутая комбинация ассоциаций. После достаточного развития и опыта такая система может приобрести способность различать абстрактные комплексы, т. е. отличать «я» от входных сигналов окружающей среды. Хотя такая система не зависит от фиксированной программы или программиста, ее свойства согласуются со способностью головного мозга к выполнению сложных операций.
Исходя из этого представления, сознание рассматривается как форма ассоциативного воспоминания с обновлением, основанным на текущей повторной входной сигнализации, которая непрерывно подтверждает или изменяет «модель мира» или «теорию я» посредством параллельных моторных или сенсорных выходов. Весь процесс зависит от свойств селекции групп и повторного входа сигналов в нервной системе, которая уже специализирована в эмбриогенезе и в эволюции.
Благодарность
ЛИТЕРАТУРА Barondes S., ed., 1976. Neuronal Recognition, New York, Plenum Press. Bennett T. L., 1975. The electrical activity of the hippocampus and– Конец работы –
Используемые теги: Разумный, мозг, Кортикальная, Организация, Селекция, групп, Теории, высших, функций, головного, мозга0.15
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Разумный мозг. Кортикальная организация и селекция групп в теории высших функций головного мозга
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.245 сек.
Новости и инфо для студентов