рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Медицина
- /
- Физиология рефлексов
Реферат Курсовая Конспект
Физиология рефлексов
Физиология рефлексов - раздел Медицина, Текст Взят С Психологического Сай...
Текст взят с психологического сайта http://www.myword.ru
На данный момент в библиотеке MyWord.ru опубликовано более 2000 книг по психологии. Библиотека постоянно пополняется. Учитесь учиться. Удачи! Да и пребудет с Вами.... :)F Л АВ А 1
Односторонняя передача и синоптическая задержка Одна из наиболее известных черт синаптической передачи — ее направленный… ПРИНЦИПЫ НЕРВНОЙ ИНТЕГРАЦИИ 41И
ГЛАВА I
нерва не вызывает очевидных изменений эфферентного разряда. Само по себе раздражение может быть под-пороговым в отношении способности возбуждать мото-нейроны. Но при действии совместно с эффективным раздражением это «подпороговое» раздражение усиливает его эффект. Тем самым становится очевидным, что раздражение не было подпороговым для рецепторов, но разряд последних был недостаточен для создания выраженного сдвига возбудимости центральных нейронов или по крайней мере мотонейронов. Может быть и так: два афферентных раздражителя, ни один из которых сам по себе не способен вызвать возбуждение мотонейронов, при совместном воздействии вызывают рефлекторный ответ. Это показывает, что каждое раздражение вызывает облегчение центральных механизмов и что конвергирующие эффекты облегчения складываются — центральная суммация.
Эффект отдачи и разряд последействия
Возбуждающий и тормозной разряды последействия могут быть весьма продолжительными вследствие существования в центральной нервной системе открытых… * * * * 1 Торможение не имеет смысла, если оно не снижает активность, или возбудимость.L
ГЛАВА II
части мозга (ствол) можно рассматривать как продолжение сегментарной системы. Но, кроме того, развивается ряд структур, которые лишены признаков сегментарное™ и которые достигают доминирующего положения чо отношению ко всем частям сегментарной системы, обеспечивая более совершенные механизмы общей интеграции в масштабе организма. Поэтому их называют надсегментарными. Наиболее примитивный из этих механизмов— ретикулярная формация, которая оказывает облегчающее и тормозящее .влияние «а различные сегментарные и межсегментарные нейроны. Уже на очень раннем этапе эволюции эта формация представляла собой сеть, объединяющую отдельные рефлекторные дуги, каждая из которых обеспечивает лишь локальную реакцию на локальное раздражение. Анатомически ретикулярная формация производит впечатление наименее специализированной нервной ткани. Это лабиринт взаимно переплетающихся волокон, между которыми довольно равномерно расположены тела клеток. Поэтому ее нельзя отнести ни к серому веществу, которое состоит главным образом из тел нейронов, ни к белому веществу, которое образовано нервными волокнами и не содержит самих клеток. Это ретикулум, или сеть, с многочисленными контактирующими между собой нейронами, имеющими короткие отростки. Деление на серое и белое вещество происходит по мере специализации функций; ретикулярную формацию можно рассматривать как основу, своего рода матрикс, для развития обоих этих веществ, с которым они сохраняют тесную связь даже у. человека. Чрезвычайно существенно, что ретикулярная формация, которая прежде всех остальных видов центральной нервной ткани получает импульсы от всех видов рецепторов на всех уровнях и образует функциональные связи со всеми эфферентными путями к эффекторам, остается хорошо развитым образованием даже у наиболее высокоспециализированных животных и сохраняет с рецепторами и эффекторами как прямые, так и непрямые связи — через скопления серого вещества и пучки волокон. В действительности ее объем, диапазон ее связей, а быть может, ,и значение возрастают, хотя до последних лет внимание исследователей было приковано
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
S?
к относительно более развитым структурам —серому веществу коры, подкорковым ядрам и проводящим путям. Еще совсем недавно на ретикулярную формацию смотрели как на остаток «сырья», из которого возникли важные части нервной системы, как на своего рода «наполнитель», не заслуживающий специального изучедия. В последнее время оценка роли ретикулярной формации в корне изменилась, и это в общем справедливо, так как на самом деле ретикулярная формация не только представляет собой материал, из которого развились более специализированные интегрирующие механизмы, но и сама является весьма важным интегратором.
Среди ядер, раньше других образовавшихся из ретикулярной формации, находятся так называемые жизненно важные центры, такие, как дыхательный я сосудо-двигательный. Они не очень отчетливо обособлены от остальной массы ретикулярной формации, но характеризуются несколько более высокой концентрацией нейронов и особым богатством связен с мускулатурой дыхательной и циркуляторной систем. Развитие многих других специализированных скоплений в ретикулярной формации у высших животных происходит параллельно с интенсивным ростом коры мозга и мозжечка.
Особенно интересно проследить развитие ладсегмеи-тарных центров, связанных с образованием специализированных органов чувств, которые сообщают чрезвычайно важную информацию организму в целом, а не отдельным его частям. Эти центры образуются на головном конце организма, т. е. в области первого контакта с меняющейся средой при движении животного. Наибольшее значение имеют дистантные рецепторы (телерецепторы), которые улавливают энергию, .выделяемую объектами, задолго до того, как животное вступает в непосредственный контакт с ними. Таковы обонятельный, зрительный и слуховой рецепторы.
Тот, кто изучает нервную систему в филогенезе, не может не поражаться огромному множеству экспериментов, которые были поставлены природой в процессе развития такого типа нервной организации, который был бы наилучшим образом приспособлен для оптимального использования информации, получаемой через специаль-
ГЛАВА If
ные рецепторы, расположенные на голове, в сочетании с информацией, получаемой от глубоких и поверхностных рецепторов тела. Каждый из специализированных органов чувств — органы равновесия, слуха, зрения и обоняния (о вкусовой чувствительности известно мало) — связан с первичной проекционной областью коры '. Эфферентные пути, идущие от этих областей к ретикулярной формации, обеспечивают возможность известной регуляции со стороны каждого из этих механизмов.
Органы равновесия в вестибулярном аппарате внутреннего уха обеспечивают рефлекторным путем ориентацию животного по отношению к горизонтальной плоскости. Кора мозжечка, получающая от них импульсы, относится к древнейшей его части, которая у человека сравнительно мала и широко заменяема по функции. В значительной мере поддержание равновесия у человека осуществляется на основе зрительной информации, и, хотя вестибулярный аппарат и древний мозжечок участвуют в регуляции позы и равновесия, это участие, как правило, не является жизненно необходимым. Следует упомянуть, что органы равновесия и древний мозжечок тесно связаны с ретикулярной формацией, а также с ядрами вестибулярного нерва, которые представляют собой скопления клеток «ретикулярного» происхождения. Хотя разрушение этого примитивного механизма равновесия у человека приводит временно к тяжелым расстройствам, постепенно происходит почти полное восстановление регуляции равновесия, осуществляемой через другие органы чувств, особенно с помощью зрения.
Примитивный слуховой мозг не достигает высокого ' уровня развития ни у одного из видов животных, хотя у птиц и рептилий, у которых слуховые области новой коры неразвиты, он является высшим слуховым ядерным образованием. У человека первичный слуховой центр представлен нижним двухолмием, размеры которого довольно малы. Через него осуществляется рефлекторное сокращение мышц среднего уха и рефлекторный поворот
* * * *
1 Подробное объяснение развития первичных коррелирующих центров читатель найдет у Эллиота [220].
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 59
глаз и головы к источнику неожиданного звука. Внезапный громкий шум вызывает старт-рефлекс, который охватывает и другие группы мышц. Однако нет основания считать, что нижнее двухолмие имеет большое значение для нервной интеграции у человека.
Примитивный зрительный мозг явно выражен уже у костистых рыб и птиц; у человека же он представлен слаборазвитым верхним двухолмием. Глаз получает огромное количество информации, и при наличии адекватного нервного механизма и эффекторов для ее использования она имеет первостепенное значение в определении поведения организма. У человека филогенетически более поздние зрительные центры, локализующиеся в коре, в значительной степени взяли на себя осуществление зрительных рефлексов и вместе с другими кортикальными областями произвольный контроль над эффекторами, участвующими сво многих из них. Например, при рефлекторной аккомодации, регулирующей толщину хрусталика, степень конвергенции глаз и диаметр зрачка при рассматривании объектов, находящихся на различном расстоянии от сетчатки, используются пути, подходящие к зрительной коре. Однако некоторые рефлексы интегрируются верхним двухолмием среднего мозга. Защитное смыкание век в ответ на внезапную вспышку света, поворот глаз и головы в сторону объектов, попадающих в периферическую часть поля зрения, зажмуривание и отдергивание головы .при внезапном приближении объекта— эти акты, очевидно, быстро осуществляются через прямые рефлекторные пути среднего мозга.
Примитивный обонятельный мозг, очевидно, имел большее значение для выживания наземных позвоночных, чем остальные рано возникшие рецепторные зоны коры. На ранних стадиях эволюции и у эмбрионов высших млекопитающих полушария образованы почти полностью обонятельным мозгом [220]. У человека и высших животных возникает массивная новая кора (неокор-текс), которая оттесняет древний обонятельный мозг. Он занимает значительно меньше места, его строение при этом изменяется, он переходит на положение второстепенной структуры соответственно той скромной роли, какую играет обонятельная информация в интеграции. Од-
ГЛАВА П,
нако обонятельный мозг составляет чрезвычайно важную часть нервного механизма, участвующего в организации эмоций, и мы тщательно рассмотрим его структуру и функции в следующей главе.
Ни одна из перечисленных нами примитивных структур не оказалась оптимальным механизмом для общей интеграции. Координация сенсорной информации в целом требовала центрального механизма. Каждый отдельный сенсорный орган обеспечивает полезную информацию, «о поведение, направляемое каким-либо одним из них, более стереотипно и в меньшей мере адаптивно, чем в том случае, если оно формируется на основе суммарной информации от всех органов чувств плюс поступающие по каналам обратной связи «текущие отчеты» от различных частей организма. Таламус — своего рода «ворота в кору» — служит центральным коллектором различных видов сенсорной информации. Расположенное рядом полосатое тело, очевидно, служило в филогенезе, до развития новой коры, основным выходом моторных сигналов из таламуса. Таким образом, таламо-стри-арную систему можно рассматривать как сенсо-мотор-нуго интегрирующую систему, хотя и примитивную, перешагнувшую в своем развитии ту стадию, на которой отдельные органы чувств функционируют как относительно независимые системы.
Наряду с участием таламо-стриарной системы в интеграции соматических функций гипоталамическая область промежуточного мозга служила (и служит) регулятором висцеральной интеграции. Ее тесную связь с лимбиче-сними структурами, в том числе обонятельными, мы рассмотрим в следующей главе, а здесь лишь укажем, что к ней идут сигналы от всех внутренних органов и что импульсы, посылаемые ею через ретикулярную формацию и связанные с нею структуры, влияют как на соматические, так и на висцеральные функции. Хотя ее основная функция состоит в регуляции гомеостаза внутренней среды, эта цель- не может быть достигнута посредством одной только вегетативной регуляции. Параллельно с основной задачей поддержания физических и химических условий, необходимых для жизни и функционирования скелетной мускулатуры, должна существовать
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 61
и способность в известной мере определять уровень этой активности. Мы увидим далее, что гипоталамус имеет важное значение для регуляции не только висцеральных, но и соматических функций.
Хотя таламус и представляет собой чрезвычайно важное образование, поведение «таламических» животных в высшей степени стереотипно и почти лишено' признаков интеллекта. Хорошим примером может служить лягушка. Большая гибкость в создании новых типов реакций «а основе обширной и самой разнообразной информации требует системы со значительно большими возможностями для избирательной активации различных комбинаций эфферентных путей. Бесчисленные преобразования и комбинации в мозгу человека возможны благодаря существованию огромного лабиринта кортикальных нейронов с их многочисленными ветвями и богатыми си-наптическими связями. Только кора, где достаточно места для широкого распространения процессов, обладает способностью к созданию многочисленных потенциальных функциональных связей, столь необходимых для обучения и проявления «разумного» поведения.
СОМАТИЧЕСКАЯ И ВЕГЕТАТИВНАЯ СИСТЕМЫ
Четкую грань между висцеральным и соматическим отделами нервной системы можно провести лишь в отношении эфферентных нервных волокон. Те из них, которые иняервируют клетки скелетных мышц, безусловно являются соматическими, а те, которые иннервируют внутренние органы, составляют вегетативную, или, по определению Лэнгли, автономную нервную систему. Термин автономный не вполне правомочен, так как хотя функции этих нервов в значительной мере независимы от волевого контроля, степень этой независимости не столь велика, чтобы можно было говорить об автономии. Так, рефлекторная реакция, возникающая при раздражении интерорецепторов, не ограничивается висцеральными сдвигами, регулируемыми вегетативными эфферентными нервами; рефлекторные реакции, вызываемые раздражением соматических рецепторов, также не ограничиваются соматическими эффекторами. Например, холод
ГЛАВА II
раздражает рецепторы кожи, возбуждение которых ведет к обширным рефлекторным висцеральным сдвигам, сопровождающимся уменьшением теплоотдачи. Один из таких сдвигов — сокращение гладких мышц в стенках кожных кровеносных сосудов, их сужение и отведение крови от кожи к глубоко расположенным областям тела, вследствие чего понижается температура кожи и уменьшается теплоотдача путем излучения. Произвольно индивидуум может перейти в более теплое место, надеть пальто или начать двигаться. Мышечная дрожь — совершенно непроизвольное повышение активности скелетных мышц — является весьма эффективным механизмом теплопродукции, который трудно отнести к чисто автономным или чисто соматическим эффектам; это результат действия импульсов, проводимых по соматическим нервам к скелетным мышцам, однако он не менее «автономен», чем любой из других, несомненно вегетативных видов активности.
Существует важное анатомическое различие между вегетативной и соматической нервной системой в способе соединения центральной нервной системы с эффекторами. Тело соматического мотонейрона расположено в пределах центральной нервной системы, а его аксон направляется к волокнам иннервируемой им скелетной мышцы. Особенность .вегетативной нервной системы состоит в том, что между центром и иннервируемым эффектором находится цепочка из двух нейронов. Тело первого лежит в головном или спинном мозге, а его аксон оканчивается в клетках вегетативных ганглиев, лежащих вне мозга; и уже аксоны этих клеток оканчиваются в области нервно-эффекторного соединения.
Хотя, как было указано вначале, вегетативная нервная система является чисто эфферентной, как правило, принято говорить о вегетативных центрах и рефлексах. При рефлекторном изменении активности висцерального эффектора эфферентным звеном служат волокна вегетативной нервной системы, и мы поэтому говорим о вегетативных рефлексах независимо от того, находятся ли соответствующие рецепторы в висцеральных или соматических структурах. Таким образом, в широком смысле слова мы можем относить к вегетативной нервной систе-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 63
ме те ее части, которые влияют на внутренние Органы. Речь идет обо всех органах, в которых эффекторы представлены гладкими мышцами или железами. Пищеварительные железы, потовые железы, слезные железы, мочевой пузырь, желудочно-кишечный тракт, сердце, кровеносные сосуды, радужная оболочка глаза иннервируются вегетативной системой. Кроме того, вегетативная система иннервирует многие железы внутренней секреции, такие, как мозговое вещество надпочечников и островки поджелудочной железы.
СИМПАТИЧЕСКАЯ И ПАРАСИМПАТИЧЕСКАЯ СИСТЕМЫ
Подразделение вегетативной системы на эти два отдела основано на анатомических и физиологических данных. Наиболее важное анатомическое различие заключается, вероятно, в локализации ганглиев. Ганглии симпатического отдела лежат, как правило, вблизи грудного и поясничного отделов спинного мозга; отсюда и название торако-люмбальная, часто прилагаемое к симпатической системе. Поскольку синапсы находятся вблизи спинного мозга, преганглионарные волокна первого эфферентного нейрона © этой двухнейронной цепи относительно коротки, тогда как постгангдионарные волокна, будучи достаточно длинными, достигают места лервно-эффекторного соединения. Парасимпатические ганглии лежат вблизи иннервируемого эффекторного органа или даже в нем самом, так что в этом случае преганглионарные волокна длинные, а постгангдионарные — короткие.
Другое анатомическое различие заключается в том, в какой области центральной нервной системы находятся клетки, дающие начало преганглионарным волокнам. Мы указали выше, что симпатическая система исходит из торако-люмбального отдела спинного мозга. Парасимпатическая складывается из двух далеко отстоящих друг от друга частей — краниальной и крестцовой, и поэтому часто говорят о кранио-сакральном отделе вегетативной системы.
Функционально симпатическая и парасимпатическая системы характеризуются антагонистическими взаимо-
ГЛАВА II
/отношениями, что также служит основой для их разгра-
|ничения.
Обозначение торако-люмбального отдела как «симпатического» происходит из отчасти воображаемой, отчасти действительно существующей тенденции симпатических разрядов к генерализации, а также из их неспецифичности; при активации одного эффектора с симпатической иннервацией .все остальные также возбуждаются как бы из «симпатии» к нему. Теперь установлено, что возможны частичные симпатические разряды [332], и все же иррадиация на несколько эффекторов более типична для симпатической системы, чем для парасимпатической. Это объясняется тем, что симпатические пре-ганглионарные волокна ветвятся и оканчиваются в ганглиях, иниервирующих различные органы, тогда как постганглионарные волокна парасимпатической системы исходят из ганглиев, лежащих непосредственно в эф-фекторном органе или на нем. Поэтому по большей части одиночное преганглионарное симпатическое волокно может действовать на ряд эффекторов, а преганглионарное парасимпатическое волокно действует лишь на один из них [296].
Двойная иннервация
Раздражение нервов, идущих к различным внутренним органам, показывает, что большинство этих органов снабжено симпатической и парасимпатической иннервацией, оказывающей противоположные влияния. Раздражение парасимпатического блуждающего нерва замедляет деятельность сердца, а симпатического — ускоряет ее. Перистальтика кишечника усиливается при раздражении парасимпатических волокон и ослабляется при раздражении симпатических. Возбуждение симпатической системы вызывает сужение сосудов; те же кровеносные сосуды, которые иннервируются парасимпатическими нервами (в немногих ограниченных областях), расширяются при их активации.
Из приведенных выше примеров ясно, что ни симпатическую, ни парасимпатическую систему в целом нельзя классифицировать либо как тормозную, либо как воз-
;ПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
буждающую, но любой орган, иннервируемый обеими системами, возбуждается одной из них и тормозится другой.
Механизм антагонистических взаимоотношений между симпатической и парасимпатической системами
Противоположное действие двух отделов вегетативной нервной системы на эффекторы объясняется тем,- что химические медиаторы, выделяющиеся на месте нервпо-эффекторного контакта, различны, .Парасимпатический медиатор — ацетилхолин, а симпатический — норадрена-лин (с некоторыми исключениями). Соответственно передачу возбуждения называют холинэргической и ад-ренэргической. По большей части висцеральные эффекторы, возбуждаемые ацетилхолином, тормозятся норад-реналином, и наоборот.
Объяснение периферических влияний симпатической и парасимпатической систем не раскрывает механизма центрального распределения импульсов на эфферентные нейроны двух противоположных систем '. Это .имеет важнейшее значение при анализе причин и лечении эмоциональных и психических расстройств, в которых вегетативная система играет важную роль. Психотерапевт должен уметь воздействовать на центральную нервную систему путем .внушения и беседы. Афферентный приток частично изменяется под влиянием того, что он говорит, и тех условий, в которые он помещает больного. Это приводит к изменению характера центральной активности, к изменению поведения больного и, таким образом, к изменению информации, идущей по обратным связям от его эффекторов, и к преобразованию его взаимодействия со средой. Если, кроме того, используются химио-терапевтические средства, то возможности регуляции центральных процессов значительно расширяются, так как область химических влияний и их диапазон практи-* * * *
1 Существуют некоторые исключения из общего правила об их антагонистическом действии, но мы не будем их здесь рассматривать.
3-517
ГЛАВА II
чески неограниченны. Ниже мы уделим особое внимание анализу влияния различных веществ и других терапевтических воздействий на вегетативные функции и поведение, а также рассмотрению возможных механизмов этих влияний.
В норме изменения вегетативной активности могут быть вызваны различными способами. Повышая и понижая активность некоторых афферентных нервов, можно изменять вегетативные рефлексы, влияющие на сердце, кровеносные сосуды, кишечник и другие внутренние органы. Эти рефлексы сохраняются при глубоком наркозе и после удаления части мозга или всего мозга в целом. Они могут исходить от любых рецепторов, в том числе от расположенных во внутренних органах. Например, болевые раздражения, возникающие при растяжении кишечника или действии тепла на кожу, вызывают изменение частоты сердечных сокращений и сопровождаются рефлекторным изменением просвета кровеносных сосудов. Это в свою очередь приводит к изменению уровня артериального давления. Подобные изменения можно вызвать в эксперименте раздражением некоторых областей центральной нервной системы.
Центральные и периферические вегетативные сдвиги— неизбежные сьушики эмоциональных расстройств. Наша ближайшая задача состоит в' описании функций центров, которые регулируют поток вегетативных импульсов к внутренним органам; это позволит нам подготовить читателя к обсуждению связи между эмоциями и активностью вегетативной нервной системы.
ВЕГЕТАТИВНЫЕ ЦЕНТРЫ СПИННОГО МОЗГА
Симпатические и парасимпатические нервы находятся в состоянии постоянной активности, или тонуса, даже в отсутствие внешних раздражений. Этот тонус поддерживается активностью вегетативных центров, локализующихся главным образом в продолговатом мозге и гипоталамусе, но имеющихся и з спинном мозге. Последние активируются афферентными импульсами, идущими к ним через афферентные нейроны от рецепторов, а также от продолговатого мозга и гипоталамуса. В отсутствие
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 67
супраспинальных влияний (например, после пересечения нисходящих путей) спинномозговые вегетативные рефлексы осуществляются автономно. Так, раздражение седалищного нерва вызывает повышение частоты сердечных сокращений и сужение сосудов, особенно в области живота. Такие рефлексы осуществимы даже у спиналь-ного животного, у которого путем поперечной перерезки спинной мозг отъединен от головного. У интактного животного наблюдаются сходные, но более генерализованные и более эффективные сдвиги.
Изменение внутренней среды влияет «а вегетативную активность сшшального животного. Гипогликемия вызывает симпато-адреналовый разряд, а гиперкапния и асфиксия приводят к повышению кровяного давления [16], но чувствительность к этим раздражителям у такого животного значительно ниже, че.м в норме.
Прямое раздражение спинальных центров приводит к частичной активации симпатической системы, причем характер реакции зависит от места раздражения [332]. Например, если электроды находятся в торако-люмбаль-ной части спинного мозга, даже самое незначительное по силе раздражение вызывает резкое повышение кровяного давления. Аналогичное раздражение в области нижнего шейного отдела спинного мозга вызывает симпатическую реакцию со стороны глаза (расширение зрачка, сокращение третьего века), но не приводит к повышению кровяного давления. Это лишь один пример того, что симпатическая система не всегда действует как единое целое, так что истинное значение этой системы нельзя оценить без учета ее способности к локальному, или частичному, возбуждению. Частичные симпатические разряды также возможны при раздражении продолговатого мозга и гипоталамуса.
ВЕГЕТАТИВНЫЕ ЦЕНТРЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно важные симпатические и парасимпатические центры находятся в продолговатом мозге, где они четко локализованы. Электрическое раздражение симпатических центров продолговатого мозга у экспериментальных животных вызывает отчетливые реакции во
3*
68 ГЛАВА II
всех областях тела. Они выражаются в повышении частоты сердечных сокращений, повышении кровяного давления, максимальном расширении зрачка, сокращении третьего века, пилоэрекции, потоотделении, ослаблении перистальтики кишечника и повышении содержания сахара в крови. В основе повышения содержания сахара в крови лежит следующий механизм: симпатическая активация мозгового вещества надпочечников приводит к выделению активных гормонов, которые стимулируют процесс распада гликогена в печени. Мозговое вещество выделяет адреналин и норадреналин, о которых мы подробно скажем ниже при обсуждении физиологии эмоций.
Активность симпатических центров продолговатого мозга в равной степени повышается в ответ на рефлекторное или химическое раздражение. При вдыхании воздуха с повышенным содержанием СОг или пониженным содержанием Ог у интактного животного возникают характерные симптомы возбуждения симпатической системы. Асфиксия при пережатии трахеи вызывает мощный разряд в симпатических центрах в результате комбинированного влияния гиперкапнии и гипоксии. После высокой перерезки спинного мозга та же степень асфиксии весьма незначительно сказывается на функции органов, имеющих симпатическую иннервацию; отсюда ясно, что указанные реакции почти целиком опосредуются центрами, лежащими выше спинного мозга. Установлено, что С02 оказывает прямое раздражающее действие на симпатические центры продолговатого мозга [351]; понижение содержания 02 выражается в прямом подавлении их возбудимости. Однако, если напряжение кислорода в жидкостях организма падает очень низко, возбуждаются хеморецепторы каротидного синуса, что приводит к рефлекторной активации симпатических центров продолговатого мозга, несмотря на то, что их возбудимость в условиях гипоксии понижена.
Тщательными исследованиями установлено, что симпатический центр (пресс'орная зона) расположен в основном в ростральной части продолговатого мозга [16]. Перерезка в этой области приводит к резкому падению кровяного давления и исчезновению тонических импульсов, регистрируемых в ускоряющем нерве сердца. Де-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 69
прессорная зона расположена в каудальной части продолговатого мозга и также характеризуется тонической активностью. Расширение сосудов, вызываемое раздражением депрессорной зоны, а также усилением разрядов барорецепторов, связано не с активацией парасимпатических сосудорасширяющих нервов, а с торможением симпатического центра. Вот почему у симпатэктомиро-ванного животного сосудорасширяющие эффекты отсутствуют '.
Прямое раздражение центра замедления сердечной деятельности (тесно связанного с ядром блуждающего нерва) или его рефлекторное возбуждение приводит к падению кровяного давления, зависящему от замедления сердечного ритма. При этом активность желудочно-кишечного тракта повышается2.
Следует подчеркнуть, что центр блуждающего нерва и депрессорная зона, подобно симпатическому центру продолговатого мозга, постоянно активны, а поскольку их действия противоположны, это обеспечивает контроль и регуляцию кровяного давления. Сосудосуживающий центр способствует поддержанию нормального тонуса кровеносных сосудов посредством импульсов, идущих по симпатическим волокнам к гладкой мускулатуре их стенок. Депрессорная зона, вероятно, понижает активность сосудосуживающего центра (прессорная зона) посредством реципрокного торможения. Центр ускорения сердечной деятельности (симпатический) повышает возбудимость водителя ритма сердца, а центр, тормозящий сердце (парасимпатический), противодействует этому влиянию. Таким образом, устранение любого из этих центров приводит к снятию его тонического влияния и доминированию антагонистической системы. * * * *
1 Расширение сосудов скелетных мышц, вызываемое раздражением гипоталамуса, не зависит от целостности депрессорной зоны продолговатого мозга, а связано с активацией холинэргических симпатических нервов [900а].
2 Среди других парасимпатических реакций, вызываемых раздражением продолговатого мозга, следует упомянуть рефлекторную рвоту. Она возникает при раздражении различных афферентных нервов, а также при изменениях внутренней среды. В этой реакции участвует продолговатый мозг [80],
70 ГЛАВА II
ВЕГЕТАТИВНЫЕ ЦЕНТРЫ ГИПОТАЛАМУСА И ИХ ФУНКЦИИ
Раздражение гипоталамуса
Гипоталамус — это совсем маленький участок мозга, но вряд ли существует какой-либо другой орган, Который имел бы столь важное значение для благополучия всего организма в целом. Он лежит в основании мозга, за перекрестом зрительных нервов, над «главной железой» организма — гипофизом, с которым ои тесно связан анатомически и функционально (см. [692, 736]).
В гипоталамусе, как и в продолговатом мозге, симпатические и парасимпатические эффекты вызываются из различных участков, хотя здесь это разделение не столь четко выражено. Парасимпатические реакции возникают при раздражении передней области (непосредственно за хиазмой), а симпатические — при раздражении задней и латеральной областей. Так, электрическое раздражение заднего гипоталамуса приводит к сужению сосудов брюшной полости и повышению кровяного давления. Кроме того, оно вызывает расширение зрачка, •сокращение третьего века и пилоэрекцию, но подавляет перистальтику кишечника.
Весьма интересно, что достаточно интенсивное раздражение симпатических центров гипоталамуса может привести к повышению секреции гормонов мозгового вещества надпочечников, влияние которых складывается с действием общего разряда симпатической системы. Это отчетливо видно по реакции органа, лишенного симпатической иннервации. Из ф,нг. 2, Б видно, что слабое раздражение гипоталамуса вызывает сокращение интактио-го (/), но не денервированного (//) третьего века, незначительное повышение кровяного давления и умеренное учащение сердечного ритма. При более интенсивном раздражении (ф.иг. 2, А) не только усиливаются эти симпатические реакции, но и происходит сокращение денервированного третьего века. Поскольку возможность его возбуждения нервным путем исключена (так как оно денсрвировано), можно предположить, что возбуждающим агентом служит химическое вещество, приносимое кровью. Соответствующие эксперименты показали, что
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 71
речь идет о гормоне мозгового вещества надпочечников. Этот гормональный эффект проявляется значительно позже, чем прямой эффект возбуждения симпатического нерва. Такая задержка характерна для гормональных влияний и связана с тем, что для переноса гормонов от соответствующих желез до органов-мишеней необходимо некоторое время [316].

Фиг. 2, Нервные и гуморальные симпатич?ские влияния при раздражении заднего отдела гипоталамуса [316].
Опыты па кошке под пентоталовым наркозом н флакееднлом. Характеристика раздражений: А. 1,5 в, 71 имп/сек. продолжительность импульса 1,6 мсек, длительность раздражения 7 сек. 15. Те же условий, но продолжительность импульса 0,8 мсек. Видно, что более длительные импульсы вызывают секреторную активность мозгового вещества надпочечников, что ведет к сокращению денервированного третьего века. / — нормальное третье веко; // — де-исринрованнос третье веко; /// -— кровяное давление.
Генерализованный характер реакции, вызываемой раздражением гипоталамуса, иллюстрирует фиг. 3, на которой приведена запись активности мочевого пузыря, кишечника, денервированного третьего века и кровяного давления. Раздражение заднего гипоталамуса вызывает ослабление перистальтики кишечника и сокращение мочевого пузыря, повышение кровяного давления и секреторную активность мозгового вещества надпочечников (о чем свидетельствует сокращение денервированного третьего века). Все эти реакции совпадают с результатами раздражения эфферентных симпатических нервов, идущих к соответствующим органам. Следует отметить, что на кишечник и мочевой пузырь симпатическая
η
ГЛАВА li
система оказывает тормозящее' влияние, а на гладкие мышцы кровеносных сосудов и миокард (не регистрировалось) — возбуждающее. Данный эксперимент показывает, что гипоталамус может быть источником генерализованного симпатического разряда, подавляющего одни вегетативные эффекторы и возбуждающего другие [318].
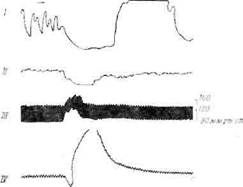
Фиг. 'J. Широко иррадирующая симпатическая и симпато-адренало-
вая реакция на слабое раздражение заднего отдела гипоталамуса
[318].
Характеристика раздражения: 1 в,'99 имп/сек, 0,8 мсек, 15 сек. / — мочевой
пузырь; // — двенадцатиперстная кишка; /// — кровяное давление; IV— де-
нервировапное третье веко.
Как упоминалось выше, парасимпатические реакции легче вызвать путем раздражения переднего гипоталамуса. Такое раздражение повышает тонус и усиливает перистальтику кишечника, снижает кровяное давление и частоту сердечных сокращений, т. е. производит действие, прямо противоположное только что описанному 1- * * * -,
1 Тормозящее влияние на мочевой пузырь зависит от секреции мозгового вещества надпочечников.
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
(при раздражении симпатического центра заднего гипоталамуса повышается кровяное давление и снижается гонус кишечника). При раздражении переднего (парасимпатического) отдела гипоталамуса происходит сокращение мочевого пузыря (органа, имеющего парасимпатическую иннервацию) и сужение зрачка.
Хотя, раздражая различные отделы гипоталамуса, можно разделить симпатические и парасимпатические эффекты, не исключена возможность смешанных реакций. Указанные зоны гипоталамуса в какой-то степени перекрываются, что особенно убедительно показал в своих экспериментах Гесс [454]. Кроме того, при раздражении заднего гипоталамуса наряду с симпатическими центрами возбуждаются волокна, идущие от расположенных рострально парасимпатических центров. В результате этого возникают комбинированные симпатические и парасимпатические реакции, а поскольку обычно они являются антагонистическими, конечный эффект для определенного органа может быть незначительным или нулевым. Существенное значение имеет не только локализация, но и характер раздражения. Например, раздражение с низкой частотой обычно вызывает парасимпатические реакции, а раздражение той же точки с высокой частотой даст симпатический эффект. Наконец, следует иметь .в виду, что периферические реакции, вызываемые раздражением гипоталамуса (например, повышение кровяного давления), нередко влекут за собой вторичные вегетативные рефлексы, которые видоизменяют первичный результат раздражения гипоталамуса.
Местная анестезия и разрушение областей гипоталамуса
Обсуждавшиеся до сих пор факты касаются фазиче-ских, или кратковременных, реакций на раздражение. А между тем гипоталамус, подобно продолговатому мозгу, оказывает тонические, или постоянные, влияния на вегетативную систему. Их легко продемонстрировать, исключив иа время или навсегда влияние части переднего или заднего гипоталамуса. Сделать это можно либо путем введения небольших количеств барбитуратов
74 ГЛАВА II
в специфические области гипоталамуса (временная блокада), либо путем очагового разрушения токами высокой частоты (стойкое разрушение).
Если задний гипоталамус действительно оказывает тонические симпатические влияния, то его разрушение или снижение его активности местным введением анестезирующего вещества должно привести к падению кровяного давления и снижению частоты сердечных сокращений. Эксперименты показали, что это действительно так. Аналогичные воздействия на область переднего гипоталамуса, исключающие или снижающие парасимпатические разряды, должны были привести к повышению кровяного давления и частоты сердечных сокращений. Это также удалось получить в эксперименте, хотя эффект был не столь демонстративен, как в опытах по исключению симпатической активности заднего гипоталамуса. Можно сделать вывод, что гипоталамус обладает тонической активностью как в отношении симпатической, так и .в отношении парасимпатической системы, но что $ норме симпатический тонус на этом уровне значительно выше парасимпатического [356, 741].
Соматические эффекты при раздражении гипоталамуса
При изучении поведения экспериментальных животных в состоянии бодрствования или поверхностного наркоза было отмечено, что раздражение гипоталамуса вызывает не только вегетативные, по и соматические реакции. Симпатические разряды, вызываемые раздражением заднего гипоталамуса, сопровождались изменениями полы - кошка выгибала спину, выпускала когти, дыхание ее заметно учащалось; она пыталась укусить экспериментатора. Весь вид кошки с расширенными зрачками, взъерошенной шерстью на спине и хвосте олицетворял ярость. Возбудимость всей соматической системы в целом была повышена.
Эта картина прямо противоположна той, которая возникает при раздражении переднего гипоталамуса. Парасимпатические реакции, такие, как сужение зрачков, падение кровяного давления и уменьшение частоты сер-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
дечных сокращений, сопровождаю юн расслаблением скелетных мышц, и животное засыпает. В других экспериментах раздражение переднего гипоталамуса вызывало мочеиспускание и дефекацию с характерными позами. Приведенные опыты показывают, что вегетативные реакции, возникающие при раздражении гипоталамуса, связаны с целым комплексом соматических — симпатические реакции сопровождаются повышением возбудимости соматической нервной системы, парасимпатические характеризуются параллельным снижением возбудимости соматической нервной системы [453].
Наконец, следует упомянуть, что изменения ритма и глубины дыхания также могут быть .вызваны раздражением гипоталамуса. Обычно повышенная распиратор-ная активность связана с симпатическими влияниями, а пониженная— с парасимпатическими [454]. Повреждение заднего гипоталамуса приводит к снижению легочной вентиляции [743].
Вегетативная функция коры
Симпатические и парасимпатические реакции могут быть вызваны раздражением коры или различных частей лимбической системы. Раздражение коры вызывает в целом более слабый и не столь генерализованный эффект, какой возникает при раздражении гипоталамуса или продолговатого мозга. Некоторые вегетативные реакции устраняются после перерезки пирамидного тракта, другие передаются через гипоталамус к низшим вегетативным центрам. Вызванные раздражением коры прессор-ные реакции связаны с сужением сосудов почек; если почка денервирована, эти реакции не возникают. Удаление моторной коры сопровождается сужением кожных сосудов противоположной стороны тела. Это показывает, что тоническая иннервация в физиологических условиях поддерживается корой. Недавно было показано, что при раздражении моторной коры (а также гипоталамуса) повышается кровоснабжение задних конечностей. Дальнейшими исследованиями установлено, что расширение наблюдается только в сосудах мышц и зависит от хо-линэргических вегетативных нервов, так как оно спи-
ГЛАВА И
мается атропином и сохраняется при перевязке спинного мозга в поясничном отделе, когда импульсы от мозга могут достигнуть этих мышц только через симпатическую цепочку [217]. Этим расширением сосудов объясняется повышение кровотока, связанное с началом произвольного сокращения мышц (а также, возможно, с тем, которое возникает, когда мы думаем о движении, не производя его в действительности).
Это явление служит примером того, что вызванные раздражением коры вегетативные разряды связаны с соматическими функциями коры. Двигательная активность коры сопровождается расширением сосудов в мышцах и повышением секреторной активности потовых желез в области ладоней (считается, что пот способствует удержанию предметов в руках) [164]. Раздражение зрительной коры сопровождается изменением диаметра зрачка, а акт жевания, связанный с раздражением коры, сопровождается слюноотделением. Напротив, состояния, сопровождающиеся возбуждением гипоталамуса или продолговатого мозга, приводят к генерализованной симпатической реакции.
ГЛАВА
НЕРВНЫЙ СУБСТРАТ ЭМОЦИЙ
Название этой главы отражает наше глубокое убеждение в том, что в основе эмоций, как ощущаемых, так и выражаемых, лежит активность нервной системы. Путем изменения активности желез и мышц в эмоциональную деятельность вовлекается весь организм, но основой эмоций все же служит центральная нервная система.
Возникает вопрос, не принимают ли какие-то определенные части центральной нервной системы более прямого участия в эмоциональных процессах по сравнению с другими, и если да, то каким образом. На протяжении всего изложения перед нами отчетливо выступает первостепенная роль гипоталамуса, это убеждает нас в том, что ему принадлежит такое же место и в механизме интеграции эмоций. Его значение объясняется не тем, что он является «седалищем» эмоций, или каким-то «эмоциональным центром», или группой таких центров. Скорее оно связано с его исключительными взаимоотношениями с остальными отделами нервной системы. Это небольшая, но сложная структура, связи и взаимодействия которой с другими отделами нервной (и эндокринной) системы имеют особое значение для состояний организма, лежащих в основе эмоций.
КОРТИКО-ГИПОТАЛАМО-ЛИМБИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
Давно известно, что выражение эмоций может осуществляться в отсутствие новой коры. Однако активность новой коры, несомненно, имеет важное значение для интеграции эмоциональных ощущений. Брэйди [90]
глАвА Ш
дал краткий обзор первых клинических и экспериментальных данных, касающихся участия коры в эмоциональных процессах. Эти данные указывают на особую роль лобных долей. Точно установлено, что повреждения лобной и префронтальной областей или их удаление снижают эмоциональную реактивность, однако довольно часто такие разрушения приводят к повышенной эмоциональной лабильности, которая характеризуется резкими колебаниями эмоционального состояния. Сдвиги, возникающие в результате участия коры, должны рассматриваться в плане интегративных отношений между лим-бической системой и диэнцефало-кортикальными взаимодействиями.
Следует подчеркнуть, что, как показали Мерфи и Гельгорн [671], раздражение гипоталамуса оказывает сильное генерализованное облегчающее влияние на активность новой коры. Такое облегчение наблюдается при разных условиях. Вызванные раздражением коры сокращения мышц и разряды в пирамидных путях усиливаются при одновременном раздражении гипоталамуса [310]. Раздражение гипоталамуса подавляет «диа-ловые» волны в электрокортикограмме, приводит к повышению частоты и амплитуды кортикальных воли, а также частоты стрихнинных пиков, вызываемых местным нанесением стрихнина на кору. Таким образом, раздражение гипоталамуса при самых разнообразных условиях приводит к повышению активности новой коры. Это возбуждение коры бывает двухсторонним, хотя оно сильнее выражено на ипсилатаральной стороне. Оно было обнаружено в моторной, сенсорных и ассоциативных зонах, а также в базальных ядрах мозга. Обширность кортикальных зон, доступных облегчающим влияниям со стороны гипоталамуса, указывает на необходимость их учета при анализе интегративного действия нервной системы, в том числе функций коры. Без всестороннего учета влияний гипоталамуса нельзя изучать механизмы, лежащие в основе деятельности и эмоций как в норме, так и в патологии.
В другой статье Мерфи и Гельгорна [672] показано, что, поскольку гипоталамус способен активировать ассоциативные области коры, он, по-видимому, имеет особое
ИПРВНЫИ СУБСТРАТ ЭМОЦИЙ
значение для эмоциональных процессов. Изменения возбудимости и уровня активности ассоциативных областей должны изменять характер ощущений, возникающих при появлении в коре импульсов, приходящих по специфическим сенсорным проекционным путям. Это не только вероятно, но просто неизбежно, так как значение «специфических» сенсорных импульсов определяется взаимодействием нервных структур, в том числе и ассоциативных областей. Эти значения в свою очередь и определяют эмоциональные ощущения.
Кроме описанного выше прямого влияния гипоталамуса на новую кору, можно указать на его значение как центра лимбической системы, которая также имеет важные функциональные связи с новой корой. Прибрам и Мак-Лин [726] исследовали взаимосвязи между лимбической и экстралимбической корой медиальной и ба-зальной областей полушарий методом стрихнинной ией-ронографии. Они выделили пять обширных областей с реципрокными связями; о наличии таких связей свидетельствовало то, что нанесение стрихнина в пределах этих областей лимбической коры вызывало разряды в экстралимбической коре, и наоборот.
Таким образом, новая кора, которая, несомненно, играет важную роль в субъективном истолковании внутренних состояний, т. е. в возникновении аффекта или ощущения, по всей вероятности, оказывается способной к осуществлению таких интерпретаций на основе своего взаимодействия с другими структурами, особенно гипоталамусом, лимбической и ретикулярной системами, а также за счет взаимодействия между различными областями самой новой коры.
В этой главе мы намерены описать главным образом анатомические связи гипоталамо-димбической системы, которая вместе с ретикулярной формацией и новой корой, как мы считаем, составляет основной механизм центрального нервного субстрата эмоций.
ГЛАВА III
ОБ ИСПОЛЬЗОВАНИИ ТЕРМИНА ЭМОЦИИ
Более четверти века назад было высказано предположение, что термин эмоция удобен для обозначения некоторых специфических форм изменения поведения, которые не поддаются объяснению, но что в интересах более точных исследований от использования этого термина следует отказаться [202, 203]. Несколько ранее было предсказано, что эмоции постепенно исчезнут из сферы психологии [643]. Теперь кажется, что это предсказание вряд ли осуществится в недалеком будущем, быть может, потому, что, как утверждал Данлоп [205], факт сушесгвования эмоций доказывается при всяком акте интроспекции. Беспокойство, подавленность, воодушевление, безразличие, гнев, страх, предвкушение приятного, опасения — все эти состояния знакомы каждому, так как все мы сами их переживали. Никто не станет отрицать, что наше внешнее поведение отражает сдвиги эмоционального состояния и, хотя наблюдаемое поведение не отражает характера или интенсивности ощущения с абсолютной точностью, оно является достаточно надежным его индикатором. Это справедливо, несмотря на гот факт, что доступное наблюдению поведение в значительной степени является результатом соматической активности, которую можно рассматривать в основном как побочный продукт глубоких внутренних сдвигов.
Вегетативные сдвиги точнее отражают эмоциональное состояние, чем реакции скелетных мышц. Всякому знакомы такие проявления эмоций, как изменение цвета лица, ускорение сердечного ритма, холодный пот и сухость во рту. Если добавить к этому все те различные показатели, которые доступны исследователю, например интенсивность секреции различных желез, кровяное давление, температура кожи, перистальтика кишечника и т. д., можно составить длинный ряд «индикаторов эмоций». Их смысл и значение не всегда легко оценить, несомненно, однако, что такие показатели имеются в избытке.
Считая, что все эти явления, т. е. эмоциональные ощущения, выражение эмоций, внешнее поведение и изменения со стороны внутренних органов, представляют
нервный субстрат эмоции
собой результат определенных видов активности центральной нервной системы, мы, естественно, пытаемся найти те ее области, в которых изменения активности кажутся наиболее достоверно связанными с эмоциями. Для этого необходимо сначала рассмотреть общее развитие нервной системы в ее отношении к различным аспектам поведения, поскольку никто не может ощущать чужих эмоций и составляет представление о них лишь на основании поведения (в том числе со слов) других людей.
РАЗВИТИЕ НЕРВНОЙ СИСТЕМЫ И ПОВЕДЕНИЕ
Яковлев [964] в своей интересной статье сводит поведение к различным формам движения, которые у всех организмов проявляются в трех сферах: 1) внутренней; 2) внешних реакциях, отражающих сдвиги внутреннего состояния; 3) внешних движениях, направленных на изменение окружающей среды. Первая из этих форм, висцеральная, согласно предположению Яковлева, наблюдается у низших организмов и на ранних стадиях эмбриогенеза более высоко организованных организмов, как единственная форма поведения; речь идет о «движении молекул и клеточном метаболизме». По мере того как в организмах возникают более сложные механизмы и развивается зачаточная, диффузная нервная система, три сферы поведения отчетливо дифференцируются на: 1) двигательную активность внутренних органов, включающую не только внутриклеточные сдвиги, связанные с обменом, ' но и дыхание, кровообращение, секрецию, экскреторную функцию и т. д.; 2) внешнее выражение внутренних состояний, или выражение эмоций; 3) двигательную активность эффекторов, создающую изменения во внешнем материальном мире, окружающем животное.
Яковлев подчеркивает, что эти сферы поведения не являются независимыми категориями, а представляют собою интегральные части поведения в целом. Он рассматривает двигательную активность внутренних органов как основу всего поведения, а се нервным субстратом считает самую «сердцевину» (core) центральной нерв-
8>:
ГЛАВА III
ной системы, которая представляет собой дуффузное сплетение короткоаксопных нейронов, образующих сеть, или ретикулум. В состав этой системы входят такие обособленные компоненты, как средний мозговой пучок, дорсальный продольный пучок Шютца и перивентрику-лярная система волокон, сетевидных по структуре и служащих для соединения воедино обонятельного переднего мозга, гипоталамуса, таламуса, центрального серого вещества и клеток, из которых идут импульсы в эффектор-ную часть симпатической и парасимпатической систем. Эта сердцевина, которая служит нервным субстратом висцеральной сферы поведения, отчетливо видна в среднем мозге человека, где нервная ось сохраняет свою исходную форму (фиг. 4,А). Центральное положение в стволе мозга всех млекопитающих занимает ретикулярная формация. На фиг. 4,5 видно, что из ретикулярной области исходят симпатические и парасимпатические волокна, идущие к внутренним органам после синаптиче-ских переключений в вегетативных ганглиях.
«Промежуточная» система Яковлева также состоит из сетевидного сплетения волокон, но для нее типичны более длинные и толстые дендриты и аксоны и более выражена тенденция к образованию обособленных ядер и пучков волокон. Эта система «интегрирует главным образом «осевые» движения, служащие для поддержания позы при внешнем выражении внутренних состояний, или эмоций». Эта система организована по принципу последовательного соединения. В ней нет кортико-спинальных нейронов, так как кора воздействует на мо-тоиейроны скелетных мышц, лежащие в стволе мозга и спинном мозге только через ряд переключений, находящихся в различных подкорковых ядрах (фиг. 4,5). Именно кора филогенетически промежуточного возраста регулирует выразительные аспекты поведения.
«Наружная» система выступает как особый компонент нервной оси только у млекопитающих, где она связана с новой корой, отсутствующей у низших животных. Эта система развивается из промежуточной системы и характеризуется наличием длинных волокон чисто кортикального происхождения, которые связываются с фондом мотонейронов двумя различными путями. Первый

Фиг. 4. Центральные эффекторпые системы [964]. Л. Поперечный разрез среднего мозга; внутренняя система (не заштриховано), промежуточная (пунктир) и внешняя (показано черным). Б. Внутренняя висцеральная система. В. Промежуточная система выражения внутренних состояний (мотонейрон скелетной мышцы, показанный внизу, получает волокна, подходящие к нему после переключения в различных подкорковых ядрах). Г. Внешняя эффекторнзя система.
ГЛАВА III
идет через кортико-бульбарные и кортико-спинальные волокна, в обход различных подкорковых ядер непосредственно к мотонейронам. Этот путь известен как пирамидная система; она представлена на фиг. 4,Г одиночным нейроном, тело которого находится в моторной коре, а аксон заканчивается на мотонейроне спинного мозга. (Хотя эту систему принято представлять указанным образом, большинство, если не все ее волокна, на самом деле оканчиваются синапсами на короткоаксон-ных вставочных нейронах, принадлежащих к внутренней, диффузной системе.) Именно эта система обеспечивает особую иннервацию скелетных мышц, которая в ходе эволюции приобрела независимость от функций поддержания позы и локомоции. Использование руки в качестве орудия зависит от целости пирамидной системы, которая обеспечивает тонкое управление пальцами. Именно человеческая рука сделала возможным преобразование окружающего материального мира.
Однако пользование руками невозможно без осуществления постуральных и локомоторных функций, регулируемых главным образом более древними подкорковыми ядрами. При этих взаимосвязанных движениях и изменениях позы важнейшая роль принадлежит второму типу связи новой коры с мотонейронами. Многие пути, идущие от коры, не подходят непосредственно к мотонейронам, а заканчиваются в тех подкорковых ядрах, которые организуют поддержание позы, локомоцию и выразительные движения у позвоночных, не относящихся к млекопитающим. Эти экстрапирамидные пути переключаются в ядрах и участвуют в деятельности мышц торса и проксимальных отделов конечностей. Таким образом, особенно у высших млекопитающих, экстрапирамидная система через промежуточные ядра обеспечивает фоновую активность для актов, филогенетически осуществляемых наиболее поздней, внешней системой. Следовательно, подкорковые ядра выступают в двойной роли и у высших животных не могут быть отнесены к одной системе. (В действительности все части центральной нервной системы настолько взаимозависимы, что попытка категорического разграничения функций или структур связана со многими неожиданностями.)
ЕРВНЫЙ СУБСТРАТ ЭМОЦИЙ
Так, Яковлев [964] предполагает, что поведение развивается в направлении от внутреннего к внешнему и что таким же образом идет развитие центральной нервной системы. Внутренняя система управляет внутренними состояниями через вегетативную регуляцию внутренних органов и является основой, из которой развивается все остальное, подобно тому как висцеральные процессы служат основой развития всего поведения. Промежуточная система, развивающаяся вслед за внутренней, представляет собой своего рода «вырост» последней, интимно с ней связана и обеспечивает поддержание позы, локо-моцию и внешнее выражение внутренних состояний. Наконец, внешняя система, которая обеспечивает влияние организма на внешнюю среду, является «выростом» второй системы и сохраняет тесную функциональную связь с ней.
Параллельно развитию поведения и его нервного субстрата в направлении «изнутри кнаружи» идет эволюция и дифференциация в направлении от каудального конца к краниальному, возрастает значение передней части тела во всех видах поведения. Параллельно эн-цефализации, сопровождающей эволюционное развитие в сфере висцеральных, выразительных и двигательных функций, идет развитие внутренней, промежуточной и внешней коры. На фиг. 4,Б видно, что внутренняя система, которая состоит из сетевидных структур спинного мозга и ствола, увенчана «энтопаллиумом», который непосредственно окружает верхний конец ствола мозга и интимно связан с гипоталамусом и ретикулярной формацией, где берут начало эфферентные висцеральные нервы. С «энтопаллиумом» тесно связана обонятельная луковица; именно поэтому наиболее древняя кора и связанные с нею структуры получили название «обонятельного мозга».
Мезопаллиум (см. фиг. 4,S) окружает энтопаллиум и соединяется с мотонейронами скелетных мышц после ряда переключений в подкорковых ядрах. Эктопаллиум образует наружный слой полушарий, отделенный от ме-зопаллиума белым веществом, состоящим из ассоциативных волокон различных областей новой коры и проекционных волокон, которые соединяют неопаллиум с под-
] л А в Л i π
корковыми ядрами и мотонейронами. Эти три области коры в общем соответствуют архи-, палео- и неопалли-уму, называемым так на основании предполагаемого филогенетического возраста.
После этого введения мы можем более подробно рассмотреть анатомические взаимоотношения «внутренней» и «промежуточной» систем, составляющих вместе то, что обычно называют ретикуло-гапоталамо-лимбичеекой системой.
АНАТОМИЧЕСКИЕ СВЯЗИ РЕТИКУЛО-ГИПОТАЛАМО-ЛИМБИЧЕСКОЙ СИСТЕМЫ
В этой книге гипоталамусу отводится центральное место. Его значение в эмоциональных ощущениях и выражениях эмоций, а также в соматическом и вегетативном аспектах поведения обусловлено его стратегическим анатомическим положением и характером связей. Мак-Лин [600] в своей удивительно интересной статье остроумно подчеркнул, что главным добавлением к нервному «шасои» (состоящему из спинного мозга и ствола) служит «водитель», выбирающий возможные направления поведения и направляющий поведение организма в целом. «Предшественник такого водителя,— пишет он,— существует в гипоталамусе и обонятельном аппарате рыб — двух структурах, которые так тесно связаны у некоторых из этих примитивных организмов, что разграничить их практически невозможно». Он ссылается на Хэррика [447] как на автора данного суждения. В этом суждении подчеркивается значение понимания взаимоотношений обонятельного мозга и гипоталамуса с остальной центральной нервной системой.
Гипоталамус получает значительную часть информации от лимбических структур переднего мозга, состоящих из древних корковых и подкорковых областей, образующих «лимб», или границу вокруг ростральной части ствола мозга у входа в полушария. Сзади он соприкасается со средним мозгом без резкой границы с ним (исключение составляют отчетливо выделяющиеся мам-милярные тела гипоталамуса). В латеральном направлении область гипоталамуса простирается за маммиляр-
НЕРВНЫЙ СУБСТРАТ ЭМОЦИЙ
ные тела в вентро-медиальную часть ретикулярной формации среднего мозга, а в медиальном направлении она переходит в центральное серое вещество вокруг сильвие-ва водопровода. Гипоталамус имеет мощные реципрок-пые связи с лимбическими структурами переднего и среднего мозга.
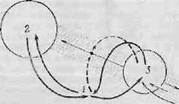
Фиг. 5. Гипоталамус (1) как узловая точка, связывающая лимби-
ческие структуры переднего мозга (2) с лимбической областью
среднего мозга (3) и ретикулярной формацией ствола (обозначено
точками) {680].
Наута [680], аналитические данные которого положены в основу предыдущего абзаца, рассматривает гипоталамус как «узловую точку обширного нервного механизма, простирающегося от медиальной стенки полушарий мозга каудалыю до нижней границы среднего мозга». Он показал это схематически (фиг. 5). Обширные реципрокные связи гипоталамуса с лимбическими областями переднего и среднего мозга представлены двумя жирными сплошными стрелками. Наута полагает, что обе цепи (гипоталамус—передний мозг и гипоталамус—средний мозг) можно рассматривать как части одной большой цепи, соединяющей лимбические области переднего и среднего мозга, причем оба.подразделения переключаются в основном в гипоталамусе. Эта система лимбических цепей, вероятно, получает большую часть своего афферентного притока через диффузные пути спинного мозга и ретикулярной формации ствола, а не через классические «специфические» афферентные пути.
ГЛАВА III
Возможно также, что импульсы, возникающие в рецепторах различных сенсорных модальностей, как висцеральных, так и соматических, поступают в лимбические цепи через диффузные восходящие пути. Известно, что различные специфические афферентные волокна дают многочисленные коллатерали к ретикулярной формации. Кроме того, существуют афферентные системы, подходящие к этой цепи от таламуса и базальных ганглиев, а также от коры, и эфферентные системы, через которые лимбические структуры могут влиять на активность таламуса и коры. Пути, по которым идут разряды от гипоталамуса и лимбических областей среднего мозга, показаны на фиг. 5 пунктирными линиями, направляющимися к ретикулярной формации ствола. Последняя передает эти сигналы как вверх, так и вниз, воздействуя таким образом на центры спинного и головного мозга.
ЛИМБИЧЕСКАЯ ДОЛЯ И РАЗВИТИЕ ПРИМИТИВНОГО МОЗГА
В сравнительном исследовании мозга млекопитающих, опубликованном в 1878 г., Брока [104] отметил стабильность кортикальных структур, граничащих со стволом мозга у входа в полушария. Он приписывает честь открытия этой «доли» Герди и Фовиллу, описавшим ее за 40 лет до него. Однако использованные ими термины вышли из употребления, а общепринятым является описательный термин Брока «большая лимбическая доля». Новая кора и лежащее под нею белое вещество, состоящее из нервных волокон, соединяющих кору с более древними корковыми и подкорковыми структурами, проходят грандиозный путь развития в ходе эволюции, но лимбические структуры остаются относительно неизменными. Однако мощное разрастание новых структур приводит к изменениям и перемещениям, так что некоторые старые связи трудно проследить. В последующих разделах мы попытаемся указать на некоторые наиболее важные из этих изменений.
Мозг развивается из передней части нервной трубки, где ее выпячивания образуют боковые желудочки полушарий. До появления этих выпячиваний передний
НЕРВНЫЙ СУБСТРАТ ЭМОЦИЙ
мозг эмбриона млекопитающего на ранней стадии развития имеет в основном цилиндрическую форму, причем
стенка состоит из массы [553]. Как показывает фиг. 6, эти массы клеток являются зачатками гип-покампа, перегородки, полосатого тела, пириформ-ной коры и неокортекса. На более поздних стадиях развития в связи с разрастанием коры и лежащих под ней волокон (в том числе массивного мозолистого тела) полосатое тело оттесняется в подкорковую область, а другие структуры первичной коры (гиппокамп, перегородка и пириформная кора) смещаются к центру. Новая кора и белое вещество полушарий буквально подавляют эти первичные корковые структуры.
Ринэицефалон, или первичный обонятельный мозг, занимает сравнительно небольшую часть полушарий мозга высших млекопитающих. Слух и зрение, которые дают гораздо более обширную информацию об окружающем мире, чем обоняние, становятся более важными для направления поведения организма в целом. Новая кора с огромным количеством нервных клеток и связей развивается
плотноупакованных клеток

Фиг. 6. Основные отделы коры головного мозга человека на разрезе [5S3].
А. У эмбриона 15 мм. Б. У эмбриона
50 мм. В. У взрослого (основная массч
гнпнокампа у взрослого сдвинута назал..
в плоскость разреза но попала).
ГЛАВА III
в основном в связи со слуховыми и зрительными дистантными рецепторами, особенно в связи с органом зрения и возникновением исполнительных механизмов, посредством которых можно влиять на зрительную информацию.
Однако первичная кора (с ее подкорковыми придатками) не исчезает и не утрачивает своих функций. Следует признать, что ее функции не вполне ясны и в какой-то части гипотетичны, но несомненно одно, что эта кора играет важную роль в регуляции висцеральных функций и эмоциональных процессов, связанных с внутренними состояниями. Мак-Лин [600] назвал эту кору «висцеральным мозгом», а последние исследования пролили свет на ее роль в организации эмоций и мотивации поведения. После описания основных анатомических связей мы рассмотрим экспериментальные данные, которые подтверждают важную роль лимбических и гипоталами-ческих структур в эмоциональных процессах.
Афферентный обонятельный путь начинается биполярными нейронами в обонятельном эпителии. Дисталь-ные концы этих нейронов прободают кутикулу эпителия и служат обонятельными рецепторами. Тела нейронов лежат среди опорных клеток обонятельного эпителия, а их короткие аксоны идут в составе обонятельного нерва через горизонтальную пластинку решетчатой кости, оканчиваясь в обонятельной луковице. Последняя.представляет собой первичный обонятельный центр [605]. У низших млекопитающих и у эмбрионов высших млекопитающих на ранних стадиях развития обонятельная луковица — это не что иное, как наиболее ростральная часть первой, или обонятельной, доли полушарий. В мозге человека она представляет собой относительно небольшое образование, лежащее на основании мозга; от этого образования берет начало обонятельный тракт, который в каудальном направлении делится на латеральную, медиальную и промежуточную полоски. Промежуточная полоска сразу же погружается в переднее продырявленное вещество (или у некоторых видов — в обонятельные бугорки). У человека она мала, непостоянна и, вероятно, не имеет большого функционального значения.
Чтобы понять анатомические связи обонятельных структур ствола мозга и новой коры, необходимо знать
НЕРВНЫЙ СУБСТРАТ ЭМОЦИИ
их эволюцию. На фиг. 7 схематически показано развитие медиальной обонятельной полоски. В мозге амфибии (тритона) обонятельная луковица — это фактически обонятельная доля, наиболее ростральная часть головного мозга. Волокна, идущие от нее каудально, по направлению к медиальным частям переднего мозга (гиппо-

Фиг. 7. Схема эволюции медиальной обонятельной полоски [553].
Схемы связей у тритона (А), крысы (Б) и человека (В). Переднее обонятельное ядро связано с передним отделом гмппокампа (/). с задним отделом гиппокампа (2) и с ядрами перегородки (3). У человека длинный рудимент гипгюкампа располагается нал мозолистым телом.
ГЛАВА III
камп и перегородка) заканчиваются синапсами на нейронах переднего обонятельного ядра.
Переднее обонятельное ядро у высших животных состоит из клеток, переместившихся из обонятельных луковиц через тракт и полоски на основание полушарий. На схемах мозга крысы и человека для простоты оно изображено компактным. Только часть волокон тракта заканчивается в этом ядре; остальные, не прерываясь, идут далее, ко вторичным центрам.
Следует отметить, что большая часть гйппокампа смещена у крысы в вентральном и каудальном направлениях. У человека перемещение гйппокампа продолжалось, пока он не занял своего положения в височной доле, оставив лишь небольшой «рудимент», над мозолистым телом (см. фиг. 7). Этот рудимент указывает направление перемещения собственно гйппокампа и поддерживает связь между ним и структурами в области перегородки. Не ясно, существует ли прямая нервная связь между обонятельной луковицей и гиппокампом [725]. Для проводимого нами анализа не существенно, имеют ли чисто обонятельные по функции волокна, идущие от обонятельных луковиц, прямые связи с отдельными рннэнцефальными структурами или подходят к ним только после синаптического переключения.
Латеральная полоска идет к структурам,, лежащим на основании полушария — к пириформной области — и миндалевидному ядру. Глур [382] рассматривает миндалевидное ядро как подкорковое скопление серого вещества, принадлежащее к обонятельному мозгу, или лим-бической системе. Среди его афферентных связей четко определены анатомически лишь обонятельные волокна, достигающие его по латеральной обонятельной полоске [382]. Поэтому мы можем рассматривать миндалевидное ядро как вторичный обонятельный центр, подобно другим структурам, к которым подходят вторичные обонятельные волокна от обонятельной луковицы и переднего обонятельного Ядра. (Это довольно широкие обобщения, но они полезны для того, чтобы получить общее представление об обонятельном мозге.)
Эллиот [220] представил простую и удобную схему, иллюстрирующую эволюцию некоторых основных обо-
НЕРВНЫЙ СУБСТРАТ ЭМОЦИИ
нятельных путей и подчеркивающую важные связи, существующие между ними и гипоталамусом (фиг. 8). Первичные обонятельные нейроны (рецепторы) связаны

Фиг. 8. Схема эволюции основных обонятельных путей [220].
Среднесагиттальный разрез мозга лягушки (А) и человека (£). Первичные и вторичные пути (тонкие линии) связывают обонятельный эпителий с обонятельной луковицей и полушарием, / — обонятельный бугорок; 2—перегородка; S — гиппокамп; 4 — гипоталамус. Из полушария пути после переключения идут по длинной дуге в гипоталамус. Перегородка и обонятельный бугорок связаны с гнппокампом более прямым путем, медиальным передне-мозговым пучком (свод).
с первичным обонятельным центром, т. е. с обонятельной луковицей. Вторичные нейроны, лежащие в луковице, непосредственно связаны с 1) обонятельным бугорком (переднее продырявленное вещество), 2) перегородкой, 3) гнппокампом и 4) гипоталамусом. (Переднее обонятельное ядро, в котором происходит переключение
ГЛАВА III
многих из этих волокон, а также волокна латеральной полоски здесь не изображены.) Мы считаем нужным особо подчеркнуть тот факт, что гипоталамус получает прямые волокна не только от обонятельной луковицы, но и от связанных с ней переключающих ядер. Один из этих постсинаптических путей, а именно путь, идущий от гиппокампа, образует дугу, располагаясь над верхним концом ствола мозга (отсюда его название свод). Другие волокна, идущие от вторичных обонятельных центров (бугорок, или переднее продырявленное вещество, и перегородка), а также волокна от первичного центра (обонятельная луковица) идут по довольно прямому пути к гипоталамусу в составе медиального передиемозго-вого пучка. Последний представляет собой древний путь, который благодаря своему положению у основания мозга почти не изменяется в ходе эволюции. Этот комплекс волокон связывает вентро-медиальные области полушарий мозга с преоптическои и гипоталамическои областями [522]. Ларселл [562] утверждает, что в его составе есть восходящие и нисходящие волокна, проходящие вдоль всей латеральной гипоталамическои области, отдающие волокна к ядрам гипотоламуса и (по данным некоторых исследователей) распространяющиеся до покрышки среднего мозга. Эти связи можно видеть на фиг. 9.
Наута [679] описывает медиальный передпЪчозговой пучок как большой древний путь, содержащий значительную часть связей, соединяющих лимбическую кору с гипоталамусом, центральным серым веществом и ретикулярной формацией среднего мозга. На фиг. 9 выделен медиальный переднемозговой пучок, но изображены только его восходящие волокна. Можно видеть, что значительное число волокон идет от ядер покрышки среднего мозга и центрального серого вещества через гипоталамус. Рострально от гипоталамуса эти волокна поворачивают к перегородке и миндалевидному ядру, которые, таким образом, очевидно, являются основными областями конвергенции афферентных обонятельных импульсов и восходящих импульсов от структур ствола мозга. Фиг. 9 представляет собой полезное схематическое обобщенное изображение основных элементов лим-

Фиг. 9. Схема лимбической доли [604].
Архикортекс показан штриховкой, а палеокортекс—пунктиром. Выделен медиальный пе-реднемозговой пучок I — основная связь между лимбической долей, гипоталамусом {2) и лимбической областью (3), которая состоит из центрального серого вещества и околоцевт-ральной ретикулярной формации среднего мозга и осуществляет жизненно важную связь между лимбической корой, нижней частью ствола мозга и спинным мозгом; 4 — переднее ядро таламуса; 0 — центральное серое вещество среднего мозга, в — вентральное и дорсальное ядра покрышки; 7 — маммиляр-ное тело; 8 — гипофиз.
ГЛАВА III
бпческой системы. Примитивная древняя кора (архикортекс), особенно гиппокамп и его рудимент (связанный с поясным пучком), окружена филогенетически более поздней старой корой (палеокортекс), состоящей в основном из поясной и гиппокамповой извилин. Архикортекс и палеокортекс с их афферентными и афферентными связями, функционируя вместе с гипоталамусом и ретикулярной формацией ствола мозга, регулируют висцеральную активность организма и обеспечивают «внешнее выражение внутренних состояний». Однако эмоции в нашем понимании, в том числе аффект и его выражение, требуют для своего осуществления взаимодействия между этими структурами и новой корой (неокортекс). Только последняя с ее пластичностью, способностью сопоставлять информацию различных' видов, хранить информацию и сравнивать явления, происходящие в данный момент, с прошлым опытом,— только такая структура может обеспечить осмысленный опыт и сознание. Таким образом, необходимо учитывать анатомические связи, которые, очевидно, представляют собою цепи, сформировавшиеся специально для обеспечения указанных взаимодействий гипоталамуса, лимбической системы и новой коры.
Круг папецА
НЕРВНЫЙ СУБСТРАТ ЭМОЦИИ рентной связью коры (архикортекса) с гипоталамусом. Позднее с появлением…РАЗРУШЕНИЯ, ДЕЛАЮЩИЕ ЖИВОТНЫХ БОЛЕЕ РУЧНЫМИ
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИЙ 111 они вновь и вновь обнюхивали их, брали в рот, бросали как несъедобные или… Результат двухстороннего удаления миндалевидного ядра и лежащей над ним коры у кошек удивительно сходен с тем, что…Центральные механизмы, участвующие в регуляции кровяного давления барорецепторами
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 157 щению артериол и .повышению частоты сердечных сокращений. Это сосудосуживающий… Наоборот, депрессорные центры имеют тенденцию снижать артериальное давление путем замедления сердечного ритма…Вегетативное равновесие и реципрокная настройка
ГЛАВА V каким образом это происходит. Эксперименты, которые мы сейчас опишем,… возбудимость парасимпатической системы при симпатической настройкеЭксперименты с гипертензивныма веществами
Ранее 'было описано, что .повышение кровяного давления вызывает усиление барорецепторяых рефлексов, влияющих «а продолговатый мозг и гипоталамус и… Специфичность этих эффектов видна из следующих фактов. Разрушение заднего… АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 179Проба с мехолилом как показатель реактивности симпатической системы у человека
I. Лица, у которых мехолил вызываем незначительное (несколько миллиметров) кратковременное снижение давления с быстрым восстановлением исходного… II. Лица, у которых кровяное давление падает на 15—25 мм рт. ст. и… III. Лица с еще более резким падением кровяного давления, которое не возвращается к исходному уровню и через 15 мин. …Г Л Л В Л IX
Поскольку мы считаем, что эмоции вызывают возбуждение гипоталамической системы, можно ожидать, что состояние тревоги должно облегчать приобретение… Участие обонятельного мозга В процессе выработки условных рефлексов участвует также древний обонятельный мозг, электрическим раздражением которого…Предневротические состояния
■СПЕРИМЕНТАЛЬНЫЕ НЕВРОЗЫ жению в0 время реакции на условный сигнал. Условная реакция исчезала на… Эти работы показывают, что под влиянием боли, эмоциональных конфликтов и других подобных условий возникают симптомы,…Поведенческие реакции и секреция АКТГ
ГЛАВА XII ден.ия были сделаны на обезьянах, у которых отмечалось беспокойство,… Механизм секреции АКТГI !
СТРЕСС И ЭМОЦИИ
ми свидетельствующими о том, что при экспериментальном несахарном диабете нарушена секреция не только антидиуретического гормона, но и АКЛТ [610].
Эти исследования могут служить убедительным доказательством участия гипоталамуса в эмоциональном возбуждении и наличия в этих условиях глубоких и сложных изменений в эндокринной системе. За последние годы эти работы пополнились прямыми доказательствами увеличения активности коры надпочечников — повышением концентрации кортикостероидов в крови.
СЕКРЕЦИЯ АДРЕНОКОРТИКОСТЕРОИДОВ И ЭМОЦИИ.
Эксперименты на животных
Лишшак и Эидрочи [585] определяли концентрацию адренокортикостероидов в крови .вены надпочечника у кошек. Они обратили внимание на то, что у одних животных секреторная активность коры надпочечников была в 8—10 раз выше, чем у других. Эти животные отличались выраженной агрессивностью и неустойчивостью поведения; достаточно было весьма слабого раздражителя, чтобы вызвать у них реакцию ярости. Далее было установлено, что у собак, у которых предварительно выработанный условный слюноотделительный рефлекс затормаживался болевым раздражением, длительность этого торможения соответствовала интенсивности секреции адренокортикостероидов. В течение этого периода (длительностью 1 —14 дней) у собак наблюдались невротические явления (тремор, беспокойство). Эти наблюдения ясно показывают, что интенсивность секреции адренокортикостероидов может служить показателем эмоционального состояния и степени активности гипоталамуса и что эмоциональные расстройства также связаны с этой секрецией.
Опыты на обезьянах, в которых об интенсивности секреции адренокортикостероидов судили непосредственно по концентрации 17-оксикортикостероидов в плазме, показали исключительную чувствительность этого индикатора эмоциональной реактивности [632]. Перевод животного из клетки в экспериментальный станок, где его
11-517
-■ , I
:S22
ГЛАВА XII
движения были несколько ограничены, вызывал значительное повышение содержания гормона в плазме. Однако при >повторении экспериментов быстро наступала адаптация и степень повышения уровня гормона уменьшалась параллельно с ослаблением эмоциональной реакции. В этих опытах были установлены следующие .интересные факты: 1) секреция адренокорти-костсроидов выше у тех животных, которые имели возможность наблюдать из клетки за другими животными, чем у лишенных этой возможности; 2) выведение гормонов с мочой уменьшалось в субботу и воскресенье, когда при том же режиме кормления обстановка в помещении, где находились животные, была менее беспокойной; 3) уровень адренокортикостероидов плазмы был выше у обезьян, живущих совместно с другими особями, чем у содержащихся в индивидуальных клетках. Сходные результаты наблюдались в опытах на мышах [143], в которых о секреторной активности коры надпочечников судили по шир.ине .пучковой и сетчатой зон. Гиперплазия этих зон возрастала с увеличением плот-кости популяции животных одного пола. (Интересно, что эмоциональная реакция на присутствие других особей не захватывает всего гипоталамуса. Напротив, го-надотропные функции в условиях сообщества ослабляются.).
Большой интерес представляет вопрос о влиянии выработки условных рефлексов на секрецию адренокортикостероидов [634]. Были поставлены две серии опытов, в которых использовалась реакция страха при включении электрического тока. В первой серии у животного вырабатывали реакцию нажатия на рычаг при пищевом подкреплении. Затем вырабатывали условную эмоциональную реакцию в ответ на щелчки; это осуществляли повторными нанесениями электрического удара в конце 5-минутного действия щелчков. Поведенческим выражением эмоции было частое мочеиспускание и дефекация, пилоэрекция, дрожание, усиленная двигательная активность и прекращение реакции нажатия на рычаг. Во второй серии вырабатывали реакцию избегания электрического удара путем нажатия на рычаг. Затем интервалы между нажатием на рычаг и включением тока сокраща-
CTI-ECC И ЭМОЦИИ 323
принуждая животное тем самым более часто нажи-Л ть на рычаг, чтобы избежать удара. По мере уменьшения интервалов стресс усиливался, и параллельно увеличению частоты нажатия на рычаг повышалась сек-пеиия адренокортикостероидов. Если в первой серии опытов электрический удар был неизбежен, а эмоциональная реакция явной, то во второй внешнее поведение животных не изменилось, а между тем секреторная активность коры надпочечников изменилась аналогичным образом. Ясно, что внешнее выражение эмоционального возбуждения не является надежным индикатором возбуждения гипоталамуса.
Ранее упоминалось, что реакция коры надпочечников на новую ситуацию быстро угасает. Эта же реакция в экспериментах с условными рефлексами удерживается в течение многих месяцев. Хотя повышение секреции АКТГ, вызываемое с гипоталамуса, имеет место в обоих случаях, степень и, возможно, механизм гипоталамиче-ской активации различен.
Секреция адренокортикостероидов и эмоции у человека
Ссылаясь на ряд косвенных данных, таких, как эо-зинопения и повышение содержания адренокортикостероидов в моче, многие авторы [473, 474, 477, 478, 4801 отмечают, что и у человека эмоции связаны с активацией гипоталамуса и секрецией АКТГ передней долей гипофиза. Телесные повреждения (ожоги) или ожидание операции оказывают такое же влияние. Что касается двигательной активности, то оказалось, что не сама по себе физическая нагрузка, а лишь сочетание с условиями эмоционального возбуждения вызывает v человека адре-нокортикальную реакцию [282, 464, 475]. Дальнейшие исследования показали, что фрустрация сопровождается повышением содержания 17-кетостероидов, степень которого тем больше, чем более .выражено расхождение между надеждами и их реализацией [57, 441]. Возраст и эмоциональность также влияют на секрецию кортикосте-поидов при стрессе [464]. Интересно, что даже оживленная беседа больного с психиатром сопровождается
ГЛАВА XII
отчетливым повышением секреции адренокортикостерои-дов [456, 464]. Аналогичные результаты наблюдались у здоровых испытуемых перед тяжелыми экспериментами с лишением сна. Соответствующие изменения отмечались у членов экипажа военного самолета перед выполнением сложного задания [632]. В течение недели перед длительным беспосадочным полетом члены экипажа жили вместе тремя отдельными группами. Интересно, что содержание адренокортикостероидов в моче было совершенно одинаковым у членов каждой группы; причем между группами наблюдались резкие различия. Этим подтверждается важная роль общения с другими людьми в определении степени эмоциональной реактивности и интенсивности гипоталамо-гипофизарных разрядов.
В недавно проведенных тщательных исследованиях Перски и др. [705] обнаружили прямую зависимость между степенью выраженности эмоционального ответа и содержанием гидрокортизона в крови. Не вдаваясь в подробности, укажем, что содержание АКТГ или близких ему веществ в крови повышалось при состоянии тревоги [706]. Было показано, что различные другие эмоции (предвкушение, гнев, подавленность и даже эйфория) также вызывали эти реакции.
Ряд моментов требует особых разъяснений. Было показано, что такие различные воздействия, как серьезные телесные повреждения (например, сильные ожоги), с одной стороны, и доверительная беседа, с другой, оказывают сходное влияние на организм; и то и другое влияет на гипоталамо-гипофизарную систему; и то и другое повышает секрецию АКТГ. Опыты Масона [633], в которых сравнивалось влияние искусственно введенного АКТГ с действием АКТГ, выделяющегося в экспериментах по выработке условных рефлексов, показали, например, что звук, сигнализирующий опасность, способен вызывать максимальную секрецию АКТГ. В свете этих данных возникает вопрос, можно ли рассматривать эмоциональные реакции, возникающие в различных ситуациях у человека и в опытах по выработке условных рефлексов у животных, как стресс, т. е. как условие, при
рЕСС И ЭМОЦИИ
:з2й
котором на гипоталамус действуют сверхсильные разд-пажители. На этот вопрос нужно ответить утвердительно При эмоциональных пробах раздражитель вызывает активацию гипоталамуса посредством кортикофугальных импульсов. Эксперименты подтверждают существование положительной обратной связи между гипоталамусом и корой. Можно думать, что это и есть механизм, ответственный за мощное влияние эмоционального возбуждения на нисходящий разряд гипоталамуса. Хотя эта концепция развивалась на основе изучения взаимоотношений неокортекса и гипоталамуса [309]. ее уместно распространить и на лимбическую кору [680], влияние которой на эмоциональную реактивность гипоталамуса обсуждалось в этой главе.
Результаты различных исследований, описанных нами выше, позволяют сделать вывод, что в условиях, когда возбудимость гипоталамуса варьирует в широких пределах, гипоталамо-кортикальный разряд происходит параллельно с нисходящим разрядом. Эти исследования подтверждаются описанными в этом разделе наблюдениями, а именно уменьшением эмоциональной и адре-нокортикальной реакции в процессе угашения. То же наблюдается и при действии фармакологических препаратов. Под действием резерпина у обезьян одновременно уменьшились условнооефлекторная реакция беспокойства и секреция [633]. В работе Перски и сотр. [705, 706] показано, что и у человека высвобождение АКТГ коррелирует с интенсивностью эмоционального нарушения. Следовательно, предложенная интерпретация приложима и к человеку.
Взаимоотношения гипоталамуса и коры надпочечников в условиях стресса вообще и эмоционального стресса в частности рассмотрены более подробно, так как эта область изучена лучше, чем вопросы влияния стресса на другие тройные гормоны передней доли гипофиза. Необходимо, однако, подчеркнуть, что влияние эмоций на гипоталамо-гипофизарную систему носит более сложный характер, чем это можно себе представить на основании опытов с АКТГ. Селье еще в одной из своих первых работ утверждал, что стресс-состояние ведет к подавлению Функции половых желез. Поскольку- исследованиями
ГЛАВА XII
последних лет убедительно показано, что функциональная активность и анатомическая целостность половых желез зависят от импульсов из гипоталамуса [169], снижение интенсивности и блокада сперматогенеза после тяжелой психической травмы [872] подтверждает, что в этих условиях имеет место торможение определенных отделов гипоталамуса. Однако механизм этого явления пока неизвестен.
Возможно, что дальнейщие результаты остроумных опытов Рихтера [754] внесут большую ясность в эту проблему. Рихтер .показал, что длительное плаванье вызывает у крыс глубокие нарушения двигательной активности, веса, а также функции яичников. У нормальных крыс наблюдается 4—5-дневный цикл спонтанной двигательной активности, совпадающий с фазами активности яичников. Максимум обоих видов активности приходится на время течки, когда наблюдается ороговение слизистой влагалища. После истощающего плавания наблюдаются характерные изменения: животное начинает потреблять значительно больше пищи и жиреет, как при разрушении определенных отделов гипоталамуса, В это же время уменьшается двигательная активность и циклическая активность яичников и возникает своего рода «мнимая беременность» продолжительностью 11— 17 дней. Гиперфагия, нарушение овариального цикла одновременно с гипертрофией надпочечников и атрофией лимфатических узлов (обнаруживаемой при аутопсии) позволяют предположить, что состояние тяжелого стресса прежде всего сказывается на гипоталамусе. Такое предположение подтверждается тем, что вслед за острым стрессом наступает каталепсия и полидипсия — состояния, обусловленные разрушением заднего и переднего отделов гипоталамуса соответственно.
Аналогичные явления можно вызвать у диких крыс путем кратковременного воздействия яркого света, который вызывает, как известно, усиленную активность гипоталамуса. Однако стресс, вызываемый воздействием «внешних» факторов, не является единственным состоянием, при котором наблюдается усиление активности гипоталамуса. Секреция «тройных» гормонов передней долей гипофиза регулируется не только импульсами из
стресс и эмоции
гипоталамуса, но и содержанием в крови гормонов соответствующих желез внутренней секреции. Уменьшение концентрации того или иного гормона стимулирует секрецию, возможно через посредство гипоталамуса. Поэтому, если в результате удаления щитовидной железы или по какой-нибудь другой причине концентрация ти-реоидных гормонов в крови снижается, то тот отдел гипоталамуса, который ответствен за регуляцию секреции тиреотропного гормона, переходит в состояние напряженного функционирования. Рихтер показал, что при этих условиях у крыс возникают такие же нарушения овариального цикла, как и в условиях пробы с истощающим, плаванием.
Эти исследования значительно расширили наши представления о механизмах действия стресса на организм. Причем внешние и внутренние стресс-факторы в этом отношении в равной мере эффективны.
Изменения, вызванные снижением концентрации ти-реоидного гормона в крови, не только стимулируют секрецию тиреотропного гормона, но и вызывают изменения веса тела, двигательной активности и функции половых желез. Очевидно, в этих условиях изменяется активность гипоталамуса в целом; нередко после острого стресса наблюдаются патологические сдвиги в поведении.
ЗАКЛЮЧЕНИЕ
■В четвертой части мы обсуждали три различных явления: экспериментальные неврозы, влияние ограничения ceHcqpHoro притока и влияние стресса. Несмотря на кажущиеся различия, они имеют по крайней мере два общих свойства. Они находятся «а границе между физиологией и патологией и характеризуются изменением активности гипоталамуса и эмоциональным возбуждением. В зависимости от интенсивности и длительности раздражений, вызывающих экспериментальный невроз, нарушения поведения могут быть либо незначительными и кратковременными, либо серьезными и стойкими. В последнем случае даже у низкоорганизованных животных нарушается индивидуальное и социальное поведение и
ГЛАВА XIJ
возникают состояния, подобные психическим заболеваниям у человека. Об этом неоднократно писал Павлов. Ограничение сенсорного .притока даже на несколько часов вызывает психические нарушения типа галлюцинаций. Эмоциональный стресс вызывает усиленную секрецию адренокортикостероидов; эксперименты Вудбери [960] и многочисленные клинические случаи осложнений при применении кортикостероидов .в терапевтических целях свидетельствуют об исключительной чувствительности мозга к этим гормонам. Необходимо добавить, что стероиды, выделяемые половыми железами, оказывают глубокое влияние на поведение. Это можно обнаружить при изучении материнского инстинкта и инстинкта гнездования. Недавно было показано, что стероиды изменяют возбудимость гипоталамуса и лимбической системы [524, 525]. Это позволяет считать, что в основе эмоциональных расстройств могут лежать и изменения внутренней среды. Однако исследования Хогленда [473, 475, 478], пытавшегося обнаружить качественные или количественные различия в секреции адренокортикостероидов при психических расстройствах, не дали результата.
Наш анализ подтверждает, что в основе симптомов экспериментального невроза лежит нарушение равновесия систем гипоталамуса, возникающее при интенсивном раздражении. Исследование стресса показывает, что изменение активности гипоталамо-гипофизарной системы в условиях сильного эмоционального возбуждения может играть важную роль, изменяя реактивность мозга в целом и гипоталамическои системы в частности. Следует напомнить, что по данным Рихтера .влияние чрезмерного эмоционального возбуждения не ограничивается корой надпочечников, а распространяется и на другие эндокринные- железы (щитовидную, половые).
Тенденция к сильным эмоциональным, «взрывам», наблюдающаяся при переводе животных, содержавшихся в условиях ограничения сенсорного притока, в нормальную обстановку, требует дальнейших разъяснений. Поскольку подобное поведение обычно наблюдается в случаях снижения корковой регуляции (аноксия, опьянение, наркоз), одним из факторов, ответственных за наруше-
рЕСС И ЭМОЦИИ
ние поведения в состоянии ограниченного сенсорного притока, может быть отсутствие или снижение кортико-гипоталамических импульсов. Координированная функция этих путей обеспечивает интеграцию соматических и вегетативных процессов при раздражении, а также при физиологической активации тех или иных зон коры. В таких условиях симпатическая система не функционирует как целое, а обнаруживает ограниченную активность адаптивного характера. Раздражения, поступающие из окружающей среды, видимо, играют первостепенную роль в развитии кортико-гипоталамических функциональных связей, лежащих в основе иптегративных актов, равно как и созревания гипоталамо-гипофизарнои системы. Необходимо, однако, допустить, что нарушения эмоций и восприятий, наблюдаемые у человека после нескольких часов ограничения сенсорного притока, отличаются по своему происхождению от симптомов, наблюдавшихся у экспериментальных животных при ограничении сенсорного притока в течение многих месяцев и лет.
ЧАСТЬ ПЯТАЯ
ЭМОЦИИ, НАРУШЕНИЕ РАВНОВЕСИЯ СИСТЕМ ГИПОТАЛАМУСА И СОМАТИЧЕСКИЕ РАССТРОЙСТВА
ГЛАВА XIII
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ГИПЕРТОНИИ
РОЛЬ БУФЕРНЫХ НЕРВОВ
Барорецепторы сино-аортальной зоны оказывают тоническое тормозное действие на симпатические центры через свои афферентные «буферные» нервы. Следовательно, при денервации этой области симпатические центры освобождаются от тормозного влияния и, подвергаясь возбуждению, вызывают повышение кровяного давления.
Для выяснения роли нейрогенного фактора в патогенезе гипертонической болезни проводилось тщательное изучение .повышения кровяного давления в остром опыте [458]. С этой же целью проводились наблюдения над животными с перерезанными буферными нервами в условиях хронического опыта. Существуют, однако, серьезные основания к тому, чтобы поставить под сомнение вопрос о приложимости этих данных к проблеме гипертонии у человека. В этих опытах гипертония была нестойкой; через различные промежутки времени уровень кровяного давления возвращался к норме. Не исключена возможность, что это является результатом неполной денервации сино-аортальной зоны в хроническом опыте. Следует иметь в виду, что у лабораторных животных (за исключением кролика) нервные волокна, проводящие импульсы от барорецепторов аорты к центральной нервной системе, идут в составе ваго-симпатических стволов. При попытке денервации сино-аортальной зоны экспериментатор может либо разрушить эфферентные волокна системы блуждающего нерва, что влечет за собой смерть от аспирационной пневмонии, либо, не полностью денервировав аорту, создать условия, при которых восстановление нормального кровяного давления
ГЛАВА XIII
осуществляется за счет сохранившихся депрессорных (буферных) нервов аорты. Более того, следует напомнить, что даже полная денервация сино-аортальной зоны не исключает влияния буферных нервов, так как ба-рорецепторы существуют в области легких и брыжейки [412, 457]. (Недавно было опубликовано сообщение о развитии стойкой гипертонии при двухстороннем сужении области каротидного синуса, хотя каротидный нерв оставался интактным [921]. Однако Кэзди [531а] не удалось подтвердить эти результаты.)
КЛИНИЧЕСКАЯ И ЭКСПЕРИМЕНТАЛЬНАЯ ГИПЕРТОНИЯ
Было показано, что изменения со стороны сердечнососудистой системы при экспериментальной «нейроген-ной» гипертонии отличаются от тех изменений, которые наблюдаются в клинике. После денервации сино-аортальной зоны кровяное давление становится лабильным, заметно ускоряется сердечный ритм, возрастает минутный объем сердца; при клинической гипертонии (а также при экспериментальной почечной гипертонии) минутный объем не меняется, а .возрастает сопротивление периферических сосудов току крови.
Очевидно, что клиническая, так называемая эссенци-альная, гипертония отличается от экспериментальной нейрогенной гипертонии и скорее напоминает экспериментальную почечную гипертонию. Это, однако, не исключает важной роли барорецепторных механизмов в клинической гипертонии. Нужно добавить, что облегчение симпатического разряда при перерезке буферных нервов оказывает прямое влияние на почечный кровоток. Такая перерезка вызывает гипертонию у собак даже после симпатэктомии, но при условии, что симпатические нервы почек и надпочечников не повреждены. Если же у такого симпатэктомированного животного перерезать и почечные нервы, уровень кровяного давления восстанавливается [408]. На основе наблюдавшегося в эксперименте выраженного сужения сосудов почек после сино-аортальной денервации можно заключить, что снижение активности барорецепторов каротидного синуса
нейрофизиологические механизмы гипертонии 335
сопровождается ишемией почек, ведущей к почечной гипертонии.
Хотя при анализе нейрогенной гипертонии мы не рассматривали функциональных сдвигов, характерных для почечной гипертонии, ряд опытов из этой области исследования имеет отношение к нашей проблеме — функционирование барорецепторов. Важно.знать количественные характеристики их активности в условиях гипертонии независимо от вызвавшего ее механизма. Регистрация потенциалов действия каротидного нерва при ритмичных колебаниях давления в сонной артерии, имитирующих нормальную пульсацию, выявила существенные различия между контрольными собаками и собаками с гипертонией. Как показано на фиг. 34, предельная величина кровяного давления, при которой начинают функционировать барорецепторы, в состоянии гипертонии смещена кверху по сравнению с контрольными животными. У животного с гипертонией при давлении 60 мм рт. ст. активность барорецепторов не регистрируется, тогда как у контрольного животного при той же величине давления наблюдаются отчетливые разряды барорецепторов. Видно также, что при 240 мм рт. ст. активность барорецепторов у контрольных собак носит непрерывный характер, тогда как у животных с гипертонией она ограничена только периодом систолы. Поскольку частота разрядов и число активных элементов по мере повышения давления внутри синуса возрастают, сдвиг порогового давления и уменьшение активности при данной величине давления (например, 120 мм рт. ст. на фиг. 34) ясно показывают, что гипертония связана с пониженной реактивностью барорецепторного механизма. Порог раздражения барорецепторов увеличивается, но они продолжают функционировать, особенно при высоких показателях кровяного давления, которые характерны для гипертонии [533, 613].
Мы попытаемся в дальнейшем оценить роль барорецепторов в патогенезе гипертонии у человека, но сначала необходимо обсудить другие нейрогенные механизмы, которые могут играть роль в возникновении гипертонии. Если высвобождение симпатической системы от сдерживающего влияния буферных нервов, идущих от сино-
Фиг. 34. Иейрограммы каротидного синуса у «>бак с нормальным (Л) и повышенным (б) давлением [613].
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ГИПЕРТОНИИ 337
аортальной зоны, нельзя считать достаточно убедительным объяснением механизма гипертензии у человека, то необходимо, по-видимому, исследовать, не могут ли играть определенной роли в этом заболевании другие формы возбуждения симпатических центров. Такое исследование оправдано с точки зрения повседневного опыта, свидетельствующего о значительном влиянии эмоционального возбуждения на сердечно-сосудистую систему. Существует ряд физиологических данных, подтверждающих предположение о том, что эмоциональное возбуждение может вызвать довольно стойкое повышение кровяного давления с соответствующими патологическими изменениями в различных тканях организма.
Аудаогенная гипертония
Отмечали, что длительное воздействие звуковых раздражителей приводит к гипертонии; правда, ряду авторов не удалось воспроизвести эти результаты. Ротлин и др. [771] показали, что успех эксперимента зависит не только от соответствующего раздражителя, но и от чувствительности животного. Так, у инбредных крыс звуковое раздражение не вызывает столь ■выраженного повышения давления, как у гибридных (самец — дикая норвежская крыса, самка — лабораторная крыса). Хотя возникающая при этом гипертония постепенно исчезает, она все же длится более двух месяцев после прекращения раздражения —довольно большой отрезок жизни крысы. Генетические факторы, очевидно, также оказывают существенное влияние на эмоциональную реактивность. О роли этих факторов свидетельствуют опыты с пробой на сосудосуживающее действие холода (повышение кровяного давления при опускании руки в ледяную воду). Повышенные сосудистые (симпатические) реакции наблюдаются не только у больных гипертонией, но и у их детей. У 95% людей, родители которых страдают гипертонией или дают повышенную сосудистую реакцию (по сравнению с контролем), отмечается положительная реакция на холодовой тест. Кроме того, при гипертонии У обоих родителей в 90% случаях развивается гипертония у детей [468]. Вместе с тем описан случай развития
ГЛАВА XIII
гипертонии только у одного из лары однояйцевых близнецов; очевидно, это объясняется более сильным эмоциональным стрессом, выпавшим на долю одного из генетически идентичных близнецов [955].
Сначала звуковое раздражение вызывает у крыс явные двигательные проявления сильного страха; но спустя 1—2 недели этот соматический симптом исчезает. Вегетативные же реакции сохраняются и через несколько месяцев приводят к гипертонии. Очевидно, в этом случае, так же как и при экспериментальных неврозах, возникшие под влиянием эмоций гипоталамические реакции сохраняются после исчезновения внешних проявлений эмоций. Важно подчеркнуть, что после, казалось бы, полного исчезновения аудиогенной гипертонии повторное звуковое раздражение оказывается более эффективным — гипертония развивается быстрее, чем в первый раз. Предварительное введение животным препаратов спорыньи (гидергин), подавляющих активность симпатических центров и устраняющих периферическое действие адреналина, значительно снижает эффективность гипертензивного действия звукового раздражения. Хотя этих данных недостаточно для того, чтобы доказать возможность развития стойкой гипертонии в результате сильного эмоционального возбуждения, они тем не менее свидетельствуют об определенном влиянии этого фактора на кровяное давление. Результаты этих работ можно сопоставить с клиническими наблюдениями, касающимися возникновения длительной, но не постоянной гипертонии под влиянием сильных эмоций.
ПОВЫШЕННАЯ РЕАКТИВНОСТЬ СИМПАТИЧЕСКОЙ СИСТЕМЫ И ГИПЕРТОНИЯ
Шанк[811] использовал в своих опытах естественное эмоциональное возбуждение: он помещал кошек на несколько месяцев вблизи лающих собак. У 50% животных возникла гипертония, причем у некоторых наблюдалось, кроме того, увеличение сердца с гипертрофией левого желудочка. Эти исследования подтверждают предположение, что усиленное возбуждение симпатического
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ГИПЕРТОНИИ 339
отдела гипоталамуса может играть важную роль в патогенезе гипертонии. Наблюдавшиеся у этих кошек гистологические изменения в передней доле гипофиза (ба-зофилия) свидетельствуют о повышении гипоталамо-гипофизарной активности. Кроме того, отмечалась гипертрофия коры надпочечников (расширение пучковой зоны). Эти эксперименты необходимо повторить, чтобы доказать воспроизводимость этих результатов. Интересно, что при таком же эмоциональном возбуждении (присутствие лающих собак) у кошек отмечали ишемию почек [68]. Введенная внутривенно китайская тушь обычно равномерно распределяется в корковом слое почек; у кошек, бурно реагировавших на встречу с собакой, тушь в корковом слое почек либо вовсе отсутствовала, либо ее было значительно меньше. Не исключена возможность, что сужение сосудов под влиянием эмоционального возбуждения может привести к возникновению анатомических и гуморальных сдвигоз, характерных для почечной гипертонии [384].
Ранее упоминалось, что экспериментальный невроз возникает .при конфликтной ситуации, например если на фоне осуществления условного пищевого рефлекса применяется электрический удар, вызывающий защитную реакцию. Применение этого и сходных методов на обезьянах приводит к развитию гипертонии, которой предшествует гипотензивная фаза ([605], см. также [709]). К сожалению, мы не знаем, наблюдалась ли в этих случаях постоянная гипертония, но интересно отметить возникновение коронарной недостаточности, приводящей к инфаркту. Другой успешный способ создания экспериментальной гипертонии — это изоляция обезьяны от остальной группы, которая продолжала находиться в ее поле зрения. Особенно сильное эмоциональное возбуждение наблюдалось лри кормлении остальной группы на глазах у изолированного животного.
На основании этих и других экспериментов [949] можно сделать вывод, что: 1) различные виды эмоционального напряжения вызывают временное или постоянное повышение кровяного давления, которое может сопровождаться глубокими изменениями со стороны сердечно-сосудистой системы; 2) важную роль в этом про-
ГЛАВА ХШ
цессе играет не только интенсивность и частота эмоционального возбуждения, но и индивидуальная чувствительность животного, которая в значительной мере определяется генетической конституцией [771, 873]. Так, известно, что у некоторых пород собак, отличающихся особой возбудимостью, возникает длительная гипертония после денервации сино-аортальной зоны; у других собак этого не наблюдается [810].
Клинические работы, и в том числе работа Вольфа и др. [954], подтверждают эти выводы. Было замечено, что психическое напряжение при беседе с психиатром вызывает у больных гипертонией более выраженные функциональные сдвиги, чем у здоровых людей. Хотя при гипертонии наблюдаются некоторые характерные особенности психики — навязчивые состояния, неуверенность [65, 406, 809], один и тот же человек может реагировать на такую беседу в разное время по-разному. В случаях беспокойства или необходимости подавить свои эмоции повышение кровяного давления связано с увеличением ударного объема; периферическое сопротивление сосудов при этом остается неизменным. Вместе с тем при таких эмоциях, как негодование и гнев, а также при явных признаках общих эмоциональных расстройств кровяное давление повышается главным образом за счет увеличения периферического сопротивления; ударный объем при этом остается без изменений.
Установлено, что в зависимости от характера эмоций пульсовое давление может либо возрастать, либо уменьшаться, а систолическое давление в обоих случаях возрастает [904]. При этом, естественно, возникает вопрос, каким образом один и тот же конечный результат —повышение кровяного давления — достигается при разных эмоциональных состояниях с помощью различных механизмов. Ответом отчасти может служить тот факт, что хотя при обоих видах эмоций наблюдается повышение симпатической активности, тем не менее при состоянии негодования возрастает выведение с мочой норадренали-на, а при состояниях тревоги — выведение адреналина [835]. Адреналин вызывает сужение сосудов вомногих органах и тканях, за исключением скелетной мускулату-
нейрофизиологические механизмы гипертонии 341
ры и печени, где под влиянием адреналина сосуды расширяются. Следовательно, адреналин вызывает лишь незначительное повышение периферического сопротивления, а иногда даже его падение, поэтому гипертонию, вызванную адреналином, можно отнести в основном за счет увеличения работы сердца. Норадреналин, напротив, вызывает сужение не только тех сосудов, которые суживаются под действием адреналина, но и сосудов печени и скелетной мускулатуры, поэтому общее периферическое сопротивление при действии норадреналина значительно возрастает.
Одного этого достаточно для повышения кровяного давления; неудивительно поэтому, что в соответствующих экспериментах не наблюдалось увеличения ударного объема сердца. Нужно добавить, что как при «тревоге», так и при «негодовании» почечный кровоток уменьшается; этого не наблюдают только у больных, перенесших симпатэктомию.
Доверительная беседа с психиатром вызывает активацию симпатической системы, аналогичную той, которая наблюдается при выполнении сложного задания, при болевом раздражении или при введении гистамина (в последнем случае, очевидно, происходит рефлекторное раздражение симпатических центров за счет снижения кровяного давления). Это ведет к уменьшению времени свертывания крови, повышению вязкости и увеличению объема эритроцитов [805].
Оказалось, что выработать симпатический условный рефлекс у человека довольно легко. Используя сужение сосудов при погружении руки в холодную воду в качестве безусловной реакции и сочетая ее с условным сигналом (зуммер, свет или словесный раздражитель), можно заметить, что последний вскоре тоже начинает вызывать сужение сосудов. Особенно интересно, что спустя 30— 60 дней интенсивность условной реакции становится выше, чем непосредственно после выработки условного рефлекса, хотя на протяжении этого интервала безусловное подкрепление не применялось. У некоторых испытуемых даже воспоминание или мысль об условном сигнале вызывала сужение сосудов [639]. Из этого ясно, что любой фактор, связанный с эмоциональным стрес-
ГЛАВА ХШ
сом, может вызвать симпатическую реакцию спустя много времени! Частое повторение симпатических эмоциональных реакций создает соответствующую основу для процессов суммации, которые могут постепенно вызывать усиление активности симпатической системы.
Обычными признаками повышения симпатической реактивности у больных гипертонией в эмоциональных ситуациях являются: усиленное потоотделение, тахикардия и вазомоторная неустойчивость, характеризующаяся заметными колебаниями кровяного давления и сердечного ритма в сочетании с гиперемией лица. Шредер [809] обозначил эту поразительную эмоциональную реакцию термином «гипертензивный диэнцефальный синдром»; он указал, что при наличии повышенной чувствительности эту реакцию легко вызвать подкожным введением гистамина. Наблюдения показывают, что у больных, предрасположенных к гипертонии, эмоциональные расстройства связаны с более продолжительными симпатическими разрядами, чем у здоровых людей, а патогенное действие этих разрядов определяется индивидуальными особенностями и внешними факторами (степень и частота эмоционального стресса). В свете данных о роли гипоталамуса в эмоциональных состояниях можно предположить, что усиленные симпатические разряды в гипоталамусе играют основную роль во включении процессов, приводящих к гипертонии. Одним из таких примеров может служить ишемия почек, вызванная путем воздействия на симпатическую систему.
Возражения, часто приводимые против такой роли симпатической системы в патогенезе гипертонии, не кажутся убедительными. Среди них можно указать на следующее. Наличие сужения периферических сосудов после симпатэктомии; это может быть вызвано необратимыми анатомическими изменениями и повышением фонового тонуса (увеличение «автоматизма») гладкой мускулатуры стенок денервированных кровеносных сосудов [592]; нужно также учитывать изменение самих симпатических волокон. Симпатэктомия не устраняет гипертензивного влияния словесных эмоциональных раздражителей на кровяное давление. Дело в том, что эмоциональное возбуждение вызывает не только симпа-
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ГИПЕРТОНИИ 343
хические разряды в гипоталамусе, но и стимулирует активность передней доли гипофиза и нейрогипофиза. Антидиуретический вазопрессорный гормон ' обнаруживается в крови в большом количестве [653] и, несомненно, играет роль в возникающих при эмоциях гипертепзив-ных реакциях у симпатэктомированных больных. Этой интерпретации не противоречит тот факт, что удаление задней доли гипофиза не приводит к снижению кровяного давления ни в норме, ни при гипертонии [526]. Следует напомнить, что секреция этих гормонов в гипоталамусе продолжается после удаления нейрогипофиза [64, 460]. В этой связи интересно отметить, что экстракт задней доли гипофиза (питрессин), так же как и эмоциональный стресс, вызывает гипертонию в большом кругу кровообращения, не влияя на давление в легочной артерии [683]. Кроме того, следует отметить, что при клинической и экспериментальной гипертонии содержание антидиуретического гормона в моче возрастает. Старые клиницисты строили догадки о роли вегетативной нервной системы в патогенезе гипертонии. Так, Мюллер и Фольгар [670] предполагали, что развитию эссенциалыюи гипертонии предшествует «сосудистый невроз». Мы предпочитаем рассматривать начальную стадию гипертонии как выражение повышенной реактивности симпатического отдела гипоталамуса. Эта повышенная реактивность в значительной степени определяется генетическими факторами и проявляется в психической сфере определенными чертами характера, а в физиологической сфере — симпатотонией в условиях покоя [180]. В норме состояние равновесия вегетативной нервной системы характеризуется преобладанием симпатического отдела. Поэтому симпатическая реакция на эмоциональную конфликтную ситуацию должна считаться физиологической нормой. Этот небольшой перевес симпатической системы развивался в филогенезе параллельно развитию реакций страха и бегства, выполняя, как показал Кэннон, важную физиологическую роль в
* * * *
1 Антидиуретическим и сосудосуживающим действием обладает полипептид, выделенный из задней доли гипофиза [439].
ГЛАВА XIII
трудных ситуациях. Именно повышенная реактивность симпатической системы лежит в основе предрасположения к гипертонии.
РАЗДРАЖЕНИЕ ГИПОТАЛАМУСА, РЕАКТИВНОСТЬ СИМПАТИЧЕСКОЙ СИСТЕМЫ И ГИПЕРТОНИЯ
В свете нашего утверждения о роли повторных разрядов гипоталамуса в возникновении гипертонии представляют интерес физиологические опыты с повторными раздражениями этой структуры, при которых наблюдаются значительные сдвиги равновесия вегетативных

Фиг. 35. Временная суммация в симпатической системе.
У кошки под нембуталовым наркозом раздражали задний отдел правого
гипоталамуса (3,5 в, 139 имп/сек, 1,6 нсеК, 10 сек с интервалами 12 сек).
I — сокращение третьего века; II — кровяное давление; /// — пульс;
IV — раздражение гипоталамуса.
центров. При раздражении симпатического отдела гипоталамуса с определенными интервалами на «периферии» наблюдались характерные признаки преобладания симпатической системы: сокращение третьего века, повышение кровяного давления и ускорение сердечного ритма (фиг. 35). Возможны и более выраженные изменения. Повторное раздражение может нарушить равновесие внутри вегетативной системы, а именно повысить реактивность и частоту разрядов симпатической системы и реципрокно понизить активность парасимпатического отдела [315]. Это явление иллюстрирует фиг. 36: повторные раздражения гипоталамуса вызывают прогрессирующее повышение кровяного Давления и частоты сердечных сокращений, тогда как феномен «отдачи» в парасимпатической системе (замедление сердечного рит-
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ГИПЕРТОНИИ 345
ма с увеличением амплитуды пульсовых колебаний) уменьшается. Существенные изменения реактивности и нарушение равновесия в вегетативной системе лежат в основе усиления симпатических разрядов, а длительное воздействие подобных сдвигов, вероятно, определяет возникновение гипертонии или постепенное развитие болезни Рейио, при которых довольно часто проявляются эмоциональные расстройства.

Фиг. 36. Временная суммация в симпатической системе с уменьшением активности парасимпатической системы.
У кошки под нембуталовым наркозом раздражали задний отдел правого
гипоталамуса (1,5 е, 5,1 имп/сек, 0,8 мсек, 10 сек с интервалами Ш сек).
/ — кровяное давление; // — пульс.
ПОВЫШЕННАЯ ЭФФЕКТИВНОСТЬ ГИПОТЕНЗИВНЫХ ВЕЩЕСТВ ПРИ ГИПЕРТОНИИ
Наконец, нам хотелось бы рассмотреть в свете экспериментальных данных, обсужденных в этой главе, влияние гипотензивных веществ и изменения положения тела при гипертонии.
Такие вещества, как ацетилхолин и мехолил, вызывают более резкое падение давления у больных гипертонией, чем у здоровых людей [325, 840], В свете данных, описанных в гл. V, это может быть следствием либо резкого снижения реактивности симпатического отдела гипоталамуса, либо ослабления влияния барорецепторов, так как известно, что после денервации сино-аортальной зоны действие гипотензивных веществ усиливается и Длительность его возрастает. Поскольку и клинические и экспериментальные данные убедительно свидетельст-
ГЛАВА XIII
вугот о повышении симпатической реактивности' при гипертонии, установленным фактам соответствует только предположение о пониженной активности барорецеп-торов. В пользу этого объяснения свидетельствуют и результаты исследования влияния положения тела на кровяное давление при гипертонии и нормотонии [877а].
В этой работе было установлено, что у лиц с гипертонией кровяное давление падает значительно более резко, чем у здоровых людей. Это можно объяснить следующим образом: при гипертонии активность барорецепто-ров, а соответственно и их понижающее влияние на кровяное давление постоянно понижены по сравнению с нормой, поэтому влияние снижения их активности в результате соответствующего изменения положения на падение кровяного давления оказывается меньше, чем в норме; вследствие этого давление падает значительно более резко. Столь резкое падение давления при нормальной активности барорецепторов вело бы к значительному высвобождению высокореактивных симпатических центров из-под их тормозного рефлекторного влияния. Следовало ожидать, что это вызовет значительно более выраженное повышение выведения норадреналина с мочой, чем наблюдаемое у лиц с нормальным давлением. Однако содержание норадреналина в моче не изменялось! Более того, при дальнейших исследованиях было обнаружено, что в этих же условиях повышенная концентрация норадреналина в венозной крови обнаруживается не при гипертонии, а у людей с нормальным давлением [418а]. Отсюда ясно, что патологически резкое падение кровяного давления, наблюдаемое при соответствующем изменении положения тела, у лиц с гипертонией лучше всего объясняется пониженной чувствительностью барорецепторов. Ранее уже упоминалось, что подобное понижение чувствительности барорецепторов наблюдается и при экспериментальной гипертонии. Гистологические исследования показали, что при гипер-
* * * *
1 Речь идет о более выраженном повышении давления при психическом стрессе [500] и при холодовой пробе [496].
НЕЙРОФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ГИПЕРТОНИИ 347
тоиии в барорецепторах наблюдаются дегенеративные изменения, которые могут быть ответственны за ослабление барорецепторных рефлексов [461].
ЗАКЛЮЧЕНИЕ
Предполагается, что эмоциональное возбуждение ведет к усилению симпатических разрядов и повышению кровяного давления. Сила, частота и длительность этих разрядов связаны, видимо, с определенными индивидуальными особенностями и зависят как от внешних условий, так и от генетических факторов. И те и другие отражаются на активности симпатического отдела гипоталамуса и, возможно, на степени тонических симпатических разрядов '. При эмоциональном возбуждении повышается гипоталамо-гипофизарная активность; вместе с ишемией почек, вызванной симпатическими влияниями, эта активность приводит к изменениям во внутренней среде организма, которые могут служить объяснением повышенной чувствительности к норадреналипу, наблюдаемой при гипертонии [404]. Повышенная чувствительность к норадреналипу обусловлена также анатомическими изменениями [155]. При гипертонии у людей активность барорецепторов не исчезает, а уменьшается из-за повышения кровяного давления — вначале кратковременного, а затем все более стойкого [531].
В опытах на обезьянах [750] развитию гипертонии предшествовал период гипотонии, что свидетельствует о начальном усилении буферных рефлексов. Подобной ги-перреактивиостью, если она длится неделями, можно объяснить медленное развитие гипертонии и связанное с истощением окончательное падение активности барорецепторов, способствующее возникновению гипертонии [7501.
Опустив подробное обсуждение важного факта — развития гипертонии при пережатии почечных артерий,—
* * * *
Индивидуальными различиями можно объяснить, по-видимому, "т факт, что повышенное выведение норадреналина [917] наблюдается только в 30% случаев гипертонии-
ГЛАВА XIII
подчеркнем, однако, что симпатические разряды гипоталамуса при эмоциональном стрессе ведут к резкому сужению почечных сосудов с развитием ишемии. Отсюда легко предположить, что возникновение гипертонии у людей с симпатотонией связано с усиленными симпатическими разрядами, хотя на более поздних этапах в процессе начинают принимать участие изменения почечного метаболизма. На этих стадиях изменения в почках не зависят более от нейрогенных разрядов. Доказательством этого служит гипертензивное влияние ишемизи-рованного трансплантата почки, лишенного иннервации ([67], см. также [710, 809]).
Авторы полностью отдают себе отчет в том, что данное объяснение не решает проблему гипертонии, но надеются, что оно может оживить интерес к важному, но забытому аспекту экспериментальных исследований в этой области.
ГЛАВА XIV
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЯЗВЕННОЙ БОЛЕЗНИ
Со "Времен классических исследований Павлова известно, что влияние так называемых «психических» факторов на желудочную секрецию осуществляется через блуждающий нерв.
Вид или запах пищи стимулирует выделение желудочного сока. Этот процесс можно изучать количественно, если изолировать часть желудка и снабдить ее фистулой, благодаря которой можно определить величину секреции и состав желудочного сока. Такие «изолированные желудочки» разной формы с сохраненным кровоснабжением довольно широко использовались при изучении физиологии желудочно-кишечного тракта и патогенеза язвы желудка. Пережевывание пищи сопровождается секреторной активностью желез изолированного желудочка; если иннервирующие желудок блуждающие нервы перерезаны, подобная секреция не наблюдается. При нормальном пищеварении эта «психическая» фаза возникает рано; об этом можно судить по результатам наблюдения над больными с непроходимостью пищевода. Во время психической фазы выделяется приблизительно 20% всего желудочного сока. За этой фазой следует вторая, «желудочная», начинающаяся с момента контакта пищи со слизистой оболочкой привратника, выделяющего специфический гормон гастрин.
ТЕОРИЯ ПАТОГЕНЕЗА ЯЗВЕННОЙ БОЛЕЗНИ
Считалось, что язва двенадцатиперстной кишки возникает в результате усиленной секреции желудочного сока и разрушающего действия содержащейся в нем кислоты. Другая гипотеза — о циркуляторном механизме возникновения язвы — была быстро отвергнута
ГЛАВА XIV
из-за невозможности вызвать язвенную болезнь у экспериментальных животных путем воздействия на кровоснабжение желудка [196, 197], тогда как теория повышенной кислотности получила дополнительное подтверждение в многочисленных экспериментальных и клинических работах.
Дрэгстедт показал, что при язвенной болезни 'ночная секреция в 4—20 раз выше, чем в норме. При пересечении блуждающих нервов на уровне диафрагмы эта секреция, оставаясь все же выше, чем в норме, снижалась на 50% и более. Для получения соответствующего эффекта пересечение должно быть полным или почти полным, так как известно, что импульсы, поступающие по блуждающему нерву, распространяются через мей-снерово сплетение на весь желудок, так что незначительное число волокон блуждающего нерва может влиять на секрецию обширных областей слизистой желудка. Операции, проведенные более чем на 1000 больных по поводу язвы двенадцатиперстной кишки, в 90% случаев были удачными — прекращались боли и рубцевалась язва.
Представление о роли повышенной желудочной секреции в патогенезе язвы двенадцатиперстной кишки подтверждается опытами на животных: длительная стимуляция желудочной секреции приводит к появлению язвы. В этих опытах можно использовать гистамин— мощный стимулятор желудочной секреции; содержащие гистамин пилюли из пчелиного воска имплантируют в мышцы животного. Из таких пилюль гистамин освобождается постепенно. Ежедневная имплантация таких пилюль, как правило, ведет к появлению язвы [433]. Поскольку ваготомия ослабляет влияние гистамина на секрецию, можно думать, что в возникновении данной формы язвенной болезни играет роль рефлекторная стимуляция секреции через блуждающий нерв.
Эти наблюдения подтверждаются экспериментами, в которых длительную стимуляцию секреции осуществляли другими способами. Известно, что мнимое кормление собак с (пистулой пищевода вызывает появление язвы желудка. В основе этого эффекта лежит стимуляция «психической» фазы секреции [506].
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЯЗВЕННОЙ БОЛЕЗНИ 351
Кроме «психической» фазы секреции, существует так называемая «желудочная» фаза, во время которой слизистая привратника выделяет специфический гормон гастрин. Для стимуляции активности этого физиологического механизма были проведены сложные операции на собаках. При этом «изолированный желудочек» (с сохранением иннервации и кровоснабжения) создавали почти из всего желудка, за исключением привратника. Привратник соединяли с кардией так, чтобы пища в желудке не скапливалась. Изолированная часть, закрытая с проксимального конца, открывалась в двенадцатиперстную кишку таким образом, чтобы исключить попадание щелочных соков печени и поджелудочной железы. Уменьшение вместимости желудка в результате операции вынуждало собаку есть в течение многих часов малыми порциями, что и приводило к увеличению продолжительности секреции в изолированном желудочке. Более того, длительный контакт пищи с областью привратника создавал оптимальные условия для стимуляции секреторной активности гастрином. В результате у 100% собак возникли язвы желудка. У другой группы собак удаляли привратник и соединяли кардиальную часть с двенадцатиперстной кишкой. У этих животных гормональная фаза исключалась, но Удлинения психической фазы оказалось достаточным Для того, чтобы в 45% случаев возникли язвы. Когда в этом опыте произвели перерезку блуждающих нервов, то язва возникла лишь у одной собаки из 21 (5%). Эти остроумные эксперименты показывают, что физиологические условия, вызывающие усиленную желудочную секрецию, ведут к образованию язв; причем частота их появления зависит от кислотности [790]. Более того, в эксперименте установлено, что устранение влияния блуждающего нерва путем ваготомии или фармакологического воздействия (атропин, бантин) предотвращает появление язв [13].
ЭМОЦИОНАЛЬНОЕ НАПРЯЖЕНИЕ И ЯЗВЕННАЯ БОЛЕЗНЬ
Имеются достаточные основания для предположения 0 том, что эмоциональное напряжение определенным °бразом влияет на желудочную секрецию и возпикнове-
ГЛАВА XIV
ние язвенной болезни. У собак вырабатывали условно-рефлекторную реакцию страха. В определенной части проб зуммер сопровождался сильным болевым раздражением, наносимым с различными интервалами. У животных развивались симптомы постоянного страха. Они сопровождались усиленной желудочной секрецией и ускорением опорожнения желудка; язвы не возникали [618]. Очевидно, нужно было усилить или продолжить состояние напряжения или же проводить эксперименты на более реактивных животных, так как известно, что у собак уравновешенного типа экспериментальные неврозы, а возможно, и другие психосоматические нарушения не развиваются даже в наиболее тяжелых условиях [686]. У крыс в аналогичных условиях наблюдали возникновение язв. Животных помещали в ящик, где на пути к пище и воде располагалась решетка, которая находилась под током в течение 47 час из 48. Любая попытка добраться до пищи или воды сопровождалась болевым раздражением. У экспериментальных животных развивались язвы, и многие из них погибали от желудочного кровотечения, тогда как у контрольных животных, не получавших ни пищи, ни воды в течение 47 час из 48, язвы не возникали [791, 937].
У здоровых и больных людей эмоциональный стресс вызывает повышение желудочной секреции. Это установлено у студентов во время экзаменационной сессии [618] и у больных, у которых беседа с врачом вызывала беспокойство, гнев или возмущение [957].
Нужно добавить, что при подобных обстоятельствах довольно легко образуются условные связи. Так, если у больного несколько раз при беседе с психиатром отмечалось повышение желудочной секреции, то оно наблюдается с наступлением обычного для беседы времени, даже если беседа не состоялась [877],— время как бы само становится условным сигналом. Эксперименты показывают, что многие факторы, связанные с возникновением эмоций, могут вызывать условнорефлекторное повышение желудочной секреции на длительное время, способствуя таким образом развитию язвенной болезни. У больных с язвой желудка в ночь перед операцией наблюдается более высокий по сравнению с предыду-
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ЯЗВЕННОЙ БОЛЕЗНИ 353
щими ночами уровень желудочной секреции [199]. Влияние разных видов эмоционального стресса на желудочную секрецию у здоровых людей и у больных с язвой исчезает после ваготомии [874]. Эти данные позволяют предположить, что у больных с язвой повышена возбудимость переднего отдела гипоталамуса, регулирующего парасимпатические реакции, возникающие на эмоциональной основе; проведенные недавно исследования показали, что для таких больных действительно характерна ваготония.
В свете этих данных естественно предположить, что люди с повышенной желудочной секрецией составляют тот контингент, из которого рекрутируются больные с язвой.
Определение содержания пепсиногена в сыворотке (которое коррелирует с повышенной кислотностью желудочного сока), проведенное на тысячах призывников, показало, что у 15% обследованных из группы с гиперсекрецией на протяжении 8—16 недель военного обучения развивалась язва двенадцатиперстной кишки. Среди лиц с пониженной секрецией ни одного подобного случая не было. Необходимым элементом при этом, конечно, является эмоциональное напряжение — в данном случае военная подготовка; однако индивидуальная повышенная реактивность также необходима. Выше мы отмечали, что эмоциональное возбуждение ведет к усилению секреции антидиуретического гормона, которая регулируется супраоптическим ядром переднего отдела гипоталамуса. "Тот факт, что антидиуретическая активность сыворотки у больных с язвой в 3 раза выше, чем У здоровых людей [652], также подтверждает предположение о повышенной реактивности переднего отдела гипоталамуса у таких больных.
ГИПОТАЛАМУС И ЖЕЛУДОЧНАЯ СЕКРЕЦИЯ
Клинические данные и результаты экспериментов на животных разных видов показали, что возникновение язв в желудочно-кишечном тракте при повреждениях мозга обычно связано с травмой гипоталамуса [262]. О важной роли повышенной секреции в этом процессе
12-51?
ГЛАВА XIV
свидетельствует отсутствие язв у животных, которым вводят атропин [262].
Интересные результаты были получены в экспериментах с раздражением. В первых опытах с раздражением переднего отдела гипоталамуса было обнаружено быстро наступающее повышение тонуса и активности желудочно-кишечного тракта [50]; более поздние исследования на обезьянах показали, что раздражение этой: структуры на уровне перекреста зрительных нервов вызывает также секрецию желудочного сока, кислотность которого достигает максимума спустя 1 час после раздражения. Этот эффект устраняется ваготомией [722]. При введении электродов в различные отделы гипоталамуса и ежедневном раздражении в течение 30 дней и более у большинства обезьян возникли язвы, обнаружившие «поразительное» сходство с клинической картиной язвенной болезни [263]. Язвы возникали при стимуляции переднего или заднего отделов гипоталамуса и были связаны со значительным повышением кислотности желудочного сока. Существуют указания на то, что изменения в сосудистой и двигательной сфере тоже способствуют возникновению язв. Не удивительно, что мирные и дружелюбно настроенные животные после длительных опытов с раздражением, вызывающим эмоциональные реакции, становятся беспокойными и легко возбудимыми. Необходимо добавить, что даже в острых опытах на кошках при раздражении переднего отдела гипоталамуса Фельдману и др. [242] удалось вызвать, эрозии и геморрагические изменения слизистой.
В связи с этими экспериментами возникает важный вопрос, какова роль заднего отдела гипоталамуса в патогенезе язвенной болезни. Некоторые авторы [261, 722J приписывают ему такую же важную роль, как и переднему, поскольку раздражение заднего отдела гипоталамуса приводит к повышению желудочной секреции; правда, оно возникает со значительной задержкой по сравнению с тем же эффектом, связанным с раздражением переднего отдела гипоталамуса. Более того, вызванное таким раздражением повышение секреции не зависит от блуждающего нерва, но устраняется при удалении надпочечников. Секреторная активность моз-
БИОЛОГИЧЕСКИЕ ОСНОВЫ ЯЗВЕННОЙ БОЛЕЗНИ 355
ового вещества надпочечников, очевидно, не играет при ,т0м существенной роли, так как перерезка спинного мозга на уровне шейного отдела, препятствующая распространению симпатических импульсов из гипоталамуса к мозговому веществу надпочечников, не оказывает влияния на желудочную секрецию, вызванную раздражением гипоталамуса. Можно, следовательно, предположить, что отсроченное повышение желудочной секреции обусловлено активацией гипоталамо-гипофизар-ной системы, вызывающей секрецию АКЛТ передней долей гипофиза с последующим повышением секреции адренокортикостероидов. Правильность такого предположения подтверждается тем, что введение АКТГ и кортизона вызывает у обезьян отставленную во времени желудочную секрецию, не изменяющуюся при ва-готомии, но устраняющуюся при удалении надпочечников. Кроме того, было показано, что повышение желудочной секреции под влиянием инсулина складывается из двух компонентов. Известно, что после введения инсулина нормальному животному секреция достигает максимума через 1 — I х/г час и остается на этом уровне в течение 5 час. Ваготомия вызывает задержку реакции, а удаление надпочечников уменьшает ее длительность. При сочетании обоих воздействий реакция па инсулин устраняется. К сожалению, Дрэгстедту и др. [200] не удалось показать влияние АК.ТГ, кортизона, инсулина и адреналина на секреторную активность изолированного желудочка собаки. Пока не ясна причина этой неудачи, и до получения новых данных можно лишь подчеркнуть важную роль переднего отдела гипоталамуса и желудочной секреции, обусловленной активностью блуждающего нерва, в возникновении язвенной болезни.
Данные об усилении желудочной секреции по ночам у больных с язвой согласуются с данными о сдвиге вегетативного равновесия во время сна. Активность симпатических центров гипоталамуса во время сна понижена. В связи с этим, согласно принципу реци-прокности, происходит активация парасимпатического отдела. Эта активация выражена столь четко, что Гесс [453] высказал предположение об активной природе сна,
12*
ГЛАВА XIV
связанной с возбуждением парасимпатической системы, аналогичным тому, которое наблюдается при электрическом раздражении ядер средней линии диффузной таламической системы и переднего отдела гипоталамуса. Каковы бы ни были наши теоретические представления о механизме возникновения сна, ясно одно,- что во время сна активность переднего отдела гипоталамуса и связанных с ним диэпцефальных структур (гиппоген-ная зона таламуса, по Гессу) повышена, т. е. доминирует парасимпатическая (трофотропная) система. Этим объясняется и более выраженное повышение желудочной секреции по ночам у больных с язвой желудка. При сдвиге равновесия вегетативных систем гипоталамуса в сторону преобладания парасимпатического отдела (как при язвенной болезни) снятие торможения выявляет степень активности, присущую данному отделу. При этом нет необходимости искать какие-то нарушения активности нижележащих центров, как это делает Дрэгстедт.
РАЗЛИЧИЯ МЕЖДУ ЯЗВОЙ ЖЕЛУДКА И ЯЗВОЙ ДВЕНАДЦАТИПЕРСТНОЙ КИШКИ
Физиологическими и клиническими исследованиями язвы двенадцатиперстной кишки была твердо установлена роль нейрогенных факторов в патогенезе этой болезни и предложено лечение, оказавшееся успешным по крайней мере в 90% случаев. Основываясь на этом, можно добавить несколько слов о причинах возникновения язвы желудка. Желудочная секреция начинается с «психической» фазы (в возникновении которой основную роль играет блуждающий нерв) и переходит в «желудочную» фазу. Гормон гастрин, вырабатываемый в привратнике при контакте с пищей, обеспечивает нейтральную или слегка кислую реакцию желудочного сока. Этот гормон поступает в кровоток и вызывает желудочную секрецию. Об этом свидетельствуют, в частности, опыты с изолированным желудочком Гайден-гайна (изолированная хирургическим путем часть желудка с нормальным кровоснабжением, но лишенная иннервации), когда пища воздействует на основную
ГКЧЕСКИЕ ОСНОВЫ ЯЗВЕННОЙ БОЛЕЗНИ 357
пСть желудочка. Секреция в желудочке Гайденгайна практически отсутствует, если удалить привратник, который выделяет гастрии. Усиливая желудочную секрецию вызванную нейрогеииым путем, этот гормональный'фактор обеспечивает адекватное выделение желудочного сока для пищеварения. Избыток гормона может привести к чрезмерной секреции желудочного сока и в конце концов к возникновению язвы. Это можно наблюдать в эксперименте, если пища задерживается в привратнике в результате спазма сфинктера или ослабления перистальтики желудка вследствие ваготомии [198]. Лечение этой болезни осуществляется соответствующими операциями на желудочно-кишечном тракте, в результате которых пища перестает задерживаться в области привратника и устраняется область выделения гастрина. Поскольку активность блуждающего нерва в значительной степени определяет перистальтику желудка, легко понять, почему после ваготомии у больных с язвой двенадцатиперстной кишки наблюдается стаз пищи и почему сочетание этой операции с гастроэите-роанастомозом дает оптимальный результат.
Эти наблюдения иллюстрируют существование строго сбалансированного равновесия между вегетативной (вагусной) и гуморальной регуляцией функции желудка. Если в результате эмоциональных (гипоталамиче-ских) нарушений возрастает активность блуждающего нерва, то развивается язва двенадцатиперстной кишки, обусловленная избыточной секрецией кислого желудочного сока. Если же «психическая фаза» секреции устраняется ваготомией, то понижение двигательной активности желудка и секреции кислоты создают условия для выделения максимальных количеств гастрина, что в результате ведет к развитию язвы желудка. Теоретически можно ожидать, что колебания степени возбуждения блуждающего нерва в ходе пищеварения в желудке могут компенсироваться соответствующими изменениями интенсивности секреции гастрина. Так, недостаточная длительность психической фазы приводит к тому, что продолжительность нахождения нейтрального содержимого в желудке возрастает, что в свою очередь приводит к усиленной секреции гастрина, компенсирую'
ГЛАВА XIV
щей дефицит секреции, возникший в психической фазе. Наоборот, если выделяется много «запального» сока, связанное с этим возрастание моторной активности желудка и повышение кислотности желудочного сока создают неблагоприятные условия для выделения га-стрина, благодаря чему суммарная секреция удерживается в нормальных пределах. В общем эти теоретические рассуждения оправдываются. Однако изменения функции самого желудочка (например, при спазме сфинктера) или центральные расстройства (гипоталамус) могут серьезно нарушать это тонко сбалансированное равновесие. Чисто количественные физиологические сдвиги, как в приведенных выше примерах, могут привести к патологии.
ДАЛЬНЕЙШЕЕ ИЗУЧЕНИЕ РОЛИ УСЛОВНЫХ РЕФЛЕКСОВ В ПАТОГЕНЕЗЕ ЯЗВЕННОЙ БОЛЕЗНИ
В последние годы на обезьянах были поставлены эксперименты по выработке условных рефлексов различной степени сложности [720]. В простейшем случае (условнорефлекторная реакция избегания) через 20 сек после светового сигнала (красная лампочка) лапу животного раздражали электрическим током; животное могло избежать этого раздражения нажатием на рычаг с интервалом менее 20 сек. При этом и подобных воздействиях, применявшихся в течение нескольких недель по 6 час ежедневно, у большинства животных возникли поражения желудочно-кишечного тракта; отмечалась усиленная секреция кислоты в желудке, но никаких признаков повышения секреции АКТГ, осуществляемой через гипоталамус, не наблюдалось.
Особый интерес представляют эксперименты, проведенные на парах обезьян, подвергавшихся одному и тому же воздействию при условии, что только одно животное из пары «владело» рычагом. Именно у этого («ответственного») партнера под влиянием стресса возникала язва!
В связи с этими интересными опытами возникает масса вопросов. В какой мере использованные приемы специфичны для наблюдавшихся нарушений? Как нуж-
1ГИЧЕСКИЕ ОСНОВЫ ЯЗВЕННОЙ БОЛЕЗНИ 359
изменить воздействие и можно ли это сделать, что-11 вызвать гипертонию? Имеет ли какое-нибудь значение, характер эмоционального напряжения, если учесть, что гипоталамус находится в состоянии соответствующей вегетативной настройки?
Даже если допустить, что общий тип психосоматического расстройства, возникающего у разных людей и животных, зависит от равновесия центральных вегетативных механизмов, поддерживаемого взаимодействием генетических факторов и факторов внешней среды, останется без ответа важный вопрос о специфическом характере расстройства. Аналогичные ситуации вызывают изменение кровяного давления у одного человека, нарушение сердечной деятельности у другого и расстройство функции желудка у третьего. Представляется вероятным, что в основе таких специфических нарушений вегетативных функций лежат процессы суммации вегетативных (гипоталамических) разрядов с висцеральными рефлексами. Подобно тому как подпороговая стимуляция гипоталамуса значительно усиливает влияние раздражения седалищного нерва на соответствующие висцеральные органы, проявляющееся в изменении кровяного давления, сердечного ритма и т. д. [315], разряды, возникающие в различных внутренних органах, могут суммироваться с разрядами гипоталамуса и оказывать значительное влияние на тот или иной определенный орган или даже на его отдельные функции (например, ритм сердца). Работы Курцина [557] действительно показывают, что локальные висцеральные рефлексы играют важную роль в возникновении поражений желудка в описанном выше эксперименте с выработкой условного рефлекса.
ЗАКЛЮЧЕНИЕ
Мы рассмотрели гипертонию и язвенную болезнь как примеры психосоматических нарушений, при которых играют важную, хотя и не до конца понятную роль соответственно симпатическая и парасимпатическая системы. Мы не упомянули здесь о многих других болезнях, в патогенезе которых видное место принадлежит
Г ЛАБА XIV
эмоциональным расстройствам, просто потому, что не считаем себя достаточно компетентными в сугубо клинических проблемах. Необходимо еще раз подчеркнуть, что как симпатическая, так и парасимпатическая системы способны активироваться при раздражении вегетативных центров; этот факт согласуется с рядом клинических наблюдений. Эмоциональное напряжение может вызвать явное ускорение сердечного ритма без каких бы то ни было изменений кровяного давления у одних людей и диаметрально противоположный эффект —-у других. Вольф [956] показал, что в отношении области соматических поражений, возникающих под влиянием эмоционального возбуждения, существуют выраженные индивидуальные различия, и предлагал даже различать по этому признаку «желудочно-реактивных», «пульсо-реактивных», «прессо-реактивпых» и т. д. Вполне возможно, что клинические симптомы обусловлены специфическим возбуждением в определенных отделах вегетативной нервной системы, имеющим тенденцию сохраняться в течение длительного времени [283].
В основе такого рода нарушений лежит повышенная чувствительность центральных механизмов вегетативной нервной системы к воздействиям эмоционального характера. В гл. IX мы описывали эксперименты, показывающие, что слова, несущие эмоциональную нагрузку, вызывали симпатическую реакцию у испытуемых при весьма кратковременной экспозиции. Сходные эксперименты были проведены на обезьянах, которых обучали нажимать на рычаг при определенном зрительном раздражении, за которым следовал электрический удар. Несмотря на то что путем нажатия на рычаг животное избегало болевого раздражения, одно лишь зрительное раздражение вызывало ускорение сердечного ритма. Если же время предъявления укорачивали до 0,7 мсек так что раздражитель явно не воспринимался и реакция на него не возникала, наблюдалось замедление ритма сердца [653]. Возбуждение парасимпатической системы возникает в ответ на «неосознанный» раздражитель. Это наблюдение, действительно, стимулирует дальнейшие исследования, имеющие целью расширение наших представлений о физиологии и патологии эмоций.
Г л л в л XV
Изучение гипертонии и язвенной болезни в эксперименте показало, что многократное возбуждение вегетативной нервной системы, возможно связанное с… На правильность такого предположения указывает и тесная зависимость,… 362 Г Л А В A XVA XVIII
И СИМПАТИЧЕСКАЯ СИСТЕМА Поскольку твердо установлено, что эмоциональное возбуждение может привести к… Имеется и другой ряд доказательств участия гипоталамуса в возникновении эпилептических судорог. Во время припадка…Л XX
СОДЕРЖАНИЕ САХАРА В КРОВИ
И СОСТОЯНИЕ ВЕГЕТАТИВНОЙ НЕРВНОЙ
СИСТЕМЫ ПРИ НЕВРОЗАХ И ПСИХОЗАХ
Экспериментальные исследования на животных (гл. IV) показали, что нервная регуляция содержания сахара в крови осуществляется не только через симпа-то-адреналовую систему, что было давно известно, но также и через ваго-инсулиновую. Возбуждение правого блуждающего нерва, иннервирующего островки Лапгер-ганса, физиологическими методами или медикаментами ведет к снижению содержания сахара в крови. При разных стресс-состояниях, включающих эмоциональное возбуждение, активируются обе эти системы. Если при этих условиях влияние симпато-адреналовой системы, повышающей содержание сахара в крови, превалирует над влиянием ваго-инсулиновой системы, действующей в противоположном направлении, то при таком эмоциональном возбуждении возникает гипергликемия. Однако скрытое участие в этом ваго-инсулиновой системы доказывается тем, что субдиафрагмальная ваготомия усиливает гипергликемию, наступающую при стрессе. Более того, если устранить действие симпато-адреналовой системы путем удаления мозгового вещества надпочечников, то под влиянием сходного стресса возникает гипогликемия. Отсюда ясно, что эффект влияния эмоционального напряжения на сахар крови определяется центральным взаимодействием указанных систем.
Если перенести эти физиологические данные в клинику и применить их, в частности при изучении неврозов и психозов, то, исследуя содержание сахара в крови, можно судить о состоянии равновесия между двумя отделами вегетативной системы и об их отношении к эмоциональному состоянию больного.
СОДЕРЖАНИЕ САХАРА В КРОВИ ПРИ НЕВРОЗАХ И ПСИХОЗАХ 441
АКТИВНОСТЬ ВАГО-ИНСУЛИНОВОЙ СИСТЕМЫ ПРИ НЕВРОЗАХ
Некоторые авторы отмечают, что состояние напряженности и утомление могут быть связаны с определенной степенью гипогликемии. У больных с нарушениями личности при состоянии напряженноеш и депрессии выявляются два симптома, свидетельствующие о повышенной реактивности ваго-инсулиновой системы: уплощение сахарной кривой и гипогликемия после еды [748]. Повышение содержания сахара в крови после внутривенного введения глюкозы или плотной еды стимулирует секрецию инсулина. Как показали эксперименты Цунца и Ла-Барра [971] с перекрестным кровообращением, повышение содержания глюкозы в крови воздействует на центр ваго-инсулиновой системы па уровне промежуточного мозга. Если вследствие сдвига вегетативного равновесия в сторону преобладания парасимпатической системы этот центр находится в состоянии повышенной реактивности, сахарная кривая уплощается и гипергликемия сменяется гипогликемией.
Портис и Александер обнаружили аналогичное изменение сахарной кривой у большой группы больных-психоневротиков [12, 722а]. При этом у них наблюдалась утомляемость (как хроническая, так и в виде острых приступов), понижение работоспособности, апатия, общая подавленность и различного рода фобии.
Теоретически можно предположить, что такая повышенная реактивность ваго-инсулиновой системы устранима двумя путями: блокадой передачи разрядов "блуждающего нерва к островкам Лангерганса или восстановлением центрального равновесия в вегетативной системе. И действительно атропинизация и ваготомия (блокирующие импульсы блуждающего нерва) меняют характер сахарной кривой. Более того, аналогичный эффект наблюдается при психологических воздействиях, приводящих к улучшению эмоционального состояния. Предполагается, что благодаря таким воздействиям восстанавливается равновесие вегетативных систем гипоталамуса. (•■Индром утомления и отсутствия интереса к окружающему у таких больных получил название «вегетативного
442 г л а в л хх
отступления». Опираясь на только что описанные .наблюдения, мы предложили бы назвать его синдромом гипо-таламической парасимпатотонии.
Исследование таких больных и сравнение их ее здоровыми позволило получить дополнительные данные о гипоталамической системе. Исследование производили: 1) во время- отдыха, 2) при демонстрации кинофильма и 3) во время игры в шахматы. Для определения содержания сахара в крови через короткие промежутки времени брались пробы крови. Оказалось, что при неврозах индивидуальные колебания уровня сахара крови значительно более выражены, чем в контрольной группе; при усилении напряжения эти колебания в обеих группах возрастают [614]. Поскольку сдвига в направлении колебаний не наблюдалось, можно предположить, что они вызваны разрядами ваго-инсулиновой и симпато-адрена-ловой систем, амплитуда которых растет с увеличением напряженности, причем у невротиков в большей степени, чем у здоровых людей. Сходные колебания уровня симпатических разрядов у «импульсивных» больных описал Лейси (см. гл. IV).
Мы уже упоминали, что у лиц с «вегетативным отступлением» во время парасимпатических разрядов, приводящих к гипогликемии, отмечается утомляемость, склонность к обморокам и т. д. Следует, однако, подчеркнуть, что в основе этого синдрома лежит не одна гипогликемия, так как после введения глюкозы, вызывающего гииергликемию, появление соответствующих симптомов при последующем падении уровня сахара крови предшествует достижению им критического уровня. Это свидетельствует о связи доминирующего парасимпатического разряда с восходящим разрядом гипоталамуса, ответственным за этот синдром. На этом основании можно полагать, что on сходен с синдромом, наблюдающимся при тошноте и кратковременных обмороках центрального происхождения (см. гл. XV и XVI), который, согласно нашему анализу, обусловлен восходящим и нисходящим парасимпатическими разрядами гипоталамуса.
Следует добавить, что эксперименты Маля [618] свидетельствуют о связи состояния хронического страха
I
СОДЕРЖАНИЕ САХАРА В КРОВИ ПРИ НЕВРОЗАХ И ПСИХОЗАХ 443
со сдвигом вегетативного равновесия в сторону преобладания ваго-инсулиновой системы.
РЕАКТИВНОСТЬ ВАГО-ИНСУЛИНОВОЙ СИСТЕМЫ ПРИ ПСИХОЗАХ
Хорошо известно, что эмоциональное возбуждение вызывает значительное повышение уровня сахара крови у здоровых людей, но не приводит к подобным изменениям у больных шизофренией [82, 373, 946]. С целью получения дополнительных данных о состоянии равновесия ваго-инсулиновой и симпато-адреналовой систем было проведено специальное исследование: у здоровых людей и у больных шизофренией, находившихся в состоянии покоя, определяли уровень сахара крови и содержание в ней инсулина. Пробу на инсулин ставили на гипофизэктомированных и адреиалэктомированнЫх крысах, весьма чувствительных к инсулину [337]. Пробы показали, что уровень сахара крови и содержание в ней инсулина в контрольной и клинической группах не различаются. Если же животным вводили кровь психически больных, находившихся в состоянии возбуждения (в большинстве опытов кровь больных шизофренией, но иногда и больных с маниакально-депрессивным психозом или паранойей), то происходило резкое падение уровня сахара крови с потерей рефлексов поддержания позы, а часто с гипогликемическими судорогами [338]. Это свидетельствует о высоком содержании инсулина в крови психически больных, находящихся в состоянии возбуждения. У 21 больного возбуждение не привело к повышению уровня сахара крови, но, как показывают результаты проб на крысах, содержание инсулина в крови у этих больных было высоким (табл. 8). У 6 больных наблюдалась незначительная гипергликемия, свидетельствующая о повышении секреторной активности мозгового вещества надпочечников при возбужденном состоянии. Однако кровь 4 больных из этих 6 при введении экспериментальным животным вызывала значительное падение уровня сахара крови. Это означает, что у этих 4 больных концентрация инсулина в крови была повышена по сравнению с нормой, но эффект повышения был
Таблица 8
РЕЗУЛЬТАТЫ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ИНСУЛИНА В КРОВИ БОЛЬНЫХ ПСИХОЗАМИ ПРИ ВОЗБУЖДЕНИИ
| Больной | Содержание сахара и кропи больного, .иг/100 мл | Содержание сахара в крови экспериментального животного, ме/100 мл | Диагноз | ||
| до вводе-пия крови больного | после введения крови больного | ||||
| Не повышено | |||||
| ] | 79,5 | 56,9 | 36,5 | Маниакалыю-депрессив-яый психоз | |
| 81,1 | 59,1 | 33,3 | Психоз, связанный с нарушениями обмена | ||
| 81,1 | 58,0 | 35,4 | То же | ||
| 80,6 | 59,1 | 37,6 | Шизофрения | ||
| 83,8 | 60,2 | 29,0 | >> | ||
| 79,5 | 58,0 | 33,3 | Прогрессивный паралич | ||
| 78,5 | 60,2 | 32,2 | Параноидный психоз | ||
| 81,1 | 59,1 | 36,5 | Шизофрения | ||
| 79,5 | 62,3 | 38,7 | Раннее слабоумие | ||
| 73,1 | 61,2 | 35,4 | » » | ||
| 84,9 | 59,1 | 34,4 | Прогрессивный паралич | ||
| 69,8 | 56,9 | 32,2 | Шизофрения | ||
| 74,1 | 56,9 | 32,2 | » | ||
| 82,7 | 59,1 | 38,7 | Кататоничеекэя форма шизофрении | ||
| 77,4 | 59,1 | 35,4 | Шизофрения | ||
| 74,1 | 60,2 | 39,7 | » | ||
| 84,9 | 69,2 | 34,4 | Острая шизофрения | ||
| 75,2 | 58,0 | 39,7 | Шизофрения | ||
| 77,4 | 60,2 | 36,5 | » | ||
| 80,6 | 56,9 | 34,4 | » | ||
| 82,7 Повышено | 60,2 | 34,4 | » | ||
| 2 3 | 107,5 109,2 102,1 | 58,0 59,1 59,1 | 59,1 45,1 49,4 | Острая шизофрения Шизофрения | |
| 94,6 | 60,2 | 39,7 | Раннее слабоумие | ||
| 132,5 | 61,2 | 56,9 | Маниакально-депрессивный психоз | ||
| 109,2 | 60,2 | 50,5 | Шизофрения |
СОДЕРЖАНИЕ САХАРА В КРОВИ ПРИ НЕВРОЗАХ И ПСИХОЗАХ 445
замаскирован противоположно направленным действием гормона мозгового вещества надпочечников. В отличие от группы больных, в которой появление инсулина в крови при эмоциональном возбуждении наблюдалось в 93% случаев, в контрольной группе, несмотря на значительное возрастание уровня сахара крови при возбуждении, обнаружить инсулина не удалось (табл. 9). При
Таблица 9
РЕЗУЛЬТАТЫ ОПРЕДЕЛЕНИЯ СОДЕРЖАНИЯ ИНСУЛИНА В КРОВИ ЗДОРОВЫХ ЛЮДЕЙ ПРИ ВОЗБУЖДЕНИИ
| Содержание сахара в кропи экспериментального жиг.отного, •кг/100 мл | |||
| Испытуемый | Содержание сахара в кропи испытуемого, я*/100 мл | до введения кропи испытуемого | после введения кропи Испытуемого |
| 96,8 | 60,2 | 64,5 | |
| 94,6 | 58,0 | 58,0 | |
| ' 92,5 | 59,1 | 59,1 | |
| 98,5 | 56,9 | 64,5 | |
| 99,9 | 58,0 | 61,2 | |
| 102,1 | 58,0 | 59,1 |
объяснении полученных результатов следует иметь в виду, что у человека содержание сахара в крови зависит в основном от присутствия адреналина, так как реакции на незначительные количества инсулина, выделяющегося при возбуждении, не отмечается; экспериментальные животные, напротив, реагируют главным образом на инсулин. Две описанные пробы дают возможность понять сущность адренало-инсулинового равновесия. В норме возбуждение сопровождается усиленной секрецией адреналина; секреция инсулина при этом не меняется. При психозах, напротив, возбуждение почти во всех случаях сопровождается усиленной секрецией инсулина, а секреция адреналина не меняется [338].
446 Г Л А В Л XX
На основании этих экспериментов можно сделать вывод, что состояние эмоционального возбуждения в контрольной группе характеризуется преобладанием сим-пато-адреналовой системы над ваго-ипсулиновой. тогда как при психотическом возбуждении наблюдаются об- ] ратные соотношения. Эти данные свидетельствуют о том, что определенные невротические и психотические состояния обычно характеризуются относительным преобладанием ваго-инсулиповой системы. Отсутствие гипергликемии при сильном возбуждении свидетельствует, кроме того, о гипореактивности симпато-адреналовой системы при психозах. (Нам представляется сомнительным предположение о том, что пониженная чувствительность к инсулину у некоторых больных шизофренией имеет отношение к данной проблеме.)
ЗАКЛЮЧЕНИЕ
При психоневрозах, и особенно при синдроме «вегетативного отступления», отмечается гипогликемия и уплощение сахарной кривой, что свидетельствует о повышении активности ваго-инсулиновой системы. Поскольку возникающие при этом симптомы сходны с теми, которые наблюдаются у людей с резко усиленной активностью парасимпатических центров, можно пола- I гать; что это повышение в свою очередь обусловлено доминированием парасимпатического отдела гипоталамуса.
Состояния острого эмоционального возбуждения приводят у здоровых людей к значительной активации симпато-адреналовой системы, проявляющейся в гипергликемии. При психозах, однако, подобная реакция не наблюдается, но отмечается повышение содержания инсулина в крови. Состояние вегетативных центров в этих условиях характеризуется повышением активности ваго-инсулиновой системы при понижении активности симпато-адреналовой.
ГЛАВА Л Л J
ДАЛЬНЕЙШЕЕ ИЗУЧЕНИЕ РОЛИ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ В ВОЗНИКНОВЕНИИ ПСИХОЗОВ
Поскольку в последнее время опубликовано несколько обзоров по этому вопросу [296, 306], мы ограничимся здесь кратким упоминанием о ранних работах Хоскинса и его сотрудников. Изучение терморегуляции при действии холода, сердечно-сосудистых реакций и изменения уровня сахара крови в условиях стресса показало, что при хронической шизофрении реактивность вегетативных центров понижена. Кроме того, отмечено снижение реактивности гипоталамо-гипофизарной системы, в особенности при психическом стрессе. Приводимые ниже результаты, проведенных недавно работ подтверждают эти наблюдения.
ВЕГЕТАТИВНЫЕ РЕАКЦИИ ПРИ ПСИХИЧЕСКИХ ЗАБОЛЕВАНИЯХ
Из всех психически больных наименьшая кожная температура при действии холода отмечена у больных шизофренией [825]. Суточные колебания температуры у них менее выражены, чем у здоровых людей; у больных хронической шизофренией реакция на действие холода либо отсутствует, либо носит парадоксальный характер (повышение температуры вместо понижения [122]). Сопротивление кожи у этих хронических больных как в покое, так и при действии эмоциональных раздражителей выше, чем у здоровых людей и невротиков [235]. Повышение кровяного давления при холодовой пробе выражено слабее, чем в контрольной группе, а иногда даже отсутствует [211, 499]. Сахарная кривая лежит
448 глава xxi
выше, чем у здоровых людей [255]. Подобный эффект можно наблюдать и у здоровых людей при сильном эмоциональном напряжении, у больных же он имеет место даже при ослаблении эмоциональных нарушений [256]. Реакция гипоталамо-гипофизарной системы, приводящая в условиях обменного стресса [253] и при физической нагрузке (действие высоких температур [712]) к высвобождению АКТГ и уменьшению количества лимфоцитов, слабее выражена у психически больных, чем в контрольной группе. По-видимому, эти наблюдения подтверждают точку зрения Хоскинса [488], утверждавшего, что для шизофрении типичен «инертный характер симпатических реакций»; правда, это относится скорее к хро-< нической шизофрении.
В последних работах на основании пробы с мехоли-лом было показано, что средний сдвиг кровяного давления у больных острой шизофренией соответствует по нашей классификации группе I (симпатическая гиперреактивность), тогда как реакцию невротиков того же возраста можно отнести к группе II [841]. Предпринятое нами обследование 105 больных шизофренией (результаты которого не опубликованы) не выявило существенных различий между ними и группой хронических больных, обследованных Нельсоном и Гельгориом [682]. Правда, большое значение имеет возрастной фактор, так как, согласно наблюдениям, процент лиц с симпатической гипореактивностью с возрастом растет.
Как отмечено в гл. VI, среди психически больных вообще и среди больных шизофренией в особенности лица с симпатической и парасимпатической гипо- и гиперреактивностью встречаются значительно чаще, чем в контрольной группе. Случаи нарушения равновесия Berej тативных систем при разных уровнях симпатической активности более часты в группе психически больных, чем в контрольной группе.
При сравнении симпатической реактивности разных групп психически больных на основании пробы с мехо-лилом оказалось, что симпатическая гипореактивность (группа III) характерна для больных с эндогенной депрессией. Эти результаты основываются на анализе 9 случаев, описанных Фанкенштейном (все относятся к
I
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА И ПСИХОЗЫ 449
группе III), 22 случаев из работы Слоэна и на основании данных работы Голда и Альтмана [383, Ь]. Учитывая значение возрастного фактора в определении реактивности к мехолилу, было бы желательно провести новые исследования на сопоставимых возрастных группах. Необходимы также дополнительные исследования для объяснения следующего интересного наблюдения: гипергликемия, вызванная электрошоком, становится менее выраженной при наркозе у больных с депрессией и переходит в состояние гипогликемии у больных шизофренией [424].
При исследовании больных хронической шизофренией применяли лизергиновую кислоту, которая, как было показано, стимулирует гипоталамус и вызывает не только повышение симпатической активности, но и эмоциональное возбуждение и расстройства восприятия. Как и у здоровых людей, у больных наблюдалась реакция расширения зрачка, но психические реакции (смех, 'напряженность, зрительные иллюзии) у больных были минимальными. Интересно, что у нескольких больных, у которых наступило улучшение после электрошоковой терапии, чувствительность к этому веществу повысилась [145, 252]. Вполне возможно, что подобное изменение эмоциональных реакций и восприятия является отражением усиления гипоталамо-кортикальных разрядов. Интересные работы Хилла и др. [463] показали, что при кататоническом ступоре реакция коры на гипогликемию отсутствует; не наблюдается также усиленного потоотделения на ладонях; правда, с улучшением клинического состояния обе указанные реакции восстанавливаются. Мы считаем, что изменения вегетативных, корковых и «психических» функций взаимосвязаны и что состояние возбудимости и равновесия в вегетативной системе играет решающую роль в перечисленных процессах.
СОДЕРЖАНИЕ НОРАДРЕНАЛИНА И АДРЕНАЛИНА В КРОВИ И МОЧЕ ПРИ ПСИХОЗАХ
На существование связи между психическим состоянием и активностью вегетативных центров указывают исследования по изучению выведения адреналина и нор-
15—517
ГЛАВА XXI
адреналина с мочой. У одного и того же больного во время маниакальной фазы маниакально-депрессивного психоза выведение обоих веществ с мочой значительно повышено по сравнению с депрессивной фазой и нормой. Далее, интенсивность выведения норадреналина и адреналина в группе больных, находящихся в депрессивной фазе, была в среднем ниже, чем в группе больных с маниакальной фазой психоза; причем эти -различия оказались статистически достоверными. Хотя и при депрессии у больных порой наблюдались беспокойство и тревога, сопровождавшиеся повышением содержания указанных гормонов в плазме, тем не менее такие состояния встречались реже и были не столь стойки, как при маниакальной фазе. Результаты проб позволяют ав-тчрам предположить существование следующей корреляции: состоянию психического возбуждения соответствует гиперреактивность симпатической системы, а состоянию психической депрессии — гипореактивность этой системы [875]. Характер эмоционального состояния в группе психически больных варьировал от «пассивного самоуничижения» до крайней враждебности; соответственно изменялась интенсивность выведения норадреналина [222]. Риган и Рейли [744], определявшие содержание адреналина и норадреналина в плазме, показали, что существует связь между определенными эмоциональными проявлениями и отношением норадреналин/адреналин, а Сильверман и др. [835] отметили усиленное выведение этих веществ у психически больных по сравнению с невротиками и здоровыми людьми. (Манджер и др. [625], однако, не обнаружили каких-либо характерных изменений содержания этих гормонов при состояниях тревоги.) Как будет подробно описано в гл. XXIII, лизергино-вая кислота вызывает сдвиги психического и симпатического характера; интересно, что ее химические производные, не обладающие психотропным действием, не действуют и на симпатическую систему. Тот же параллелизм наблюдается и у психически больных. В случаях хронической шизофрении, в которых лизергиновая кислота не вызывает изменений настроения, зрительных восприятий и эмоционального состояния, не наблюдается и изменений в интенсивности выведения норадреналина
АТИВНАЯ НЕРВНАЯ СИСТЕМА И ПСИХОЗЫ
и адреналина. У больных маниакально-депрессивным психозом, напротив, обнаруживаются характерные сдвиги в психическом состоянии и параллельно им усиленное выведение норадреиалина и адреналина; при инволюционной депрессии психические сдвиги сопровождаются усиленным выведением одного лишь адреналина, но не норадреиалина [222]. При сопоставлении в процессе дальнейшего изучения реакций на мескалин и лизер-гкновую кислоту у больных хронической шизофренией и здоровых людей выявилась более выраженная чувствительность в контрольной группе. Психические (галлюцинации) и физиологические (определяемые по повышению содержания адреналоподобных веществ в плазме) реакции были отчетливо выражены в контрольной группе и отсутствовали у больных [625].
Эти результаты получены на небольшой группе больных и требуют подтверждения на более обширном клиническом материале. На данном этапе они свидетельствуют о связи между содержанием иорадреналина и адреналина в крови и моче и психическим состоянием больного. У разных групп психически больных наблюдаются качественные изменения реактивности вегетативных центров, проявляющиеся в изменении соотношения между содержанием норадреиалина и адреналина.
Лиделл и Вайл-Малгерб [574] сравнивали содержание норадреиалина и адреналина в спинномозговой жидкости у больных с подозрением на органическую патологию мозга и у больных шизофренией. Отношение норад-реналин/адреиалин было одинаковым, но концентрация обоих веществ была меньше в последней группе. Манд-жер и др. [625] обнаружили пониженное содержание норадреиалина и адреналина при умственной отсталости по сравнению с психозами. Однако эти исследования надо рассматривать как предварительные ввиду недостаточного числа обследованных.
АКТГ И ПСИХОЗЫ
Поскольку секреция АКТГ передней долей гипофиза находится под контролем гипоталамуса и изменяется при его разрушениях, можно ожидать, что сдвиги в ак-
15*
452 глава xxi
тивности гипоталамуса должны оказывать влияние на активность коры надпочечников. Так, в исследованиях Хоглэнда, Пинкуса и их сотрудников [476, 477, 479, 712—714] было показано, что, несмотря на нормальный уровень секреции АКТГ в покое, реакции больных шизофренией на различные виды стресса (определяемые по выведению 17-кетостероидов и мочевой кислоты) резко понижены. При высокой температуре и влажности окружающего воздуха число лимфоцитов в крови здоровых людей понижается, а у больных шизофренией повышается. Психический стресс (пробы на психомоторную координацию и другие аналогичные пробы) вызывал незначительную реакцию со стороны коры надпочечников в группе больных. Повышение секреторной активности коры надпочечников после пробуждения наблюдалось у 50% обследованных здоровых людей и лишь у 3% психически больных.
Дальнейшие исследования выявили высокую эффективность ряда других стресс-факторов при шизофрении. Так, по данным некоторых авторов [21, 479, 644, 696, 896], электрошок, инсулиновая кома и введение адреналина вызывают у больных шизофренией и здоровых людей одинаковое повышение выведения АКТГ. Эти результаты становятся понятными, если учесть, что в секреции АКТГ участвуют несколько механизмов: она может быть вызвана секреторной активностью мозгового вещества надпочечников, непосредственным и рефлекторным возбуждением гипоталамуса или, наконец, через кортико-гипота-ламические связи. Возникновение эозинопении и лимфо-иении у больных шизофренией под влиянием электрошока и отсутствие этих симптомов при психическом стрессе объясняются тем, что в первом случае возникает мощное возбуждение центров симпато-адреналовой системы на уровне продолговатого мозга, отсутствующее в последнем случае. Эти данные подтверждают наш основной тезис об изменении реактивности гипоталамуса при шизофрении.
Интересно отметить, что выведение 17-кетостероидов с мочой у больных шизофренией с параноидным синдро-ном значительно выше, чем у больных, не обнаруживающих этого синдрома [44, 787]. Более того, психическое
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА И ПСИХОЗЫ 453
состояние больных шизофренией отражается на интенсивности выведения стероидов. Так, Бэтт и др. [44] отмечали, что у больных шизофренией максимум секреции наблюдается при состоянии раздражения и агрессивном поведении, тогда как в состоянии покоя секреция становится минимальной. Спонтанные ремиссии сопровождаются постепенной нормализацией уровня кортико-стероидов независимо от того, был ли он до этого выше или ниже нормы. В отдельных случаях маниакально-депрессивного психоза степень интенсивности выведения кортикостсроидов периодически варьирует, причем минимум секреции приходится на гипоманиакальную фазу [121, 761]; у больных с периодической кататонией в первые дни развития очередного приступа выведение 17-кетостероидов возрастает [415]. Эти данные представляются весьма многообещающими в смысле установления корреляции между секрецией адреностероидов, регулируемой гипоталамусом, и психическим состоянием. Однако делать общие выводы пока преждевременно (см. также [871]). Такие явно патологические реакции, как минимальная секреция стероидов при максимальной психомоторной активности, требуют дальнейшего изучения.
Кроме того, работа ворчестерской группы выявила существенные различия между больными шизофренией и здоровыми людьми в отношении выведения натрия и калия [254]. Отмечено далее, что у большинства больных хронической шизофренией в отличие от здоровых людей альдостерои в моче не обнаруживается [480]. Этот факт имеет серьезное значение в свете, новых данных о влиянии промежуточного мозга на секрецию аль-достерона [236].
Данная область исследования затрудняется сложными взаимоотношениями между корой надпочечников, щитовидной железой и половыми железами. Достаточно ясно показано, что реактивность коры надпочечников к действию АКТГ при шизофрении часто понижена [478, 657, 826]. Прогностическая ценность этой реакции сомнительна, а интерпретация данных, касающихся изменения интенсивности выведения стероидов в условиях стресса, представляется еще более затруднительной. Накоплены
454
глава xxi
некоторые данные о качественных различиях выводимых стероидов при шизофрении [473].
Хотя, как известно, введение адренокортикоидов может привести к нарушениям психики, не следует считать деятельность коры надпочечников причиной психических расстройств при шизофрении. Попытки лечения с помощью АКТГ и адреналэктомии не удались [473]; весьма вероятно, что предлагаемое сейчас лечение альдостеро-ном [474] также не будет иметь успеха. Изменение секреторной активности коры надпочечников следует рассматривать как следствие стойких изменений функции гипоталамуса, а не как причину этих заболеваний. Б этом плане интересно, что чувствительность коры надпочечников к действию АКТГ изменяется при поражениях гипоталамуса [173] и что показатели секреции АКТГ (эозиио-пения, выведение кетостероидов и др.) могут не изменяться при шизофрении [747]. Эти наблюдения согласуются с результатами экспериментов Слашера [843]: у крыс с поражением гипоталамуса выделение кортикосте-рона в условиях стресса затормаживалось, но реакция коры надпочечников на аскорбиновую кислоту не изменялась.
ГЛАВА XXII
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ, ЛЕЖАЩИЕ В ОСНОВЕ ШОКОВОЙ ТЕРАПИИ
ЭЛЕКТРОШОК И ДРУГИЕ ВОЗДЕЙСТВИЯ, ВЫЗЫВАЮЩИЕ СУДОРОГИ
Не останавливаясь на вопросе о том, что послужило непосредственной причиной введения электрошоковой терапии и лечения метразолом в психиатрическую клинику, мы ограничимся рассмотрением соответствующих физиологических данных. Повышение уровня сахара крови под влиянием электрошока и метразола и отсутствие этого эффекта после перерезки чревных нервов или удаления мозгового вещества надпочечников свидетельствуют о центральном возбуждении симпатической системы. После удаления мозгового вещества надпочечников эти воздействия усугубляют гипогликемию. Поскольку гипогликемию связывают с сохранением целостности блуждающих нервов, можно сказать, что она вызвана активацией ваго-инсулиновой системы. Хотя во время судорожной активности возбуждаются оба отдела вегетативной нервной системы, симпатический отдел доминирует [239, 343]. Это положение подтверждается и результатами измерения кровяного давления [806]. Дополнительными индикаторами активации симпато-адреналовой системы в этих условиях являются: уменьшение времени свертывания крови [923], максимальное расширение зрачков, сокращение третьего века и повышенная реактивность симпатической системы [333]. В опытах на крысах было показано, что одиночный электрический удар вызывает снижение содержания адреналина в мозговом веществе надпочечников [854].
По поведению человека и животных и по результатам опытов с непосредственным раздражением можно сделать вывод об участии гипоталамуса в осуществлении этих реакций. При подпороговом (в отношении возникно-
456 глава xxti
вения судорог) электрическом раздражении у человека наблюдаются выраженные эмоциональные реакции; пороговое раздражение сопровождается, кроме того, усиленным потоотделением, Как при инсулиновой коме [471]. Аналогичные симптомы наблюдались при введении мет-разола [781]. Выше мы описывали влияние судорог, вызванных электрическим или химическим путем, на секрецию гонадотропных и адренотропиых гормонов передней долей гипофиза [306]. Поскольку перерезка ножки гипофиза прекращает это взаимодействие, можно сделать вывод, что судороги изменяют активность гипоталамуса, которая вторично, через ножку гипофиза, стимулирует секреторную активность последнего.
Из практики применения электрошоковой терапии в психиатрической клинике известно, что действие электрошока на симпатическую систему при повторении растет (фиг. 45) [360].
Гесс [454] показал, что на уровне гипоталамуса реактивность симпатической и соматической систем изменяется параллельно. Эта связь проявляется в том, что при повторных сеансах электрошока возрастает не только симпатическая и эмоциональная реактивность, но и уровень общей активности.
Электрошок и концентрация нор адреналина и адреналина в плазме
Вайл-Малгерб [934], изучая влияние электрошока на содержание норадреналина и адреналина в плазме у психически больных, установил, что концентрация этих гормонов возрастает соответственно на 75 и 40%. Этот эффект можно изменить барбитуратами, веществами, блокирующими синаптическое проведение в ганглиях вегетативной системы, и курареподобными веществами (411].
Это можно объяснить следующим образом. Барбитураты уменьшают эффективность влияния электрошока на секрецию норадреналина и адреналина, снижая возбудимость вегетативных центров. Ганглиоблокирующие вещества устраняют приток импульсов из возбужденных симпатических центров, возбуждаемых ганглионарных клеток и из мозгового вещества надпочечников. Введение
ИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ

Фиг. 45. Влияние повторного электрошока на вегетативное равновесие i[360].
Для каждого животного, помещенного в стандартные условия аноксии, вычерчивали кривую содержания сахара в крови. Затем из области гипергликемии вычитали область гипогликемии и разницу наносили на график. Положительные величины указывают на преобладание симпато-адреналовой системы, отрицательные—на преобладание ваго-инсулнновоГГ системы. После повторных воздействии электрошока степень преобладания симпато-адреналовой системы возрастает, а в процессе восстановления происходит извращение эффекта.
сукцинилхолина или других курареподобных веществ снижает степень повышения концентрации норадренали-на при электрошоке; это связывают с уменьшением судорожной активности [411, 934], что, однако, ие объясняет механизма, лежащего в основе данного явления. Необходимо напомнить о мощном стимулирующем влиянии проприоцепторов на гипоталамус, которое снижается в результате расслабления мышц под действием курареподобных веществ. Итак, повышение содержания
ГЛАВА XXII
норадреналина и адреналина в крови, вызванное электрошоком, подавляется непосредственным депрессирующим действием на вегетативные центры (барбитураты), снижением активирующего действия на них проприоцепто-ров (курареподобные вещества) или блокадой симпатических импульсов.
Если мнение о том, что электрошок действует на центры симпатической системы, можно считать общепринятым (об этом красноречиво свидетельствует повышение содержания адреналина и норадреналина в плазме и их выведение с мочой [852]), то по вопросу о связи этих физиологических реакций с терапевтическим эффектом все еще существуют разногласия. Вайль-Мал-герб [934] склонен связывать терапевтический эффект электрошока с его действием на симпатическую систему; другие авторы [431] придерживаются противоположного мнения, ссылаясь на то, что после введения барбитуратов и сукцинилхолина электрошок продолжает оказывать терапевтический эффект.
Вегетативные симптомы при подпороговом раздражении
При состояниях тревоги, психосоматических расстройствах и на ранних стадиях шизофрении с некоторым успехом применяется подпороговое электрическое раздражение [58, 470, 877]. При этом возникают такие вегетативные симптомы, как гиперемия лица, слюноотделение, слезотечение, расширение зрачков, гипергликемия, а также эмоциональные реакции в виде гнева и страха. Эти симптомы, сопровождающиеся эозииопонией, свидетельствуют о вовлечении гипоталамуса. Эксперименты, проведенные под нембуталовым наркозом, показали, что при большей частоте раздражения содержание адреналина в плазме повышается, а норадреналина — нет [661].
ВЛИЯНИЕ ИНСУЛИНОВОЙ КОМЫ НА ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ
Инсулиновая кома, так же как и электрошок, вызывает возбуждение симпато-адреналовой системы (см. [296]). Прежде чем обсуждать вопрос о влиянии инсули-
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 459
на на концентрацию адреналина и морадреналина в плазме и на скорость их выведения, необходимо рассмотреть нейтральное действие инсулиновой гипогликемии.
Считается, что гипогликемия повышает активность симпато-адреналовой системы за счет подавляющего влияния на корковые функции '. Только при очень глубокой коме начинает преобладать парасимпатическая система, что проявляется в замедлении сердечного ритма. Однако если принять за критерий секрецию кислоты в желудке, то можно обнаружить при гипогликемии раннюю и позднюю фазы усиленной секреции. Ранняя фаза возникает за счет активации парасимпатического отдела гипоталамуса и устраняется ваготомией, поздняя — связана с секрецией АКТГ передней долей гипофиза под влиянием импульсов со стороны заднего отдела гипоталамуса; поздняя фаза блокируется путем удаления надпочечников [261]. Как и при электрошоке, происходит возбуждение обоих отделов гипоталамуса с преобладанием симпатических влияний в большинстве органов. При гипогликемии повышенная реактивность симпатических центров возникает явно за счет резко возросшего «прессорного» эффекта аноксии [342], действия углекислоты [346] и повышенного внутричерепного давления [965].
Имеются некоторые данные о том, что повышение активности симпатического отдела гипоталамуса обусловлено не только его высвобождением из-под тормозного контроля коры, но и притоком импульсов из некоторых отделов лимбической системы, которая, как известно, необычайно чувствительна к судорожным разрядам. Во время гипогликемии судорожные разряды в коре отсутствуют, но в гиппокампе и миндалевидном ядре они проявляются [894, 905]. Поскольку эти структуры анатомически тесно связаны с гипоталамусом, вполне возможно, * * * *
1 В отдельных случаях нарколепсии с эмоциональными расстройствами гипогликемия уменьшала дремотное состояние и приводила К нормализации ЭЭГ [845]. Возможно, этот необычный эффект связан с высокой реактивностью симпатического отдела гипоталамуса и соответственно усилением гипоталамо-кортикальных разрядов.
460 ГЛАВА XXil
что они играют роль в возникновении усиленных симпатических разрядов -на гипоталамическом уровне.
Наше положение о роли гипоталамуса в реакциях, возникающих при гипогликемии, подтверждается тем, что инсулиновая кома сопровождается усилением выведения 17-гидрокортикостероидов с мочой [627]. Это объясняется повышенной секрецией АКЛТ передней долей гипофиза, что в свою очередь определяется усилением гипоталамо-гипофизарных разрядов.
После инсулиновой комы обычно возникает чувство голода. Учитывая, что это чувство регулируется гипоталамусом (см. гл. IX), а также то, что оно возникает после инсулиновой комы, несмотря на восстановление нормального уровня сахара крови, можно считать, что оно обусловлено длительными разрядами последействия в гипоталамусе ', возникающими в результате затяжного возбуждения симпатического отдела при гипогликемии 2.
Содержание адреналина и норадреналина в крови при лечении инсулиновым шоком
Многочисленные физиологические исследования показали, что инсулиновая гипогликемия вызывает возбуждение симпатических центров, а это в свою очередь приводит к повышению секреторной активности мозгового вещества надпочечников. Возможность перенести эти данные в клинику явствует из того, что после инсулиновой комы у больных наблюдается значительное повышение содержания адреналина в крови и выведение его с мочой [918], тогда как содержание норадреналина в крови остается неизменным [853]. Дальнейшее изучение влияния инсулиновой гипогликемии в клинике [599] вы-
* * * *
' Термин длительные разряды последействия в гипоталамусе отражает состояние стойкой активности после интенсивного раздражения. Такое состояние повышенной активности и реактивности может длиться минутами или часами (см. также [490]).
2 Длительное последействие симпатического происхождения, проявляющееся спонтанными сокращениями третьего века, наблюдается также после введения метразола [333] (см. также [310, 317,
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 461
явило обратные соотношения между этими двумя гормонами: при усиленном выведении адреналина содержание норадреналина понижено [222]. При ежедневном введении инсулина как в небольших дозах (проба на толерантность [222]), так и в более значительных, вызывающих кому [853], степень повышения секреции адреналина постепенно снижалась и на 5-й день подобного эффекта вообще не наблюдали. Сходные явления, отмеченные в опытах на крысах, сопровождались истощением запасов адреналина в мозговом веществе надпочечников [715]. Эти исследования показывают, что при определенных условиях интенсивность выведения адреналина (а, возможно, также и концентрация его в плазме) не может служить надежным критерием изменений, происходящих в вегетативных центрах. Очевидно, клинические симптомы инсулиновой гипогликемии не меняются, если мозговое вещество надпочечников не реагирует на нее.
О взаимодействии гипогликемии и электрошока
Гипогликемия приводит к возбуждению симпатических отделов гипоталамуса и нижележащих вегетативных центров за счет подавляющего действия на кору и высвобождения гипоталамуса из-под ее контроля, а также вследствие судорожной активности в гиппокампе и миндалевидном ядре, распространяющейся на гипоталамус. Электрошок активирует симпатические центры путем непосредственного действия на гипоталамус и продолговатый мозг. Комбинация этух двух терапевтических воздействий приводит к резко выраженному возбуждению симпатической системы, оказывающему глубокое влияние на мозг [415]. У адреналэктомирован-ных крыс введение инсулина вызывает кому, судороги и смерть; в ЭЭГ регистрируются медленные дельта-волны. Состояние комы можно продлить введением небольших количеств глюкозы, не вызывающих гипергликемии; восстановления исходного фона при этом не наблюдается. Если же на животное в коматозном состоянии воздействовать электрошоком, то, несмотря на характерную для комы гипогликемию, происходит нормализация с восстановлением исходной ЭЭГ. Если адреналэктомирован-
ГЛАВА XXII
ные крысы реагируют на электрошок замедлением сердечного ритма, то при действии электрошока на фоне гипогликемии частота сердечных сокращений повышается. По-видимому, при гипогликемии возникает состояние симпатической «настройки», которое приводит к повышению мозгового кровотока и восстановлению функции мозга. Это положение подтверждается результатами недавно проведенных экспериментов, которые показали, что при достаточно высокой скорости перфузии можно обеспечить нормальную активность мозга, даже если применять растворы, не содержащие глюкозы [292].
Нам, однако, кажется, что следует принять во внимание еще один фактор. В свете открытого в 1945 г. факта диффузной активации коры разрядами, идущими от гипоталамуса [671, 672], и обнаруженной в 1949 г. активации, получаемой с ретикулярной формации, можно объяснить поразительную нормализацию состояния животного под влиянием электрошока (хотя бы отчасти) резким усилением гипоталамо-кортикальных разрядов. Недавние эксперименты показали, что у кошки, погруженной в коматозное состояние, раздражение гипоталамуса или ретикулярной формации на несколько секунд восстанавливает позные рефлексы и электрическую активность коры [394].
Сходное, но значительно более длительное влияние электрошока на фоне гипогликемии вызвано, видимо, его более сильным возбуждающим действием на симпатические центры промежуточного мозга. Обе группы экспериментов показывают тесную связь между поведением, характером электроэнцефалограммы и вегетативной системой, а также ярко выраженную роль активации гипоталамуса в состоянии симпатической настройки. (Установлено, кроме того, что вывести животное из коматозного состояния можно путем раздражения периферических нервов [517]. Эффект, очевидно, объясняется активацией ретикуло-гипоталамической системы.)
Сарджент и Слейтер [789] сообщили, что в некоторых случаях хороший терапевтический эффект дает комбинация инсулиновой комы с электрошоком, хотя каждый метод в отдельности неэффективен. Аналогичное (но не идентичное) действие оказывает комбинация с электро-
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 463
шоком лекарственных препаратов, стимулирующих симпатическую систему (см., например, [207]).
ПРОБА С МЕХОЛИЛОМ ПРИ ЭЛЕКТРОШОКЕ И ДРУГИХ ТЕРАПЕВТИЧЕСКИХ ВОЗДЕЙСТВИЯХ
На основании данных о физиологическом действии электрошока было высказано предположение, что лечебный эффект зависит от возбуждения симпатического отдела гипоталамуса [360]. Естественно поэтому ожидать у больных сдвига реакций на мехолил и перехода из группы III в группу II и из группы II в группу I. Если же больной до лечения принадлежал к группе I, то у него должна возникнуть значительно более выраженная симпатическая реакция с перерегулированием.
Те несколько случаев, в которых до и после применения электрошока была проведена проба с мехолилом, подтвердили правильность этого предположения [14, 272, 865].
Согласно нашей гипотезе, мы предполагаем далее, что сдвиг реакции на мехолил и переход из группы III в группу II должен сопровождаться улучшением состояния больного, так как симпатическая реакция гипоталамуса нормализуется. Напротив, у больных, относящихся к группам II и I, по мере применения электрошока отклонение от нормы должно усиливаться; следовательно, улучшения не наступает и даже может произойти ухудшение состояния. Поскольку мы исходили из предположения, что центральную роль в возникновении функциональных психозов играет гипоталамус и что его состояние, определяемое при помощи пробы с мехолилом, является направляющим фактором для терапии, необходимо дать оценку имеющимся в настоящее время, хотя и неполным, данным по этому вопросу.
Фанкенштейн и др. [272] продемонстрировали влияние электрошока на результат пробы с мехолилом в 5 случаях с регистрацией состояния до и после лечения. По результатам пробы с мехолилом трое больных сначала были отнесены к группе III, а затем параллельно с улучшением клинической картины их можно было отнести к группе II; у двух больных, относящихся к группе I,
ГЛАВА XXII
характер реакции на мехолил после электрошока не изменился. При этом у одного больного и клиническая картина осталась без изменений, тогда как у другого невротическое состояние тревоги усугубилось. В последнем случае симпатическая реакция была выражена сильнее, чем до лечения.
Александер [14] поставил пробу с мехолилом в 2 случаях «шизофренического психоза». До лечения пробы выявили принадлежность к группе III; в одном случае произошло выздоровление и сдвиг реакции с переходом в группу II; в другом случае улучшения не наблюдалось, состояние тревоги усилилось, а проба с мехолилом показала принадлежность к группе I. Александер сообщает также о случае «навязчивого состояния с шизо-аффек-тивными проявлениями», при котором до лечения реакция на мехолил была типичной для группы I, а после электрошока, не сопровождавшегося судорогами, типичной для группы III. При этом навязчивое состояние и тревога уступили место депрессии. Затем вновь произошел сдвиг симпатической реактивности, одновременно исчезла депрессия и до некоторой степени вернулось состояние тревоги.
Бернштейн [60], хотя он не приводит кривых, полученных при введении мехолила, утверждает, что электро-шоковая терапия дает благоприятный эффект у тех боль-пых, которые до лечения по характеру реакции на мехолил принадлежали к группе III; у больных, принадлежащих к группе I, лечение не вызывало изменений ни в клинической картине, ни в состоянии вегетативной системы.
Наконец, Штеммерман и Оуэн [865] в обширной серии наблюдений с применением пробы с мехолилом установили, что в подавляющем большинстве случаев изменения клинической картины и реакции на мехолил протекают параллельно. Особенно интересно, что даже незначительные ухудшения сопровождались соответствующими изменениями реакции на мехолил.
Наиболее подробное исследование влияния повторной инсулиновой комы провел Гольд [383а]; у 20 больных с кататонической формой шизофрении он наблюдал после лечения усиление симпатической реакции на ме-
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 465
холил и одновременно клиническое улучшение. Алексан-дер [14] представил данные, подтверждающие наличие возбуждающего влияния инсулиновой комы на симпатические центры, а также его связь со сдвигом реакции на мехолил и переходом из группы III в группу II; одновременно наблюдалось улучшение клинического состояния. Кроме того, отмечено, что в некоторых случаях под влиянием инсулиновой комы имеет место переход из группы I (по мехолилу) в группу II при одновременном улучшении состояния больного [865]. Механизм такого парадоксального снижения симпатической реактивности после многократно повторяющейся инсулиновой комы не совсем понятен; возможно, это результат истощения симпатических центров. Такое объяснение подтверждается упомянутыми выше данными об уменьшении секреции адреналина мозговым веществом надпочечников при повторных коматозных состояниях. Более того, повышение концентрации адреналина в крови может способствовать парасимпатической «настройке» гипоталамуса, как это отмечалось в опытах на кошках, описанных в гл. V. Фанкенштейн и др. [272] наблюдали, что сдвиг реакции на мехолил независимо от того, возникал ли он спонтанно или как результат физических или психических воздействий терапевтического характера, сопровождался изменением психического состояния. Это подтверждали и другие упомянутые выше авторы. Так, у больных с психосоматическими расстройствами и повышенной симпатической реактивностью (группа I по мехолилу) переход в группу III (либо возникающий спонтанно, либо обусловленный психотерапией или применением успокоителей) сопровождался депрессией и исчезновением психосоматических симптомов. При исчезновении депрессивного состояния снова появлялись эти симптомы и повышалась реактивность вегетативной системы. Имея в виду данные о том, что различные реакции на мехолил наблюдаются не только у больных, но и у здоровых людей при различных психических состояниях (от тревоги до враждебности) и что они зависят от реактивности гипоталамуса, можно, видимо, заключить, что по крайней мере грубые физиологические и патологические сдвиги в эмоциональной сфере могут сопро-
ГЛАВА XXII
вождаться вполне измеримой реакцией симпатической системы, показателем которой служит проба с мехоли-лом. С помощью той же пробы можно обнаружить также эмоциональные расстройства у больных с хроническими заболеваниями (не психического характера) [866].
ЭЛЕКТРОШОК И ПСИХИЧЕСКОЕ СОСТОЯНИЕ
I.Отсутствие контакта со средой; отсутствие напряженности. П. Настроение изменяется от неудовольствия до тревоги; беспокойство, негативизм,… III. Быстрая смена настроения от депрессии к эйфории (недовольство — смех). … IV. Повышенная напряженность, повышенная активность, экстро-вертированное состояние; настроение: от эйфории до…Ш
Глава ххп
ком долго и при этом не возникает состояния симпатической системы, близкого к состоянию V по Плугу.
Существует еще одно условие эффективности электрошоковой терапии и аналогичных воздействий. При хронической форме шизофрении часто наблюдается пониженная реактивность симпатической системы в условиях различного рода стресса [488]; по реакции на мехо-лил они обычно относятся к группе III. Тем не менее у этих больных электрошок обычно не дает эффекта: не наблюдается ни изменений психического состояния с установлением равновесия в вегетативной системе, ни сдвига реакции на мехолил. На этом основании можно предположить, что в этих случаях выздоровлению препятствует некоторая степень «фиксации» состояния систем гипоталамуса .Этим вызваны, очевидно, и разногласия между разными авторами по вопросу о прогностическом значении пробы с мехолилом [237, 840]. У больных с хронической формой шизофрении и инволюционными психозами реактивность симпатической системы, как правило, понижена (группа III по мехолилу). Однако у больных с инволюционными психозами' наблюдается нормализация клинического состояния и реакций вегетативной системы (переход в группу II), тогда как при хронической шизофрении то и другое отсутствует. Предположение о том, что равновесие в вегетативной системе определяет психическое состояние личности, причем последнее изменяется одновременно с первым {272], не опровергается неэффективностью различных воздействий в отношении этого равновесия.
Хотя причина возникновения выраженных медленных сдвигов, приведенных на фиг. 46, неизвестна, создается впечатление, что они сходны с более быстрыми изменениями кровяного давления и сердечного ритма, возникающими при раздражении гипоталамуса электрическим током [317] или стимуляторами симпатической системы типа метразола [333]. Более того, суточный ритм (см. гл. XVIII) и периодические колебания с большими интервалами (см. гл. XXIII) также связаны с функциями гипоталамуса.
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 469
МЕХАНИЗМ ЛЕЧЕНИЯ УГЛЕКИСЛЫМ ГАЗОМ
В терапии состояний тревоги пользуются 30% -ной смесью углекислого газа с кислородом. Больному рекомендуется сделать 25—30 вдохов. Обычно вдыхание смеси продолжают до момента потери сознания, но в некоторых случаях вызывают лишь субкоматозное состояние. При вдыхании СОг потере сознания предшествует состояние двигательного и сенсорного возбуждения [637].
Наблюдения, проведенные на людях, вдыхавших газовую смесь- различного состава, показали, что при содержании в смеси 5—6% СОг первые патологические симптомы — головокружение, тошнота и одышка — появляются лишь через несколько часов; при повышении концентрации СОг до 12% и выше уже через несколько минут возникают сонливость и ступор [113, 114]. В ЭЭГ во время ступорозного состояния регистрируются медленные дельта-волны (1—2 в 1 сек).
Систематические исследования на кошках под бар-битуратным наркозом показали, что влияние низких концентраций С02 (5—10%) состоит в значительном повышении возбудимости гипоталамуса и реактивности ги-поталамо-кортикалыюй системы. Об этом свидетельствует усиление прессорной реакции при электрическом раздражении гипоталамуса и усиление ответов коры и гипоталамуса на проприоцептивные и' болевые раздражения.- Порог генерализованного возбуждающего влияния на кору при этих воздействиях понижен, а в ответ на сверхпороговое раздражение возникает более интенсивный и длительный процесс возбуждения. Это можно наблюдать и в норме, и при действии стрихнина (302,303).
Если активность гипоталамо-кортикальной системы при действии низких концентраций СОг возрастает, то реакция коры на импульсацию, поступающую по специфической афферентной системе, снижается. При световом раздражении критическая частота слияния снижается, а латентный период депрессии альфа-ритма повышается [796]. Более того, если лишить кору притока афферентных импульсов, вдыхание 10% СОг приводит к уменьшению частоты стрихнинных разрядов, тогда как
№
Глава ххп
при интактной коре, воспринимающей импульсацию из гипоталамуса, частота стрихнинных разрядов в аналогичных условиях повышается [339].
На основании этих экспериментальных данных можно сделать вывод, что относительно низкие концентрации СОг оказывают стимулирующее действие на симпатическую систему и подавляющее — на активность коры. Влияние на кровяное давление осуществляется отчасти рефлекторным путем — через хеморецепторы сино-аор-гальной зоны, а отчасти путем 'непосредственного возбуждения вегетативных центров. Подтверждением этого служит 'сохранение влияния С02 на кровяное давление после сино-аортальной денервации [351], а также сохранение возбуждающего влияния гипер.капнии на ретику-ло-:гИ'Поталамическую систему, изолированную от нервных влияний 'перерезкой мозга на разных уровнях — между воронкой и маммилярнымя телами и кпереди от места вхождения V пары нервов [77]. О сдвиге возбудимости гипоталамуса свидетельствуют изменения электрической активности, усиление прессорной реакции на электрическое раздражение и усиление секреции АКТГ, вызываемой с гипоталамуса, что проявляется химическими изменениями коры надпочечников и лимфопенией [796]. В дозах, вызывающих состояние, близкое к коматозному, СОг оказывает на симпатические центры возбуждающее влияние, сходное с влиянием электрошока и иН'Сулиновой гипогликемии, хотя механизмы всех этих влияний обнаруживают как количественные, так и качественные различия.
Изучение физиологических механизмов терапии коматозными дозами СОг можно проводить на животных, используя для вдыхания смесь газов, содержащую 30— 40% СОг. В этих условиях на наркотизированной кошке можно наблюдать, что короткая фаза возбуждения сменяется резким снижением возбудимости коры и гипоталамуса. Oi6 этом свидетельствуют понижение частоты потенциалов гипоталамуса, уменьшение амплитуды фоновых потенциалов норы и отсутствие ответа со стороны коры и гипоталамуса на проприолептивные и даже сильные болевые раздражения. Необходимо отметить, что, в то время .как реактивность гипоталамо-кортикаль-
БИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ
ной системы в отношении сильных болевых раздражений резко падает, реактивность специфической афферентной системы не изменяется, и кора .продолжает реагировать и отвечает на звуковые и световые раздражения. Необходимо далее подчеркнуть, что при возобновлении дыхания обычным воздухом может наблюдаться феномен «отдачи» — характер потенциалов гипоталамуса и :Коры свидетельствует о более высоком, чем до пробы с СОг, уровне возбуждения [302, 339].
Преобладание возбудительной или тормозной фазы зависит, видимо, от возбудимости центральной нервной системы, длительности действия СОг и его концентрации. Больные в состоянии кататонии пробуждались при вдыхании 25% СОг, но снова впадали в ступор по снятии маски аппарата {591]. (Такого рода действие напоминает эффект, наблюдавшийся на больных с длительной инсулиновой комой, которую .можно устранить с помощью высоких концентраций С02 [950].) При более длительном воздействии СОг стимулирующий эффект появлялся в момент перехода к вдыханию воздуха [184]. Эти явления возникают, видимо, в результате раздражения типоталамо-кортикальной системы —либо непосредственного, либо по механизму «отдачи», которая обычно возникает после аноксии и, в меньшей степени, после вдыхания СОг в высокой концентрации [340]. Некоторые психиатры попользовали стимулирующее действие низких концентраций СОг на симпатическую систему [950] для лечения депрессивных состояний, однако в большинстве клиник предпочитают использовать СОг в наркотических дозах, согласно рекомендации Медуиа [637]. Поскольку при вдыхании смеси с высоким содержанием СОг реактивность гипоталамуса резко снижается, можно думать, что это воздействие должно быть особенно эффективным у больных с высокой реактивностью симпатического отдела гипоталамуса. И действительно, Мориарти [664] показал, что это лечение дает максимальный эффект у людей с повышенной активностью симпатической системы (группа I на основании пробы с мехолилом) и минимальный — у людей с пониженной активностью симпатической системы (группа III); больные, относящиеся .к группе II, занимают в
ГЛАВА XXII
этом отношении промежуточное положение. Однако, несмотря на убедительность .полученных данных, для объяснения этих исследований с физиологических позиций нужно знать характер реакций на мехолил после лечения.
СДВИГ ВЕГЕТАТИВНОГО РАВНОВЕСИЯ В РЕЗУЛЬТАТЕ ЛЕЧЕНИЯ
Предшествующие рассуждения и более детальный анализ механизмов, лежащих в основе шоковой терапии [296, 300, 306, 308,(313], .показали, что электрошок, инсу-линовая кома и низкие концентрации углекислого газа относятся к числу воздействий, вызывающих сдвиг равновесия в вегетативной системе в сторону преобладания симпатического отдела. Сходный '.механизм лежит в основе терапевтического эффекта ацетилхолина и гистамина (реже применяемых в клинике [195 и 245]), поскольку в экспериментах на животных при этих воздействиях возникает повышение реактивности симпатического отдела гипоталамуса вследствие высвобождения его из-под сдерживающего влияния тонической активности ба-рорецепторов сино-аортальной области [310]. Высокие дозы С02, .наоборот, вызывают одвиг типоталамичеокого равновесия в сторону преобладания парасимпатического отдела или же уменьшают реактивность симпатического отдела. Поскольку установлено, что количественные и качественные характеристики гипоталамо-кортикальных разрядов зависят от состояния гипоталамуса, предполагается, что главными факторами, определяющими терапевтический эффект, являются восстановление гипотала-мических функций и соответствующие сдвиги типотала-мо-кортикальных взаимоотношений.
Хорошим показателем изменений внутренней среды организма при лечении шоком является определение содержания адреналина и норадреналииа в плазме и моче. Повышение содержания в крови обоих этих веществ при повторных сеансах электрошока и только одного адреналина при лечении инсулинавой комой, возможно, лежит б основе различий в клиническом эффекте этих двух воздействий. Однако не надо забывать, что содержание адреналина и норадреналииа в плазме и моче не дает
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИЙ 473
полного .представления о степени возбуждения симпатических центров; отсюда ясно, что модификации электрошоковой терапии, влияющие на содержание этих веществ в различной степени, могут тем не менее давать сходный терапевтический эффект [431].
Не лишне обсудить и другие механизмы, вызывающие сдвиги равновесия в вегетативной системе менее прямым 'путем. В наших экспериментальных исследованиях на кошках было показано, что активность гипоталамуса подвергается резким колебаниям вегетативного равновесия, которые коррелируют с ранее описанными изменениями настроения и психического состояния при электрошоке [310, 317]. Кроме того, было показано, что раздражение симпатического отдела /гипоталамуса, которое при контрольных условиях вызывает стойкое повышение кровяного давления и ускорение сердечного ритма, оказывается неспособным вызвать дальнейший симпатический сдвиг в условиях сильного возбуждения симпатического характера. Наоборот, оно вызывает изменения кровяного давления и сердечного ритма, характерные для преобладания парасимпатического отдела и сопровождающиеся понижением реактивности симпатической системы (см. [310]). Очевидно, воздействие, стимулирующее симпатическую систему, например электрошок, примененное на фоне повышенной реактивности этой системы, вызывает за счет появления таких «извращенных» реакций восстановление нормального вегетативного равновесия (см. [717]). Необходимо, однако, напомнить, что при этих условиях всегда остается опасность усугубления нарушенного равновесия.
Факт появления извращенных гипоталамических реакций вегетативного характера при увеличении концентрации адреналина [324] можно использовать для объяснения описанного выше парадоксального снижения симпатической реактивности в результате инсулино-вой комы (см. [108]).
ИЗУЧЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ В СВЯЗИ С ШОКОВОЙ ТЕРАПИЕЙ
Обсудив физиологические механизмы, лежащие в основе различных модификаций шоковой терапии, и уста-
ГЛАВА ХХИ
новив, что электрошок, инсулин-овал гипогликемия и другие аналогичные .воздействия возбуждают симпатический отдел вегетативной системы .на уровне гипоталамуса, .перейдем к рассмотрению других изменений, которые могут возникать в центральной нервной системе при этих условиях. Учитывая существование гипотала-мо-кортикальных -разрядов и роль.гипоталамуса в условных реакциях, интересно изучить влияние шоковой терапии и а условные рефлексы.
В экспериментах на крысах вырабатывали простую условнорефлскторную реакцию избегания. Условные раздражители (звук и свет) сопровождались безусловным подкреплением —приложением электрического тока к решетке, на которой находилась крыса. После соответствующего числа сочетаний крыса в ответ на предъявление условного сигнала перепрыгивала из клетки А с заряженным полом в клетку Б. Затем выработанный условный рефлекс подвергали угашению. Самопроизвольное восстановление варьировало в наших экспериментах от 0 до 20%. Если же при этом применяли такие воздействия, как электрошок, метразол, вызывающий судороги, или иисулиновую кому, то возникало стойкое восстановление предварительно угашенного рефлекса [295, 297, 298, 355]. При применении инсулина в разных концентрациях оказалось, что инсулиновая гипогликемия, при водящая лишь ,к подавлению рефлексов и судорогам, не оказывает влияния на условные рефлексы, тогда как инсулиновая кома восстанавливает их поразительным образом (фиг. 47). Отмечена строгая корреляция между эффективностью разных форм инсулинотерапии и восстановлением предварительно угашенных условных рефлексов. На основании нашего анализа влияния разных форм шоковой терапии на гипоталамус создается впечатление, что восстановление предварительно угашенного рефлекса возникает за счет увеличения возбуждения симпатического отдела гипоталамуса и усиления ги-поталамо-кортикальных разрядов [407, 423].
Это объяснение подтверждается результатами дальнейшей экспериментальной работы. Введение больших доз атропина и тироксина, повышающих реактивность симпатических центров, приводит .к восстановлению уга-

Фиг. 47. Влияние лнсулиновой комы на восстановление трех угашенных условных рефлексов
у крысы [297]. Вертикальные столбики указывают на число сочетаний, потребовавшееся для получения условной реакции в 80—100% проб. Затем условные реакции угашали и вызывали инсулиновую кому, которая приводила к восстановлению условных рефлексов в разной степени.
ГЛАВА XXII
шенных условных рефлексов или усиливает соответствующий эффект электрошока. Пентотал, напротив, снижает реактивность симпатического отдела гипоталамуса [304] и ослабляет центральный эффект электрошока. С нашей гипотезой согласуется и то, что в условиях барбитуратного наркоза угашенные условные рефлексы не восстанавливаются и электрошоковая терапия оказывается неэффективной. Теоретически у нас не было оснований считать, что решающая роль при этом (Принадлежит секреторной активности мозгового вещества надпочечников, и, действительно, введение адреналина не привело к восстановлению угашенных условных рефлексов. Более того, инсулиновая кома продолжает оказывать действие и после адреналэктомии [299]. Эксперименты показали, что за восстановление угашенных рефлексов при шоковой терапии ответственно усиление возбуждения симпатических центров, а не повышение секреторной активности мозгового вещества надпочечников, по которой это возбуждение обнаруживается. Поскольку тор'можение играет существенную роль в услов-норефлекторной деятельности и процессах обучения вообще, становится понятным, что шоковая терапия может изменить поведение.
Электрошок и другие воздействия, вызывающие судороги, применялись в наших экспериментах всего по нескольку раз. В этих условиях предварительно угашенные рефлексы восстанавливаются, а незаторможенные не нарушаются. Розен и Гент [764] в своих экспериментах применяли электрошок и метразол по 10 и более раз. При этом ослаблялись предварительно выработанные условные рефлексы и восстанавливались угашенные, хотя степень восстановления была значительно ниже, чем в наших экспериментах. В связи с этими наблюдениями возникает вопрос о том, является ли ослабление и растормаживание условных реакций результатом повреждения мозга (так как после частых судорог наблюдались кровоизлияния [20]) или изменения его функции.
Ответ на этот вопрос в известной мере можно получить при изучении условной эмоциональной реакции (УЭР). Крыс обучали нажимать на рычаг, давая им в ка-
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 477
честве подкрепления воду. Выработав стабильные реакции, несколько раз применяли щелчки, сопровождавшиеся электрическим раздражением. В результате возникала УЭР: животные перестали нажимать на рычаг при предъявлении условного раздражителя; они становились неподвижными (как бы «застывали»), и у них начиналась непроизвольная дефекация. Эта эмоциональная реакния ослаблялась или исчезала после нескольких сеансов электрошока [495]. Бели отдельные сеансы электрошока были разделены короткими (от 1 сек до 30 мин) или длинными (от 48 до 72 час) интервалами, то их влияние на УЭР было незначительным, при средних интервалах оно резко возрастало [92]; это свидетельствует о возможной роли в этом процессе длительного последействия. Было отмечено далее, что у крыс УЭР, исчезнувшая в результате 10 .последовательных сеансов электрошока, восстанавливается спустя 30 дней. Ясно, что временное исчезновение этой реакции не связано с травмой мозга. Если же после действия электрошока провести угашение условного рефлекса, то восстановления УЭР через 30 дней не происходит [496]. Учитывая усиление возбуждения симпатического отдела гипоталамуса в результате шокового воздействия, можно считать, что сильное возбуждение гипоталамуса препятствует проявлению условной реакции страха, но она восстанавливается после того, как реактивность вегетативных центров возвращается к исходному уровню. Оказалось, что восстановление УЭР и реактивности симпатических центров в описанных экспериментах происходит в один и тот же период времени.
В пользу предположения о том, что исчезновение УЭР (см. также [293]) обусловлено резким возбуждением симпатической системы гипоталамуса, свидетельствуют также результаты применения хлорпромазина; подавляя активность мозга, хлорпромазин способствует восстановлению ослабленной или исчезнувшей под влиянием электрошока УЭР [438]. Далее было обнаружено, что предварительно выработанная УЭР ослабляется в условиях эмоциональной гиперреактивности, вызванной повреждением перегородки, ведущим к нарушению гип-покампально-гипоталамических разрядов [94]. Подобно-
ГЛАВА XXII
го ослабления, однако, не наблюдается при разрушении близлежащих структур, таких, например, как поясная извилина. Отсюда следует, что повышение эмоциональной реактивности, вызываемое различными способами, сопровождается ослаблением или исчезновением некоторых предварительно выработанных условных реакций.
КРИТИЧЕСКИЕ ЗАМЕЧАНИЯ
I. Следует, ложалуй, рассмотреть некоторые критические замечания по поводу теории, постулирующей центральный симпатический механизм действия шоковой терапии. Некоторые авторы [467] утверждают, что если бы это было так, то введение адреналина при лечении психических заболеваний должно было бы давать такой же эффект, как и электрошок. Однако нет оснований ставить знак равенства между возбуждением симпатического отдела гипоталамуса и введением адреналина. В основе изменения поведения и лечебного эффекта инсули-новой комы и электрошока лежит, по-видимому, длительное усиление гипоталамо-кортикальных разрядов, изменяющих условные рефлексы и вызывающих сдвиг равновесия в вегетативных центрах в сторону резкого преобладания симпатической системы. Введение же адреналина, наоборот, приводит через барорецепторные рефлексы к снижению реактивности симпатического отдела гипоталамуса [165, 327]. Более того, изменения поведения под влиянием инсулиновой комы (восстановление заторможенных условных рефлексов) наблюдается не только в норме, но и у адреналэктомированных животных [299], а электрошок остается эффективным у больных с депрессивным состоянием после адреналэкто-мии [417]. Очевидно, что секреторная активность мозгового вещества надпочечников — это одно из многих проявлений возбуждения симпатических центров, и поэтому мысль об идентичности этих процессов противоречит логике и физиологии.
Те же соображения можно привести для объяснения того факта, что именно электрошок, а не введение АКТГ вызывает улучшение у психически больных, хотя АКТГ
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 479
сильнее активирует кору надпочечников, чем электрошок [144].
II. В гл. V -было показано, что снижение кровяного давления введением ацетилхолииа, мехолила и гистами-на приводит к возбуждению заднего отдела гипоталамуса (в результате высвобождения .из-под 'подавляющего влияния барорецепторов), что проявляется в усилении гипоталамо-кортикальных разрядов. Поэтому внутривенное введение гистамина [784] и ацетилхолииа [245] психически больным можно 'рассматривать как одну из форм шоковой терапии, вызывающей такие же гипотала-мо-кортикалыные изменения, как электрошок и инсули-новая кома, но не непосредственно, а рефлекторным путем1. Неудивительно поэтому, что внутривенная инъекция ацетилхолииа вызывает улучшение, а внутримышечное введение ингибитора холинэстеразы ДФП, вызывающего, вероятно, повышение концентрации ацетилхолииа в тканях, углубляет психоз [778]. В экапериментах с ДФП симпатические центры не освобождаются от 'сдерживающего влияния барорецепторов, поскольку кровяное давление изменяется лишь незначительно и очень постепенно «а протяжении нескольких дней.
III. Одна из основных проблем данной книги — это оценка возбудимости гипоталамуса с помощью так называемых «вегетативных тестов». Наше исследование на кошках показало (если ограничить анализ только данными пробы с мехолилом), что область гипотонии увеличивается с уменьшением возбудимости гипоталамуса [310]. Хотя известно, что на л робу с мехолилом оказывают влияние нижележащие симпатические центры, и в особенности центры 'Продолговатого .мозга, это можно не принимать во внимание по следующим причинам: 1) при сравнении здоровых людей с психически .больными ясно, что различия между этими двумя труппами определяются не на уровне .продолговатого мозга; 2) колебания
* * * *
1 Из этого рассуждения о механизмах не следует делать вывод " равной эффективности указанных и классических шоковых воздействий. Но для проведения объективного сравнения необходимо знать, какова длительность сдвига г.ипоталамического равновесия.
480 ГЛАВА XXII
реакции на мехолил, наблюдающиеся у одного и того же человека при различных эмоциональных состояниях, можно сравнить too сдвигами у животных в эксперименте при изменении реактивности гипоталамуса.
Поскольку непосредственное определение реактивности гипоталамуса представляет трудности, можно использовать для этой цели косвенные .показатели, такие, как концентрация адреналина и норадреналнна в плазме или содержание их в моче. Можно определять также выведение стероидов, поскольку известно, что различные виды стресса усиливают симпатические разряды гипоталамуса и активируют гипоталамо-гипофизарную систему. Общеизвестно, однако, что отсутствие изменений по выбранному индикатору может свидетельствовать лишь о несоответствии индикатора, а вовсе не об отсутствии изменений в состоянии гипоталамуса. Более интенсивное выведение норадреналнна у лиц с малой «областью гипотонии» по сравнению с теми, у которых «область гипотонии» велика (223], подтверждает наше положение о том, что при пробе с мехолилом возбуждение симпатических центров косвенно отражается на величине области гипотонии. Однако отсутствие стимулирующего действия инъекций мехолила на выведение норадреналнна [223] или стероидов [842] не умаляет значения твердо установленного факта, что снижение кровяного давления, вызванное мехолилом, приводит к возбуждению симпатической системы и что характер изменений кровяного давления при этом является показателем реактивности симпатических центров.
Избыточное содержание норадреналнна или адреналина (или того и другого) в плазме и моче может служить доказательством того, что в определенной экспериментальной ситуации высвобождаются значительные количества этих нейрогормонов; отрицательный результат ни в коей мере не исключает возможности выделения больших количеств этих веществ в области синантиче-ских контактов симпатической системы без проникновения в измеримых количествах в кровяное русло. Не возникает также сомнений в отчетливо доказанном симпатическом действии лизергииовой .кислоты, хотя при этом не наблюдается повышения содержания адреналоподоб-
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ШОКОВОЙ ТЕРАПИИ 481
ных веществе плазме [625]. Наконец, следует напомнить, что симпатическая система может активироваться не только в целом, но и по частям. Значительное возбуждение симпатической системы может не сопровождаться повышением секреторной активности мозгового вещества надпочечников {316, 357] м даже активацией гиооталамо-гипофизарного аппарата.
16-517
ГЛАВА XXJU
НЕКОТОРЫЕ ДАННЫЕ
ПСИХОФАРМАКОЛОГИЧЕСКИХ
ИССЛЕДОВАНИЙ
На реактивность систем гипоталамуса можно влиять не только описанными выше шоковыми воздействиями, но и введением различных веществ, изучавшихся последнее время в фармакологических опытах, а также при введении испытуемым и психически больным. Хотя мы располагаем весьма скудными данными по этому вопросу, мы надеемся, что они послужат дополнением к тем экспериментальным данным, которые легли в основу главной идеи этой книги.
ХЛОРПРОМАЗИН КАК ПРИМЕР УСПОКАИВАЮЩЕГО ВЕЩЕСТВА (ТРАНКВИЛИЗАТОРА)
Действие хлорпромазина на гипоталамическуго систему состоит в устранении эмоциональных реакций, возникающих при электрическом раздражении гипоталамуса, а именно реакции ярости у кошки и симптома бегства у кролика [81, 166, 676]. Кроме того, после «ведения хлорпромазина оказываются менее эффективными вещества, вызывающие лихорадочное состояние и, как известно, стимулирующие гипоталамус [157]. Наблюдения показали, что подавление реакции мнимой ярости у декорти-цированной кошки требует меньшей дозы хлорпромазина, чем та, которая необходима для того, чтобы вызвать дремотное состояние у интактного животного [16β]. Под влиянием хлорпромазина снижается действие болевого раздражения на освобождение антидиуретического гормона через супраоптическое ядро и нейрогипофив [190]. Отсутствие прессорного ответа на раздражение гипоталамуса у декортицированной кошки не вызы-
ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ 483
вает большого удивления, поскольку удавень кровяного давления в этих экспериментах был очень низким; интересно, однако, что в этом случае хлорпромазин в дозах заведомо подпороговых для интактного организма, вызывает значительное падение кровяного давления. Наблюдения на людях показали, что хлорпромазин вызывает умеренное снижение кровяного давления у здоровых людей, резкое падение его у больных гипертонией и усиливает гипотензивное действие мехолила.
Хлорпромазин и резерпин блокируют описанные выше гипоталамо-типофизарные разряды, приводящие к секреции АКТГ [193]. Эти вещества оказывают действие, подобное разрушению срединного возвышения гипота-дамуса.
Эмоциональные реакции, играющие важную роль в условных рефлексах, .выработанных на базе подкрепления голодом, жаждой и болью, снимаются хлорпромази-ном; правда, он более эффективен в тех случаях, когда подкреплением служит страх перед болью, а не голод и жажда [884]. Интересно, что среди множества фармакологических шроб, .применяемых для оценки транквилизаторов, наилучшей корреляцией с клинической эффективностью этих веществ отличаются тесты, основанные на типичной эмоциональной реакции (дефекация) [885].
Эти данные подтверждают предположение о том, что хлорпромазин снижает реактивность гипоталамуса и особенно (но не исключительно) его симпатического отдела и вызывает сдвиг равновесия в сторону преобладания парасимпатической системы. Поскольку тонус скелетной мускулатуры зависит от этого «эртотропного» отдела гипоталамуса, мышечное расслабление, опущение век и летаргическое состояние, наблюдаемое у обезьян после введения хлорпромазина, можно считать дополнительными симптомами вовлечения гипоталамуса [166].
Вызванные хлорпромазином изменения ЭЭГ у людей незначительны и характерны для состояния покоя. Улиц с преобладанием альфа-ритма последний становится более четко выраженным и распространяется на лобную область [55]. Тенденция ,к синхронизации корковых потенциалов, обычно наблюдаемая у экспериментальных животных [466], отмечается и у человека, проявляясь 16*
484 ГЛАВА XXIII
повышением чувствительности к гипервентиляции [883]. Замедление потенциалов возникает сначала в коре, а затем в гипоталамусе [329] и ретикулярной формации. Малые дозы хлорпромазина снижают диффузную активацию коры, вызванную электрическим раздражением гипоталамуса и centrum medianum таламуса [534, 892], но почти не действуют на порог раздражения ретикулярной формации [534]. Хлорпромазин блокирует реакцию пробуждения в коре, вызванную болевым раздражением у кошек и кроликов [466, 892], что согласуется с мощным влиянием этого раздражения на гилоталамо-кортикаль-ную систему [59]. Наблюдения Престона [723] показали 'высокую чувствительность миндалевидного ядра к хлор-промазину и подтвердили, что усиленные разряды, возникающие в этом ядре, оказывают тормозящее влияние на гипоталамус. О блокаде эмоциональных реакций хлор-промазином свидетельствует следующий простой опыт: введение этого вещества устраняет реакцию пробуждения в коре кролика, вызываемую обычно «присутствием человека» [279]. Сходное влияние хлорпромазина на ЭЭГ и кожно-гальваничсский рефлекс можно наблюдать у человека [951]. Более того, хлорпромазин заметно снижает «ориентировочную реакцию», т. е. эмоциональную реакцию на новое раздражение, предшествующую формированию условного рефлекса [378].
Понижение реактивности диффузной восходящей активирующей системы, проявляющееся по крайней мере на уровне промежуточного мозга, связано с активацией «тормозной» системы, что выражается появлением реакции вовлечения в коре при раздражении хвостатого ядра [892] или интраламинарных ядер таламуса [659]. Ре-ципрокное действие хлорпромазина на две упомянутые выше системы иллюстрируют фиг. 48 и 49. Болевое раздражение вызывает десинхронизацию корковых потенциалов только до введения хлорпромазина (фиг. 48), а порог возникновения реакции вовлечения при раздражении хвостатого ядра снижается после инъекции этого препарата (фиг. 49).
В заключение нужно отметить, что хлорпромазин ослабляет восходящие и нисходящие гипоталамические разряды. На этой основе возникает значительная син-

Фиг. 48. Реакция коры курарезированной кошки на болевое раздражение до (А) и после (Б) введения хлорпромазина (3 мг/кг) [892].
/ — до болевого раздражения; 2—5 — во время и после раздражения в течение 10 сек. Регистрацию производили в средней левой супрасильвиевой извилине. Реакция пробуждения (на А) удерживалась в течение 80 сек после конца раздражения.
Фиг. 49. Влияние х.порпромазина на порог «веретен» хвостатого ядра
у курарезироранной кошки [892]. Нанесение одиночного электрического удара на хвостатое ядро показано стрелкой, порог возникновения веретен на ЭЭГ в контрольных условиях — 9 о. А, Б — через 8 мин после внутривенного введения хлорпромазина (1 мг,'кг); " — через 3 мин после дополнительной инъекции (0.7 мг/кг). Отмечается постепенное снижение порога при увеличении дозы хлорпромазина. / — левая передняя сигмовидная извилина; 2 — левая краевая извилина; 3 — левая супрасильвиева извилина.
ГЛАВА XX1I1
хронизация корковых потенциалов, снижение реактивности симпатической системы и ослабление эмоциональных реакций, проявляющееся независимо от того, выбраны ли в качестве индикатора потенциалы коры или разного рода поведенческие реакции. Уменьшение возбудимости симпатического отдела гипоталамуса проявляется также в снижении соматической возбудимости: возникает общее расслабление и даже дремотное состояние. Беспокойство и психомоторные реакции становятся менее выраженными. Наблюдается сдвиг равновесия между симпатическим и парасимпатическим отделами гипоталамуса в сторону преобладания парасимпатического отдела и сходный сдвиг между диффузной восходящей активирующей системой, запускаемой высокочастотным раздражением гипоталамуса и ретикулярной формации, и неспецифической тормозной системой, за счет которой возникает реакция вовлечения при одиночном или низкочастотном раздражении таламуса и хвостатого ядра.
Сходное действие оказывает резерпин, однако механизм его действия иной. Решающую роль в действии резерпина, по-видимому, играет -возбуждение парасимпатических центров. В пользу этого объяснения свидетельствует индукция сна при стереотипных движениях, как в экспериментах Гесса с раздражением таламуса, а также возникновение миоза [52]. Снижение реактивности симпатических центров также играет при этом важную роль. Именно с ним связано наблюдавшееся у обезьян снижение агрессивности, торможение прессорных рефлексов каротидиого синуса, расслабление третьего века и снижение частоты токов действия ускоряющего нерва сердца [51]. Следующие наблюдения показывают, что центральное действие резерпина связано главным образом с гипоталамусом, а не с ретикулярной формацией:
1. Резерпин не устраняет реакции настороженности, вызываемой раздражением ретикулярной формации [534], но уменьшает црессорную реакцию гипоталамического происхождения [430].
. 2. Гистологические исследования свидетельствуют о повышенной активности супра оптического и паравентри-кулярного ядра гипоталамуса у крысы [645].
ДАННЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИИ 487
Положение о том, что при сдвиге равновесия в гипоталамусе под влиянием резерпина [935] и хлорпромазина [899] (Происходит блокада афферентной импульсации, все еще встречает возражения [487]. Интересно, что резерпин вызывает снижение содержания серотонина в ткани мозга [831], а судороги, вызванные электрошоком и мет-разолом, оказывают противоположное влияние [288].
ФЕНАМИН И ПОДОБНЫЕ ЕМУ ВЕЩЕСТВА
В противоположность транквилизаторам, снижающим реактивность симпатической системы, фенамин и его производные усиливают симпатические, разряды. При их введении повышается кровяное давление, ускоряется ритм сердца, происходит сужение сосудов, расширение зрачков, появляется сухость во рту и т. д. В соответствии с представлениями Гесса об эрготропной системе действие этого вещества состоит в усилении активности соматической нервной системы, что подтверждается повышением двигательной активности человека и животных и облегчением опинальных рефлексов. У здоровых людей психические сдвиги состоят при этом в повышенной настороженности, беспокойстве и эйфории. У человека изменения настроения и настороженность связаны с некоторым повышением частоты альфа-ритма [583] и сдвигом частотного спектра ЭЭГ в сторону высоких частот [372] подобно тому, что наблюдается при сенсорном раздражении или введении кофеина.
В опытах на животных было показано, что реакция пробуждения, вызванная афферентным или электрическим раздражением ретикулярной формации, резко, усиливается [466, 659]. Участие гипоталамической системы становится очевидным из следующих наблюдений:
1. Как в норме [688, 815], так и после декортикации [816] введение фенамина и его производных повышает возбудимость и агрессивность.
2. У крыс с электродами, вживленными в задний отдел гипоталамуса, порог самораздражения снижается при введении фенамина (и повышается при введении хлорпромазина [862]).
ГЛАВА XXIII
3. Эффект внутривенного или внутриартериального введения фенамина, состоящий в смене медленных потенциалов коры быстрой низкоамплитудной активностью (реакция пробуждения), сохраняется после перерезки ствола мозга на уровне четверохолмия (cerveau isole). Если же осуществить перерезку на уровне промежуточного мозга, как раз .перед мамммлярными телами, то фенамин .перестает действовать «а потенциалы коры [459, 594]. Этот опыт несомненно свидетельствует о том, что центральное действие фенамина первично связано с задним отделом гипоталамуса. При оценке реактивности тормозных систем посредством стимуляции интралами-нарных ядер таламуса было показано, что фенамин ослабляет реакцию вовлечения [658, 659].
АНТАГОНИЗМ МЕЖДУ ТРАНКВИЛИЗАТОРАМИ И АНАЛЕПТИКАМИ
Из изложенного ясно, что транквилизаторы и аналеп-тики (на примере фенамина) действуют в противоположных направлениях. Хлорлромазин снижает реактивность симпатического отдела гипоталамуса, а в больших дозах вызывает состояние, близкое к кататонии [153], подобно тому как это имеет место при разрушении заднего отдела гипоталамуса. Фенамин (амфетамин) повышает- реактивность эрготропных систем на уровне гипоталамуса. Неудивительно поэтому, что введение хлорпромазина снимает эффект фенамина и наоборот [84]. Точно так же вызванное резерпином состояние, близкое к кататонии, устраняется стимуляторами симпатической системы центрального действия типа риталина [153].
Антагонизм между транквилизаторами и аналептика-ми проявляется также в их влиянии на условнорефлек-торную деятельность. В экспериментах Брэйди [83] крыс обучали нажимать па рычат, чтобы получить пищу. Применение после этого звукового раздражителя в сочетании с электрическим привело к развитию условнореф-лектор.ной реакции страха: животное перестало нажимать на рычаг при предъявлении условного звукового сигнала без подкрепления. Как показано на фит. 50, фенамин и резерпин (транквилизатор) действуют на эти
ДАННЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИИ 489
реакции в противоположных направлениях. По сравнению с контрольной груишой животных, которым вводили физиологический раствор, число нажимов на рычаг возрастало после фенамина и снижалось при действии резерпина, т. е. эффекты соответствовали уровню возбуждения эрготропной системы. Однако во время условной реакции страха число нажимов на рычаг у животных, получавших фенамин, было меньше, а у получавших резерпин — больше, чем в контрольных условиях. Это свидетельствует, видимо, о том, что реакция страха усиливается фенамином и ослабляется резерпином, что согласуется с результатами многочисленных экспериментов, в которых было обнаружено, что эмоциональная реактивность непосредственно связана с реактивностью симпатического отдела гипоталамуса. Условно-рефлекторные исследования, проведенные на людях, также показали, что фенамин облегчает выработку условных рефлексов, а хлорпромазин оказывает противоположное действие [208, 250].

ПСИХОТРОПНЫЕ ВЕЩЕСТВА
Из числа психотропных веществ лизергиновая кислота (LSD) и мескалин оказывают на центральную нервную систему сходное с фенамином действие. В экспериментах на животных влияние этих
is. so 4S
Время, мин
Фиг. 50. Влияние фенамина (амфетамина) и резерпина на реакцию нажима на рычаг и условную эмоциональную реакцию у крысы [86а].
Прямая стрелка — включение
условного звукового сигнала; ломаная стрелка — выключение условного сигнала и подача короткого безусловного электрического раздражения.
ГЛАВА ХХШ
веществ характеризуется возбуждением симпатических центров, усилением реакции пробуждения, десинхрони-зацией коры и понижением чувствительности интралами-нарн.ых ядер таламуса в смысле возникновения синхронизированной реакции вовлечения. Кроме того, они вызывают улучшение настроения. Берингер [56] описал ощущение счастья, возникающее при введении мескали-на и связанное с характерными изменениями восприятия. Другие авторы наблюдали зрительные галлюцинации и бреды параноидного характера. Производные фенамина вызывают подобный же эффект, и Коннелл [154], который дал обзор литературы по этому вопросу, подчеркивает, что психоз, вызванный фенамином, порой совершенно неотличим от параноидного.
Минимальные дозы (20—30 мг) диэтиламида лизер-гИ'Новой кислоты (сокращенно LSD-25) вызывают изменения психики. Это проявляется в беспокойстве и нервозности при самых малых дозах и в .появлении серьезных психотических симптомов типа галлюцинаций и маний, сноподобных состояний, деперсонализации и т. п.— при несколько больших дозах [504]. По мнению некоторых авторов, последние симптомы напоминают шизофренические реакции. Обычно наблюдаются сдвиги настроения как в сторону эйфории, так и в сторону депрессии [1], нарушения восприятия, касающиеся прежде всего зрительных ощущений, а затем часто слуховых и тактильных. Постоянно отмечаются вегетативные сдвиги. Усиление симпатической активности проявляется в расширении зрачков, ускорении сердечного ритма, повышении давления и понижении температуры кожи [192]. Нужно подчеркнуть, что строгий параллелизм между вегетативными и психическими сдвигами обнаруживается не только при введении исследуемым больших доз LSD-25 [504], но и во время всего периода после инъекции, когда в течение нескольких часов наблюдается постепенное усиление и последующее ослабление симптомов [192]. .
Учитывая тесную связь между настроением и активностью гипоталамуса, а также роль последнего в восприятии, можно высказать предположение, что LSD-25 изменяет функциональную активность гипоталамуса и
!НЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИИ 491
влияет на гипоталамо-кортикальные разряды. Подтверждением этому служат результаты экспериментов на кроликах [684], .показавшие ведущую роль промежуточного мозга при действии LSD-25. В этих экспериментах мы наблюдали не «психический» эффект от введения этого вещества, а его действие на центральную нервную систему, .которое можно оценить объективно. Было показано, что LSD-25 вызывает у кроликов усиление спонтанной активности и .повышение температуры тела. Эффект сохраняется при удалении коры, но фактически полностью снимается после удаления промежуточного мозга.
Эти наблюдения ^свидетельствуют в пользу предположения о том, что LSD вызывает описанные симпатические и соматические эффекты благодаря действию на эрготропный отдел гипоталамуса. Необходимо подчеркнуть, что дополнительные симптомы—тилерглике-мию, пилоэрекцшо и исчезновение медленных потенциалов на ЭЭГ,— наблюдаемые у кролика лри введении этого вещества [772], можно воспроизвести путем стимуляции гипоталамуса, так что они, видимо, являются отражением усиления его восходящих и нисходящих разрядов. С нашей интерпретацией согласуется также появление патологической активности в обонятельном мозге человека после введения LSD-25 [660].
Интересный факт параллелизма между вегетативными и психическими явлениями еще не служит доказательством постулированной нами причинной связи между ними. Но мы располагаем еще одним рядом фактов, подтверждающих наше представление. Церлетти [141] и его сотрудники провели широкое исследование по изучению влияния 18 различных производных LSD-25 на вегетативную нервную систему животных и установлению корреляций с психотической активностью, наблюдающейся у человека. Они обнаружили строгий параллелизм между активностью симпатических центров и действием этого вещества на психику. У человека преобладают психические симптомы, а у животных — таких, например, как кролик,— вегетативные. Это не удивительно и не опровергает представления о том, что «их проявление регулируется общим координационным центром»
M
ГЛАВА XXIII
[770]. Мы действительно убеждены .в том, что «психические» и «вегетативные» эффекты — это разные выражения одного и того же нервного (или гуморального) процесса.
Тот факт, что степень возбуждения симпатических центров оказывает значительное влияние на настроение, можно проиллюстрировать также на примере действия метамфетамина. Это вещество вызывает колебания содержания адреналина -в плазме. При его .повышении наблюдается напряженность и беспокойство, при .понижении, напротив, доминируют расслабление и эйфория [574]. Уиклер [948] и ряд других подчеркивают связь между настроением и .потенциалами ЭЭГ, утверждая, что в общем «десинхронизация» (с увеличением частоты альфа-колебаний или без него) связана с появлением тревоги, галлюцинаций, фантазий и. (или) тремора, а «синхронизация» (с замедлением частоты альфа-ритма или без него) —с эйфорией, расслаблением, успокоением и (или) дремотой.
ДЕЙСТВИЕ ПСИХОТРОПНЫХ ВЕЩЕСТВ И УРОВЕНЬ НОРАДРЕНАЛИНА
Выдвинутое нами основное положение о связи колебаний настроения и психомоторного возбуждения с соответствующими сдвигами возбудимости гипоталамуса и равновесия вегетативных систем подтверждается данными современной биохимии. Основополагающие исследования Фогт [908], показавшие, что норадреналин обнаруживается в стволе мозга (причем в гипоталамусе концентрация его максимальна), привели к заключению о том, что норадреналин представляет собой главный ней-рогормон, концентрация которого зависит от степени возбуждения симпатических центров. Блестящая работа Броди и др. [105] подтвердила это положение на большом материале. За недостатком места и в связи с тем, что в настоящее время эти работы не завершены, мы ограничимся лишь кратким их описанием.
Было показано, что химический предшественник нор-адреналина, 3,4-диоксифенилаланиш (ДОФА), вызывает диффузную активацию ЭЭГ, а также симптомы
НЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ 493
активации эрготропной системы. Вещества, химически близкие норадреналину, в .противоположность последнему растворимы в липоидах, '.проникают через гемато-эн-цефалический барьер и при внутривенном введении вызывают .появление эрготропного синдрома. Это относится к фенамину, LSD, риталину, мератрану, ибогаину, тет-рагидро-р-нафтиламину и другим. Поэтому можно считать, что эти вещества действуют .на области мозга, чувствительные к норадреналину'. Судя по данным Фогт и ранее описанным опытам, к таким областям относится задний гипоталамус.
Более того, такие вещества, как марсилид (ищрониа-зид), препятствующие ферментативному расщеплению норадреналина, оказывают глубокое влияние на поведение. Введение этих веществ .при депрессии вызывает следующие положительные сдвиги: 1) снятие напряженности и депрессии, успокоение; 2) хорошее самочувствие, связанное с ослаблением фобий и т. п.; 3) возбуждение, гнпсрреактивность, бессонница и 4) маниакальные и параноидные симптомы2. Биохимические исследования показали, что ингибиторы фермента моноаминоксидазы приводят к накоплению в мозге ссротонина и норадрена-лина. Однако гиперреактивность симпатической системы связана только с накоплением последнего [105]. Эти данные .представляют решающие доказательства особой роли норадреналина в определении степени возбуждения центров эрготропной системы. Следует, однако, признать, что для окончательного подтверждения предположения о роли норадреналина в регуляции симпатических функций необходимо еще по крайней мере доказать, что активация центров эрготропной системы сопро-* * * *
1 Недавно было показано, что собаки, получавшие в течение Двух недель резерпин, нечувствительны к риталину. Поскольку под влиянием резерпина происходит исчезновение но.радреналина из гипоталамуса [485], можно думать, что центральное возбуждающее Действие риталина зависит от присутствия норадреналина [215].
2 Марсилид снижает успокаивающее действие резерпина н «нейтрализует» действие последнего на содержание в тканях норадреналина [137, 485]. Более того, по своему действию он подобен электрошоку, возбуждающее влияние которого па симпатический отдел гипоталамуса подробно обсуждалось выше.
/
494 ГЛАВА XXIII
вождастся повышением секреции норадреналина и что рассматриваемые нами психотропные вещества не действуют ни на какие другие ферменты, кроме аминоксида-зы [812]'.
ИНТЕРПРЕТАЦИЯ СУЩЕСТВУЮЩИХ ДАННЫХ
В табл. 10 дан краткий обзор общего действия этих веществ на некоторые функции мозга и поведение2. Создается впечатление, что изменения равновесия в гипоталамусе сильно влияют на настроение, эмоциональную реактивность и на общее поведение организма. Индивидуальные различия в характере реакции на эти вещества заставляют предполагать, что состояние гилотала-мического равновесия имеет первостепенное значение. Установлено, что LSD (а также ипрониазид [812]) вызывает не только состояние эйфории, возбуждения, ги-поманиакальное поведение, но также и состояние депрессии. Иногда эти совершенно различные эмоциональные состояния чередуются [140, 141], что, возможно, указывает на колебания гипоталамических состояний, о которых мы неоднократно упоминали. По всей вероятности, эти вещества влияют на оба отдела гипоталамуса, но в норме их влияния на симпатическую часть доминируют. Правильность этого предположения подтверждается тем, что на фоне барбитуратного наркоза, блокирующего сим-патичеокую реактивность гипоталамуса [304], LSD и ибогаин вместо обычного «'симпатического» действия вызывают замедление сердечного ритма и снижение кровяного давления [803]. Установлено, что 'При применении электрошока после предварительного лечения резерпином иногда возникают резко выраженные ларасимпати-
* * а *
1 Было показано, что четыре разных ингибитора аминоксидазь: вызывают сходные химические эффекты [86, 105].
2 Незначительные различия и роль биохимических сдвигов в мозге мы не рассматривали. Касаясь ацетилхолина, следует упомянуть, что кататонический ступор можно вызвать не только разрушением заднего отдела гипоталамуса, но и введением ацетилхолина и ингибиторов холинэстеразы в желудочки мозга и что клиническое состояние облегчается введением холинэстеразы [8281.
ДАННЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИИ 495
ческие разряды [61]; это еще раз свидетельствует о том, что нейротропные вещества представляют собой мощные средства «настройки» гипоталамуса и что резерпин сдвигает вегетативное равновесие в сторону преобладания
Таблиц а 10
НЕЙРОФИЗИОЛОГИЧЕСКИЕ ЭФФЕКТЫ ПСИХОТРОПНЫХ ПРЕПАРАТОВ
| Транквилизаторы | Аналептики, психотропные препараты и ингибиторы ъ:опо-аминоксидазы | |
| Состояние вегетативных центров Центральное вегетативное равновесие Активность диффузной афферентной ретикулярно-гипота-ламической системы Реакция вовлечения при раздражении хвостатого ядра или таламуса Условные рефлексы Корковые потенциалы Настроение Общая активность | Симпатическая реактивность и гипота-ламо-кортикальные разряды снижаются Сдвиг в сторону преобладания парасимпатической системы Понижается Усиливается Ослабляются Синхронизация Подавленное Ослаблена | Симпатическая реактивность и гипота-ламо-кортикальные разряды повышаются Сдвиг в сторону преобладания симпатической системы Повышается Ослабляется Усиливаются Десинхронизация Маниакально-эйфо- ричное, тревожное Усилена |
парасимпатической системы. (Выше говорилось о том, что электрошок действует на оба отдела вегетативной системы.)
Наконец, следует попытаться определить вероятные механизмы, при помощи которых эти вещества изменяют поведение, и сопоставить эти механизмы с чисто физио-
(
496 глава xxikl
логическими. Известно, что циклическое чередование бодрствования и сна связано с изменениями вегетативной реактивности и гипоталамичеекого равновесия. Будем ли мы считать сон активным парасимпатическим процессом [453] или результатом уменьшения активности центра бодрствования, расположенного в заднем отделе гипоталамуса [734], несомненно одно, что переход от со-стояния бодрствования к состоянию сна является рв' зультатом сдвига вегетативного равновесия — в состоянии сна доминирует парасимпатический отдел, а в состоянии бодрствования преобладает задний, симпатический, отдел .гипоталамуса. Активное состояние заднего отдела гипоталамуса, а также тесно связанной с ним ретикулярной формации во время бодрствования вызывает дееинхронизацию потенциалов в коре голов«ого мозга. Прямая или вызванная рефлекторным шутем активация этой системы, связанная с усилением внимания и эмоциями, усиливает дееинхронизацию, свидетельствующую о состоянии возбуждения в коре, и одновременно повышает тонус скелетных мышц. Проприоцептивные разряды и импульсы от ствола мозга через гамма-систему подкрепляют активность эрготролной системы. Настроение на фоне бодрствования колеблется от покоя до напряжения и разнообразных эмоциональных состояний (страх, гнев и т. д.), которые, судя по результатам проб с мехолилом, связаны с различными степенями симпатической активности гипоталамуса. При исследовании реакции вовлечения установлено наличие реципрокных взаимоотношений между тормозной афферентной системой и активностью рстикуло-гипоталамической системы [891]; можно считать, что во время сна реактивность тормозной системы выше, чем в период бодрствования. Соответствующее электрическое раздражение переднего отдела гипоталамуса вызывает сон, связанный с синхронизацией корковых потенциалов [11]. Активация переднего отдела гипоталамуса при нагревании также вызывает синхронизацию в коре, расслабление скелетных мышц и состояние дремоты [915]. При обоих этих воздействиях наблюдается реиипрокное торможение заднего отдела гипоталамуса. Возникновение сна можно объяонить или таким образом, или ка>к результат ослаб-
ДАННЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИИ 497
пения симпатической активности гипоталамуса и ренип-рокного повышения активности его переднего отдела.
Предполагается, что тот же механизм участвует в эмоциональных и психических сдвигах, вызываемых транквилизаторами, аналаптиками и некоторыми другими психотропными препаратами. Диапазон эмоциональных изменений .при этом сильно расширяется, но едва ли можно отрицать сходство между действием транквилизаторов и сдвигом гийоТаламического равновесия в сторону преобладания парасимпатической системы. Данные экспериментов с аналептичеокими и психотропными веществами, а также с ингибиторами аминоксидазы заставляют предполагать, что их действие противоположно действию транквилизаторов, т. е. что они вызывают сдвиг в сторону преобладания симпатического отдела гипоталамуса и усиление гипоталамо-кортикальных раз-
Основной смысл данной гипотезы состоит в признании того, что изменения интенсивности гипоталамо-кортикальных разрядов, отражающиеся в настроении и эмоциональном поведении, определяются состоянием гипоталамуса. Несущественно, чем вызывается подавление симпатического отдела гипоталамуса—соответствующими изменениями в висцеральном мозге, ослаблением афферентных импульсов, как в наших пробах с кураре, повреждением заднего отдела гипоталамуса, применением транквилизатора типа хлорпромазина или же возбуждением переднего отдела гипоталамуса2. В каждом из этих случаев эмоциональная реактивность уменьшает-
' Например, очевидно, что не все фармакологические препараты, способные снижать возбудимость заднего отдела гипоталамуса, можно считать транквилизаторами, пригодными для клинического применения. Систематическое изучение действия этих веществ на ретикулярную формацию, поведенчеокие реакции, обонятельный мозг-[798]. реактивность коры и рекрутирующей системы таламуса наряду с ^клиническими тестами создает научную основу фармакологической классификации. Многочисленные симпозиумы Г530, 540, А«, 730, 731] и особенно исследования Брэдли и Кэй [86], Ганглоф-(ojri" ье f281' 658^ Калама и Кнллама {534], а также Уиклера
игаопкасались этих вопросов.
Подобное действие, очевидно, оказывает резерпин [105].
ГЛАВА XXIII
ся. Этот принцип .применим также к объяснению связи между повышенной симшатичеокой активностью гипоталамуса и эмоциональной гиперреактивностью. Уровень возбудимости гипоталамуса определяет выраженность эмоциональных состояний. Если ретикулярная формация блокирована, а реактивность гипоталамуса снижена введением барбитуратов, активация и эмоциональные реакции исключаются. Этим отличается действие наркотизирующих веществ от действия транквилизаторов.
Очевидно, существуют большие пробелы .в имеющихся у мае сведениях о действии этих веществ. Кроме того, мы умышленно отказались от излишней детализации, чтобы подчеркнуть и разъяснить основные изменения, вызываемые ими. Нам известно, что не все транквилизаторы действуют одинаково [86, 948] и что существуют характерные различия между действием аналептических и психотропных препаратов [219]. Физиологическими [890] и фармакологическими работами [131, 593—595] установлено, что активирующая система сложнее и способна к более дифференцированному действию, чем это показано в нашем анализе. Кроме того, существуют количественные и качественные различия в реакции на транквилизаторы, аналептические и психотропные вещества у разных людей, у одного и того же человека при различных психических состояниях, а также в норме и патологии [131]. Эти наблюдения приводят к мысли о различных состояниях гипоталамической настройки, о вариабельности уровня гипоталамической реактивности и открывают широкие перспективы для дальнейших исследований в этой области.
ДЕЙСТВИЕ ФАРМАКОЛОГИЧЕСКИХ ПРЕПАРАТОВ И УСЛОВНЫЕ РЕФЛЕКСЫ В КЛИНИКЕ
Некоторые аспекты этой проблемы изложены у Алексаадера [15]. Он применил метод условных рефлексов для обследования здоровых людей и больных из психоневрологической клиники. В качестве условных раздражителей применяли звуковые сигналы, безусловным раздражителем служило электрическое раздражение руки, которое вызывало кожно-гальванический реф-
МАННЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ 499
леке, приводящий к снижению сопротивления на ладонях. Регистрировали влияние этих раздражителей на ЭЗГ. Положительные условные раздражители вызывали десииХ'ронизацию в ЭЭГ и отчетливую кожно-гальвани-ческую реакцию. Дифференцировка вырабатывалась посредством устранения подкрепления по типу классических опытов Павлова и выражалась в торможении кож-но-гальванического рефлекса на отрицательный условный раздражитель. Сопоставляя результаты обследования, Александер установил наличие выраженных количественных и качественных различий между нормой и патологией. При депрессии положительные условшые раздражители либо вовсе не вызывали условного рефлекса, либо этот рефлекс был отставленным и ослабленным, кроме того, у таких больных на ЭЭГ наблюдалась гиперсинхронизация потенциалов, а не десинхро-низация, наблюдаемая у здоровых людей. При состояниях тревоги отрицательный условный раздражитель приобретал характерные черты положительного условного раздражителя и вызывал сильную гальваническую реакцию. Эти реакции соответствуют ультрашарадо-ксальной фазе, описанной Павловым.
Чтобы объяснить эти интересные наблюдения, важно учесть изменения условных рефлексов, которые возникают у здоровых людей при применении некоторых фармакологических препаратов. Александер показал, что после введения мепробомата и бенактизина (антифоби-ческий препарат [509]) реакция десинхронизации в ЭЭГ превращается в г.иперсиихро'низацию. Выше мы говорили, что мепробомат 'снижает реактивность гипоталамиче-ской системы, ослабляя афферентные импульсы от мышц. Беиактизин ослабляет эмоции, снижает реакцию настороженности в ЭЭГ и значительно смягчает симптомы экспериментального невроза у подопытных животных [509—511]. Поскольку (реакция настороженности зависит от возбудимости ретикуло-гипоталамической системы,, можно сказать, что снижение возбудимости этой системы является одним из факторов, ведущих к изменению реакции настороженности.
Учитывая то, что было сказано ранее о физиологических механизмах, лежащих в основе экоперименталь-
ГЛАВА XXIII
ных неврозов (гл. X), можно трактовать исследования Александера" (которые необходимо повторить и расширить) как свидетельство зависимости экспериментальных неврозов, невротических и психотических состояний, а также изменений деятельности мозга, вызванных фармакологическими препаратами от сдвигов в настройке гипоталамическои системы. Условные раздражители действуют на возбуждающую и тормозную системы головного мозга, .причем положительные условные раздражители влияют главным образом на возбуждающую, а отрицательные в основном на тормозную систему; связь этих систем соответственно с задним и передним отделами гипоталамуса была отмечена выше.
При возбудительном типе экспериментального невроза дифференцировка исчезает и отрицательные условные раздражители вызывают возбуждение, которое в норме возникает ,под влиянием положительных условных раздражителей. Александер также отметил, что при состояниях тревоги условный рефлекс возникает в ответ на отрицательный условный раздражитель. И наоборот, при депрессии положительный условный раздражитель вызывает реакцию коры (гиперсинхронизацию), характерную в норме для отрицательного условного раздражителя. Можно предположить, что при депрессии равновесие гипоталамическои системы сдвинуто в сторону преобладания парасимпатической системы, а при состояниях тревоги — в сторону преобладания симпатической. Поскольку мепробомат и бенактизин ослабляю-активность ретикуло-гипоталамической системы, возможно через различные механизмы, не приходится удивляться тому, что в экспериментах Александера по выработке условных рефлексов они вызывают у здоровых людей картину, характерную для больных с депрессией. В обоих случаях, очевидно, происходит одинаковый сдвиг равновесия систем гипоталамуса.
Хотя мы еще далеки от понимания этих явлений, для дальнейшей работы в этом направлении полезно запомнить следующее: 1) хлорпромазин снижает симпатическую реактивность гипоталамуса и ослабляет гипотала-мо-кортикальные разряды, тогда как аналептические препараты оказывают противоположное действие;
ДАННЫЕ ПСИХОФАРМАКОЛОГИЧЕСКИХ ИССЛЕДОВАНИИ 501
2) хлорпромазин вызывает сдвиг равновесия систем гипоталамуса в трофотропную сторону, а фенамин — в эрготропную; этим вызывается настройка, при которой реактивность систем гипоталамуса коренным образом изменяется: 3) действие хлорпромазина характеризуется тенденцией к синхронизации, а действие аналептических веществ приводит к преобладанию десинхронизации; 4) транквилизаторы могут ускорить наступление депрессии, а фенамин (или лизергиновая кислота) может вызвать маниакальные и навязчивые состояния. Даже такие вещества, как тетрагидро^-нафтиламин, который в химическом отношении отличен от веществ, обсуждаемых в данной главе, но оказьгвает возбуждающее действие на задний отдел гипоталамуса [907], могут временно превратить депрессивное состояние в маниакальное!
Ни депрессивные состояния, ни состояния возбуждения не являются единообразными'. Тем не менее различные уровни настройки гипоталамуса могут играть существенную роль при этих состояниях.
При ажитироваиной депрессии наблюдается повышение секреции адреналина и норадреналина, тогда как при ступорозной форме психотической депрессии секреция этих нейрогормонов уменьшается [835].
ГЛАВА ЛЛ1 V
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ
И ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
ФИЗИОЛОГИЧЕСКАЯ ОСНОВА ПЕРИОДИЧЕСКИХ ЗАБОЛЕВАНИЙ
Суточный ритм общей активности вегетативной нервной системы и интенсивности секреции АКТГ считается следствием изменений возбудимости промежуточного мозга (гипоталамуса). Кроме того, существуют циклы с более длительным периодом, которые были подробно изучены у грызунов. Наибольшей четкостью отличается 4—5-дневный ритм у самки крысы, связанный с изменениями общей активности и с морфологическими изменениями со стороны влагалища, соответствующими астральному циклу. Доминирующая роль яичников в этом явлении очевидна, так как после их удаления эти циклы прекращаются.
Учитывая зависимость секреции половых и других эндокринных желез от гипофиза, выделение которым различных тройных гормонов в свою очередь регулируется импульсами с гипоталамуса, естественно задать вопрос, в какой мере состоящие гипоталамуса определяет эту циклическую активность. Проведенные в прошлом наблюдения показали, что перерезка ножки гипофиза устраняет физиологический 4—5-дневный ритм у крысы; устанавливался более медленный 9—18-дневный цикл [751]. В то время не было известно, что воротные сосуды гипофиза после такой перерезки регенерируют [427], и последующие изменения ритма могут происходить вследствие частичного восстановления этих сосудов, благодаря чему нейрогормо'ны гипоталамуса снова получают доступ .к 'передней доле гипофиза. Вполне возможно, что стойкая блокада сосудистых и нервных связей между гипоталамусам и гипофизом прекратила бы эти циклы.
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ
5D3
Имеются и другие факты, указывающие на роль гипоталамуса в циклическом поведении. Обнаружено, что сильное эмоциональное .напряжение в сочетании с физической нагрузкой вызывает тяжелые .повреждения гипоталамуса и иногда даже приводит к смерти [754]. У выживавших животных отмечалось выраженное увеличение продолжительности цикла (13—17 дней) с ослаблением общей активности и наступлением стадии диэст-руса, выявляемым путем анализа влагалищных мазков. Кроме того, в яичниках были обнаружены стойкие желтые тела — наблюдалось состояние мнимой беременности, которое вызывается также либо стерильной копуляцией, либо механическим раздражением матки. Очевидно, это состояние было вызвано или рефлекторным путем, или путем эмоционального возбуждения.
Важную роль в циклическом поведении играет также щитовидная железа. Продолжая свои опыты по изучению влияния резекции щитовидной железы, Рихтер и др. [756] вводили крысам радиоактивный иод для подавления активности щитовидной железы (в том числе и дополнительных желез, которые остаются незатронутыми при хирургическом вмешательстве). При этом общая активность снижалась, а длительность циклов увеличивалась до. 14—15 дней. В то же время в мазках, взятых из влагалища, уменьшались в числе или полностью исчезали ороговевшие клетки в течение периодов относительной неактивности. В этом случае опять-таки стойкие желтые тела в яичниках и характер эпителия влагалища напоминали состояние мнимой беременности. Правильность этого предположения была в дальнейшем подтверждена тем, что у нормальных крыс введение пролактина вызывало такие же симптомы, как и введение I131. Более того, оказалось, что при скармливании животным высушенной ткани щитовидной железы восстанавливался нормальный 4-дневный цикл. Как только дачу ткани прекращали, цикл снова становился патологическим.
Бели учесть, что гипофункция щитовидной железы и эмоциональное напряжение вызывают мнимую беременность путем увеличения секреции пролактина передней долей гипофиза и что после перерезки ножки гипофиза стерильная копуляция не вызывает секреции
ГЛАВА XXIV
пролактина, то можно сделать вывод, что влияние эмоционального возбуждения и гипофункции щитовидной железы на секрецию пролактина осуществляется через гипо-таламр-гипофизарные разряды. Рихтер предполагает, что поскольку чрезмерное возбуждение приводит к анатомическим .повреждениям гипоталамуса, в результате этого может происходить высвобождение пролактина. В экспериментах то изучению влияния гипофункции щитовидной железы эти повреждения, возможно, являются «результатом усилий организма принудить небольшой остаток ткани щитовидной железы выделять достаточное количество тироксина для .поддержания гомеостаза».
Значение указанных фактов для проблем, обсуждаэ-мых в данной книге, состоит в том, что они имеют определенное отношение к периодической кататонии, которая, очевидно, связана с колебаниями секреторной активности щитовидной железы. Кроме того, в блестящей работе Рихтера предложен новый метод воздействия на состояние гипоталамуса в интактном организме; речь идет о количественных сдвигах в механизме обратной связи, в которой участвует щитовидная железа и, возможно, другие эндокринные железы, регулируемые трапными гормонами гипофиза, секреция которых в свою очередь .контролируется гипоталамусам.
Из обширной литературы по этому вопросу можно привести несколько примеров, которые могут послужить отправным пунктом для обсуждения вопроса о роли гипоталамуса в патогенезе периодических заболеваний [745, 755]. Основанием для утвердительного ответа на этот вопрос могли бы служить наблюдения, .свидетельствующие о том, что главные симптомы периодических заболеваний обусловлены циклическими парасимпатическими или симпатическими разрядами, а также наличием циклических изменений эндокринной активности. Мы начнем с анализа вегетативных изменений при некоторых периодических заболеваниях, а затем перейдем к анализу эндокринной активности при периодической кататонии.
Периодическое лихорадочное состояние, продолжающееся в течение нескольких дней и возобновляющееся через интервалы, равные примерно 20 дням, в течение
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ 505
многих лет,— это болезнь, связанная с явными симпатическими симптомами. Температура при .подобных состояниях бывает очень высокой и сопровождается потами, тахикардией, тошнотой и рвотой. Очевидно, эти симптомы связаны с симпатическими и парасимпатическими разрядами, причем, судя по температуре тела и частоте сердечных сокращений, преобладают первые. Вместе с тем известно, что лихорадочные периоды чередуются с периодами гипотермии длительностью 5—7 дней, когда температура падает до 32,8°. При этом отмечается тошнота и рвота, что отчетливо указывает на роль парасимпатических разрядов.
Существуют другие проявления парасимпатических разрядов — периодическая рвота у детей и .периодические боли в животе, сопровождающиеся слюнотечением, тошнотой, рвотой и болевыми ощущениями. Левин и Рихтер [567] описали случай выраженной ваготонии (миоз, замедление сердечного ритма) и повышенной реактивности парасимпатической системы, проявляющейся в остановке сердца на 19 сек при надавливании на глазное яблоко (глазо-сердечный рефлекс). Приступы боли продолжались от 3 до 25 час и повторялись через каждые 5—10 дней. Им предшествовали «периоды депрессии, апатии, потери интереса к окружающему, а также усиленное слюноотделение». Во время приступа, при котором боль была настолько сильной, что заставляла больного стонать, им овладевала сонливость и отмечалось повышение сопротивления кожи ладоней, степень которого «была приблизительно пропорциональна продолжительности приступа». Для этого случая характерны парасимпатические разряды и даже парасимпатическая настройка вегетативной системы, так что болевые раздражения, исходящие из брюшной полости, вызывали вместо активации симпатической системы признаки повышения тонуса парасимпатической системы: наступала сонливость и повышалось сопротивление кожи ладоней. Предполагается, что эти симптомы связаны с состоянием периодически усиливающейся парасимпатической активности при соответствующим образом чередующихся восходящих и нисходящих разрядах.
ГЛАВА XXIV
Тот факт, что при этих и других периодических заболеваниях, не сопровождающихся психозами, постоянно наблюдаются эмоциональные расстройства [755], до известной степени подтверждает предположение о том, что одним из существенных .причинных факторов является нарушение гипоталамического равновесия. Даже если правильность этого предположения была бы доказана, тем йе менее причина столь значительной продолжительности циклов при этих болезнях, длящихся часто десятилетиями, остается неясной. В овете физиологического исследования Рихтера и клинического анализа периодической кататонии, к обсуждению которого мы сейчас приступим, представляется вполне вероятным, что в основе этих явлений лежит изменение взаимоотношений между гипоталамусом и эндокринными железами.
ПЕРИОДИЧЕСКАЯ КАТАТОНИЯ
Со времени классического исследования Гьессинга [374, 375] известно, что для некоторых случаев шизофрении характерны регулярные циклы, в течение которых периоды психического расстройства чередуются с относительно нормальными фазами. Эти ритмические изменения психического состояния сопровождаются отчетливыми изменениями обмена веществ. Гьессинг прежде всего обращает внимание на ритмические изменения скорости выведения азота у этих больных. Он выделяет тип А, у которого акинетический ступор наступает одновременно с нарушением азотистого баланса, и типы В и С, у которых кататоннческое возбуждение возникает через возрастающие интервалы после достижения максимального азотистого баланса. Он считает, что критической точке кривой азотистого баланса соответствует высвобождение токсина, действующего на гипоталамус и вызывающего психические расстройства. Хотя данных, подтверждающих правильность этого мнения, не существует, дальнейшие исследования обнаружили хорошую корреляцию между интенсивностью выведения воды и хлоридов и циклическими психическими явлениями. Было обнаружено, что возбуждение и кататонический ступор связаны с уменьшением выведения воды и соли.
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ
Значительные сдвиги вегетативной активности совпадают с психическими сдвигами [390, 425,. 749]. Во время ступора, так же как и при возбуждении, частота сердечных сокращений, кровяное давление, температура тела и основной обмен сильно возрастают, зрачки расширяются, выделяется густая слюна и сокращается длительность сна. Частота пульса 44—50 в 1 мин и кровяное давление около 100 мм рт. ст., которые характерны для неактивного периода, возрастают соответственно до 90— 130 в 1 мин и 140 мм рт. ст. в течение реактивного периода. Уровень сахара крови натощак и сахарная кривая в период психического расстройства выше, чем в период между приступами. Величина и направление этих изменений не- оставляют сомнения в том, что психическое расстройство связано с усилением симпатических и ослаблением парасимпатических разрядов.
Понижение обмена, характерное для неактивной фазы, позволяет (Предположить гипофункцию щитовидной железы; данные, приведенные на фит. 51, показывают, что лечение тироксином часто приводит к поразительным результатам. Периоды кататоническото ступора или возбуждения прекращаются, а интенсивность обмена возрастает. В других случаях эффективным оказалось лечение кортизоном, предпринятое в связи с некоторыми нарушениями уровня экскреции стероидов надпочечниками [390]. Следует отметить, что периоды кататоническото ступора обычно сопровождаются усилением симпатических разрядов и повышением интенсивности обмена [фиг. 51].
Хотя симптомы возбуждения симпатической системы проявляются как в фазе ступора, так и в фазе возбуждения, тем не менее между этими двумя состояниями существуют характерные различия, о чем свидетельствуют результаты применения психотропных веществ. Состояние апатии и подавленного настроения сменяется приподнятым настроением и раздражительностью, переходящими в максимальную активность эрготропной системы во время фазы возбуждения. Ступору, напротив, предшествует эйфорически-оптимистическое настроение и птеркинезия. Примерно через 10 час настроение становится подавленным, снижается общая активность и,

Фиг. 51. Состояние симпатической системы при кататонии [39].
А. Влияние тироксина на психическое состояние больного, страдающего регулярными двухмесячными периодами интенсивного возбуждения с четырехмесячными интервалам" Отмочены периоды возбуждения (закрашено черным) в течение года, предшествовавшего лечению тироксином, и исчезновение этих периодов под влиянием лечения. Данные Я январь — июль 1951 г. приведены в увеличенном виде. Тироксин вводили внутримышечно с 1 по 16 февраля (всего 122 .иг), а поддерживающую дозу давали до конца года. В. Связь между кататоническим ступором и возбуждением симпатической системы,
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ
наконец, наступает- ступор. Исследования показывают, что после .прекращения этой фазы во время ступора наблюдается тревожное состояние, чем объясняется наличие трофотропиых и эрготропных симптомов.
Гьессинг характеризует смену фаз при периодической кататонии следующим образом:
A
Неактивный период, предшествующий кататониче-скому возбуждению Актограмма свидетельствует о низкой общей активности, в особенности перед наступлением фазы возбуждения. Настроение колеблется между апатией и раздражительностью. Мышление замедленное, речь тихая — несколько слов, произносимых шепотом.
В
Неактивный период, предшествующий кататоничес-кому ступору Незадолго до наступления ступора наблюдается рост общей активности, связанный с эйфо-рически-оптимистическим настроением, но в этом настроении налицо отчетливый компонент раздражительности. Скорость мышления возрастает, но оно не критично.
' Б
Возбуждение Заметный гиперкинез; настроение повышенное, но раздражительное, ведущее при дальнейшем усилении психомоторного возбуждения к припадкам гнева и ярости и разрушительным действиям. Время сна укорочено, речь несвязна.
Г
Ступор Постепенное развитие психомоторного торможения. Снижение активности, подавленное настроение, возрастающее торможение мышления. Примерно через 10 час развиваются глубокий ступор и негативизм.
Строгий параллелизм .между развитием вегетативных явлений и психических сдвигов, а также тот факт, что при выздоровлении сначала исчезают психические и вегетативные симптомы [749] и лишь затем изменяется выведение азота и особенно интенсивность обмена, заставляют предполагать, что гипоталамус играет решающую роль в этой болезни. В подтверждение этого мнения Гьессинг ссылается на установленный им факт ускоре-
ГЛАВА XXIV
ния наступления активных фаз под влиянием психической травмы, а также ,на то, что вегетативные изменения, в частности ускорение пульса, могут на несколько часов предшествовать быстро развивающемуся ступору и за несколько дней указывать на приближение возбудительной фазы. Более того, характерные сдвиги со стороны водного и солевого обмена и поразительный терапевтический эффект при .применении гормонов щитовидной железы подтверждают гипотезу о нарушении функций гипоталамуса, затрагивающем выделение антидиуретического и тиреотропных гормонов. Очевидно, широкое использование новых представлений о гипоталамо-гипофизарных отношениях при анализе подобных случаев приведет к новым достижениям в этой области, а строить умозрительные предположения в настоящее время нецелесообразно. Тем не менее одно предварительное суждение кажется нам оправданным. Мы подчеркивали тот факт, что периоды психического расстройства, проявляющиеся в состояниях возбуждения и кататонического ступора, связаны с усилением симпатических разрядов и интенсивности обмена. Можно было бы ожидать, что в этих обстоятельствах гипоталамо-кортикальные разряды усилятся; однако, .наоборот, у некоторых [416], хотя и не у всех [75], больных обнаружено увеличение медленных волн в коре во время психического расстройства. Этот факт заставляет предположить, что при данной форме психического заболевания' существует диссоциация между восходящими и нисходящими разрядами гипоталамуса.
ГИПОТАЛАМУС, ЭНДОКРИННЫЙ БАЛАНС И ПСИХОЗЫ
Несмотря на недостаточность наших знаний и чрезвычайную сложность этой проблемы, нам кажется, что несколько наблюдений подтверждают предположение о том, что нарушения функции гипоталамуса играют важную роль в функциональных психозах.
* * * *
1 О циклическом поведении при других психотических состояниях СМ. [750, 752]. '
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ
При периодической кататонии и в .некоторых случаях шизофрении в период 'Спонтанной ремиссии наблюдается параллелизм между психическими нарушениями, вегетативными разрядами, изменением интенсивности обмена и выделением стероидов. Эти явления указывают на из менение регулируемой гипоталамусом секреции тирео-тропного и адреногршното гормонов. Они отражают изменение состояния гипоталамуса, а психические сдвиги, очевидно, связаны с изменениями гипоталамо-кортикаль-ных разрядов.
В норме уровень секреции «трапных» гормонов передней долей гипофиза поддерживается на довольно постоянном уровне благодаря существованию эффективного механизма обратной связи, который действует на центры гипоталамуса, а также и на саму переднюю долю гипофиза [428]. Чередование гипо- и гиперфункции щитовидной железы при .периодической кататонии и часто наблюдаемое понижение функции щитовидной железы у мужчин и повышение ее у женщин при шизофрении [746, 787], очевидно, указывают на то, что при шизофрении часто нарушаются указанные механизмы гомео-стаза. Возможно, с этим нарушением связан также более широкий диапазон колебаний выделения 17-кетостерои-дов при шизофрении по сравнению с нормой [787].
Не пытаясь дать подробный анализ влияния гормонов щитовидной железы на высшую нервную деятельность и на центральные вегетативные процессы, следует, однако, обратить внимание на следующие факты: тирео-идэктомия влияет на выработку условных рефлексов [25] и уменьшает реактивность вегетативных симпатических центров [336]; содержание иода в сыворотке больных с нейропсихическими расстройствами, не сопровождающимися заболеваниями щитовидной железы, значительно выше в случаях, характеризующихся наличием психического напряжения и состояния тревоги, причем изменения эти связаны с признаками вегетативной лабильности (колебания сердечного ритма, приливы, тошнота, понос и т. д. [105]); при хронической шизофрении уровень основного обмена обычно снижен [488]. Можно было бы думать, что снижение уровня обмена веществ, по крайней мере отчасти, является следствием сниже-
M
ГЛАВА XXIV
ния интенсивности регулируемой гипоталамусом секреции тиреотропното гормона, но и здесь проблема усложняется (вторичным?) снижением реактивности щитовидной железы [158]. Интересно, что болезнь Дауна [53] и некоторые случаи умственной отсталости связаны со снижением секреции тиреотропного гормона.
Изменения активности эндокринных желез («апри-мер, гипо- и гипертиреоз) обычно связаны с психическими расстройствами. Введение гормонов — кортизона и АКТГ — оказывает сильнейшее воздействие на личность, которое можно сравнить с изменениями, наблюдаемыми при функциональных лсихозах [146]. Несомненно, что эти гормоны оказывают влияние на все отделы нервной системы. Тем не менее, как нам кажется, судя по клиническим и физиологическим данным, роль гипоталамуса при этом является первостепенной. Гаррис и др. [429] показали, что им-плантация стилбэстрола в задний отдел гипоталамуса (но не в какие-либо иные участки мозга) вызывает типичное головое .поведение у овариэкгамиро-ваниых кошек. Кроме того, у кошек, которым введен фолликулин или во время естественной течки, раздражение влагалища вызывает типичные половые реакции и изменения электрической активности в гипоталамусе [721]. Весьма возможно, что соответствующие исследования с другими гормонами выявят новые важные связи между гипоталамусом и эндокринной системой.
НЕКОТОРЫЕ ЗАКЛЮЧИТЕЛЬНЫЕ СООБРАЖЕНИЯ
Цель данной части книги состояла в том, чтобы проанализировать на широкой основе те связи, которые существуют между патологическими состояниями психики и вегетативной нервной системой. С физиологической точки зрения интересно то, что эмоциональные нарушения, вегетативно-аффективный синдром, наблюдаемые и .у лиц, не страдающих .психозами, могут предшествовать возникновению психоза, а также возникать на некоторое время в период улучшения состояния больного, страдающего тем или иным психическим заболеванием. Отмечено также, что так называемые циклические заболевания у лиц с нормальной психикой и у .психически больных
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ
сопровождаются изменениями эмоционального состояния, совпадающими с ритмическими колебаниями в течении'этих болезней.
Психотические состояния в общем характеризуются сдвигами симпатической и парасимпатической реактивности, выявляемыми при пробах с мехолилом и норадре-налином, и нарушением вегетативного равновесия. Эндогенные депрессии характеризуются снижением симпатической реактивности; у нормальных людей подобное состояние можно вызвать с помощью фармакологических препаратов, вызывающих сдвиг вегетативного равновесия в сторону преобладания ■парасимпатической системы '. Подобный эффект можно нейтрализовать восстановлением вегетативного равновесия, либо путем раздражения симпатического отдела гипоталамуса посредством электрошока, либо с помощью фармакологических препаратов (аналептиков, ингибиторов, моноаминокси-дазы и т. д.]. Если такое раздражение продолжается слишком долго или если применяются избыточные дозьГ фармакологических препаратов, то развивается симпатическая гиперреактивность, которая связана с возбуждением и развитием маниакальных состояний.
При шизофрении .наблюдается как гипо- так и гиперреактивность симпатической системы (по данным пробы с мехолилом), что служит дополнительным показателем отсутствия единообразия в данной клинической труппе, а также указывает на необходимость анализа вегетативных нарушений в различных подгруппах при помощи целого ряда проб.
Значение вегетативных изменений ярко иллюстрирует тот факт, что глубокие изменения настроения и восприятия, вызываемые лизергиновой кислотой, отсутствуют при применении производных этого вещества, которые не оказывают влияния на вегетативную нервную систему.
Преобладание парасимпатического отдела связано с ослаблением общей активности, потерей мышечного то-* * * *
1 Преобладание парасимпатической системы, вызванное у нормальных людей длительным раздражением вестибулярного аппарата, также приводит к состоянию депрессии [398].
17-517
ГЛАВА XXIV
нуса, усталостью и состоянием каталепсии, тогда как преобладание симпатического отдела связано с повышенной активностью и усилением мышечного тонуса. Подобное сочетание изменений вегетативных и соматических функций характерно для различных состояний вегетативного равновесия на диэнцефальном уровне и отражается в активности соответственно трофотропной и эрготропной систем. Физиологические исследования, которые мы обсудили выше, показали, что сдвиг в равновесии систем гипоталамуса влияет также и на функции коры. Полагают, что качественные и количественные изменения гипоталамо-кортикальных разрядов (тесно связанных с активностью ретикулярной формации, но не идентичных с нею) служат источником психических сдвигов, наблюдаемых три эндогенных депрессиях и других функциональных психозах, или же способствуют их возникновению.
Изменения различных условнорефлекторных реакций в поведении подопытного животного после электрошока и изменения условных рефлексов и ЭЭГ, наблюдаемые при психических расстройствах, а также у нормальных людей под влиянием фармакологических препаратов, иллюстрируют, по крайней мере в основных чертах, этот процесс. Подобная физиологическая интерпретация и ее практическое значение впервые были изложены старшим из авторов в 1938 г. [294]. О правильности такого подхода свидетельствует все возрастающее число экспериментальных данных, накопленных за истекшие годы [296, 300, 306, 308, 313]. По-видимому, в настоящее время эта точка зрения находит признание. Например, ведущую роль вегетативной нервной системы в изменениях поведения под влиянием фармакологических препаратов подчеркивают Росл и и и Церлетти [141, 771]; Броди [1051 особо отмечает влияние фармакологических препаратов через сдвиг норадреналина на трофо- и эрготропную системы, а Рубин [780], подчеркивая различие зрачковых рефлексов у людей, страдающих психическими расстройствами, и в норме, делает из этого вывод о «нарушении функций адренэргического или холинэргического механизма или же их обоих». Очевидно, речь идет о тех же фактах, которые описаны нами при рассмотрении
ПЕРИОДИЧЕСКИЕ ПРОЦЕССЫ
парасимпатической и симпатической активности (гипоталамуса и вегетативного равновесия. Мориарти [664] и другие исследователи устанавливают связь между вегетативным 'состоянием, клиническим поведением и эффективностью некоторых видов терапии. Утверждают даже, что «условный рефлекс может быть выработан, изменен или угашен под влиянием одной лишь эмоциональной реакции {870]». Если это так, то, подходя с точки зрения физиолога, изучение гипоталамической системы должно было бы действительно послужить ключом к пониманию нормального и патологического поведения.
Вполне вероятно, что использование разнообразных вегетативных проб, оценивающих различные аспекты вегетативных функций гипоталамуса и ггапоталамонгипо-физарной активности, может в большой степени способствовать разрешению многих клинических проблем, например помочь пониманию механизма, посредством которого тип личности в период, предшествующий возникновению психоза, предопределяет его форму.
Наличие центральных вегетативных расстройств характерно как для психосоматических заболеваний, так и для функциональных психозов; однако между ними имеется серьезное различие. Астма может исчезнуть под влиянием эмоции. Голландские исследователи сообщали о случаях излечения язвенной болезни во время немецкой оккупации, несмотря на отсутствие специальной диеты. Описаны случаи исчезновения симптомов простудного .катара на время чтения больным лекции и возобновление их с новой силой по ее окончании {78]. Лихорадочное состояние может излечить от астмы и артрита. Во всех этих случаях гииерреактивность симпатического отдела гипоталамуса, обусловленная эмоциями или другими факторами, очевидно, подавляет парасимпатические разряды, которые вызвали психосоматические симптомы. Подобным же образом были обнаружены реци-■пронные отношения между астмой и психозом, о чем свидетельствовал характер реакции на мехолил при этих двух заболеваниях (270]. Переход из группы III по ме-холилу в группу I сопровождался прекращением приступов астмы, однако при этом развивалось состояние психоза. Очевидно, различные состояния центрального 17*
ГЛАВА XXIV
вегетативного равновесия связаны с психопатическим поведением и психосоматическими заболеваниями, и, изменяя состояние гипоталамуса (о чем можно судить по пробе € мехолилом ') посредством электрошока или имсулиноваго шока, можно вызвать либо астму, либо психоз.
В заключение мы хотели 'бы 'привлечь внимание к некоторым качественным изменениям вегетативных реакций у психически больных. Эти изменения, по-видимому, овязаны с состоянием вегетативной пасгройки, которая определяет, вызовет ли данный раздражитель симпатическую или парасимпатическую реакцию. К подобным парадоксальным реакциям относится, например, падение кровяного давления при пробах с действием холода '[499], повышение температуры тела в холодной ванне [122] при хронической шизофрении и повышение температуры кожи лица у психоневротиков под влиянием испуга (в норме в этих условиях отмечается понижение температуры) 1509]. Есть все основания полагать, что парадоксальные реакции при условных рефлексах у психически больных связаны с этими основными явлениями [15].
1 Конечно, проба с мехолилом отражает этот сдвиг весьма неполно, но дальнейшая дифференциация возможна с помощью проб с иорадреналином и других методов.
L
ЧАСТЬ СЕДЬМАЯ
ВЫВОДЫ И ПРЕДЛОЖЕНИЯ
ГЛАВА Х-Л V
ТЕОРИИ ЭМОЦИЙ
КЛАССИЧЕСКИЕ ТЕОРИИ
Прежде чем мы рассмотрим некоторые наиболее важные аспекты теорий эмоций, мы кратко опишем основные положения тех теорий, .которые вплоть до настоящего времени составляют основу наших представлений в этой области.
Теория Джеймса—Ланге
Согласно теории Джеймса — Ланге, эмоциональное ощущение представляет собой не что иное, как .восприятие изменений в функционировании внутренних органов и скелетной мускулатуры. Иными словами, эмоции являются скорее следствием, чем причиной висцеральных и мышечных реакций.
Шеррингтон [828] в ]90б г. выступил с возражениями против этой теории. Его возражения основывались на наблюдениях «эмоционального поведения» животных, лишенных значительной части сенсорной информации от различных областей тела. С тех пор были получены многочисленные дополнительные данные, основывающиеся главным обравом на работах Кэннона, которые с очевидностью доказывают несостоятельность этой теории. Хэбб возражает тем, кто стремится отвергнуть теорию Джеймса— Ланге на основании экспериментов, в которых центральная нервная система лишается притока сигналов от исполнительных органов. Хэбб [437] утверждает, что «это ^ возражение совершенно не относится к делу; Джеймс не утверждал, что эмоциональное поведение зависит от ощущений от конечностей и внутренних орга-
520 ГЛАВА XXV
нов». Согласиться с возражением Хэбба можно только в том случае, если принять, что проявление эмоций в поведении не является существенными показателями эмоциональных ощущений. Однако они являются единственными наблюдаемыми нами .признаками того, что животное .или человек («роме самого .наблюдателя) переживает эмоции, и с точки зрения здравого смысла мы должны признать их ценными показателями.
Таламическая теория Кэннона — Барда
Одна из важнейших теорий, выдвинутых в противовес концепции Джеймса — Ланге, это теория Кэннона — Бардам Согласно последней [133, 134], для эмоциональных ощущений существенными являются «таламические процессы». Как первоначально указывал Кэннон, простое переключение в таламусе импульсов, идущих к коре, приводит к восприятию воздействующего объекта, но не к эмоциональному ощущению. Если, однако, приход импульсов в кору приводит к снятию тормозных кортикальных влияний на таламус, высвобождаемые «таламические процессы» добавляют к восприятию образа «особое эмоциональное качество». Утверждалось также, что «таламические процессы» могут быть вызваны непосредственно афферентными импульсами, приходящими в таламус от рецепторов. В отличие от теории Джеймса — Ланге двигательное выражение эмоций рассматривается как результат эмоциональных «таламических процессов», а не как причина эмоциональных ощущений. Бард [37] утверждал, что «таламического процесса» самого по себе еще недостаточно. Он должен приводить к возникновению обратных сигналов, идущих в кору, и только при этом условии возникает эмоциональная окраска восприятия. Таким образом, в законченном виде теория Кэннона — Барда показывает, что эмоционально окрашенное восприятие зависит от прямых проекций от тала-муса к коре и от активации таламических процессов, которая вторично влияет на кору, приводя к возникновению эмоциональных ощущений. Учитывая тот факт, что Кэннон и Бард убедительно доказали, что эмоции исчезают после разрушения заднего отдела гипоталамуса, а также
ТЕОРИИ ЭМОЦИИ 521
то, что Гесе [454] показал различные типы эмоционального поведения при раздражении гипоталамуса у неиар-котизированной кошки, можно утверждать, что именно гипоталамус, а не таламус является центральной структурой в современном варианте теории Кэннона— Барда.
Активационная теория эмоций Линдслея
«Активационная теория» эмоций Линдслея [579] сохраняет основные положения теории Кэннона — Барда, но расширяет представление о механизмах возникновения эмоций, включая в состав участвующих в них структур и ретикулярную активирующую систему. Эта теория имеет отношение не только к эмоции, .но также и к кортикальной реакции пробуждения, соотношению сна и бодрствования и некоторым типам патологического поведения. Теория основывается на следующих экспериментальных фактах: 1) на электроэнцефалограмме во время эмоционального переживания -возникает реакция активации; 2) активация коры может быть вызвана раздражением ретикулярной формации ствола мозга; 3) при разрушении ростральной части ретикулярной формации кортикальная реакция активации исчезает, возникает апатия, сонливость .и т. д.; 4) ретикулярная активирующая система, возбуждающая кору, «или совпадает, или перекрывается» с механизмом, центрифугальные разряды которого приводят к двигательным выражениям эмоций. , ;
Теория Папеца. Эмоции и висцеральный мозг
В теоретической статье, опубликованной в 1937 г., Папец [692] выдвинул теорию эмоций, в которой приписывал первостепенную роль лимбической системе. Предполагалось, что кортикальные эмоциональные процессы возникают в гиппокампе. От гиппокампа импульсы идут через свод в маммилярные тела гипоталамуса, а затем в переднее ядро таламуса и оттуда — в поясную извилину. Папец 'рассматривал поясную извилину как рецептивную область эмоциональных переживаний. «Распространение эмоциональных процессов с поясной извили-
ГЛАВА XXV
«ы на другие области коры головного мозга создает эмоциональную окраюку психических процессов». В доказательство своей теории Палец ссылается на ряд исследований, показывающих, что целостность описанной цепи является необходимым условием для состояния бодрствования, и предположил, что она представляет собой механизм, организующий переживание и выражение эмоций.
На основании теоретической статьи Папеца и важных экспериментальных данных Клювера и Бьюси [546] Мак-Лин [604, 606] разработал теорию, согласно которой лимбическая система рассматривается как «висцеральный мозг». Термин «висцеральный мозг» не предполагает, что функции лимбической системы связаны исключительно с внутренними органами, но указывает, что эта область интерпретирует воспринимаемый сигнал в терминах эмоций, а не в виде '«интеллектуальных» символов. Интерпретируемые ею сигналы представляют собой информацию, получаемую от внутренней среды организма, под .которой Мак-Лин подразумевает все структуры тела, в том 'числе скелетную мускулатуру и внутренние органы. По мнению Мак-Лина, стратегическое положение висцерального мозга позволяет коррелировать все формы внутреннего и внешнего восприятия; оно обеспечивает формирование связей не только между оральными (обоняние, вкус, тактильная чувствительность слизистой полости рта) и висцеральными ощущениями, но также между ощущениями от половых органов, поверхности тела, -глава и уха. Он подчеркивает тот факт, что «в отличие от неопаллиума обонятельный мозг имеет многочисленные и прочные связи с гипоталамусом, в который поступают идущие от него импульсы». В то время как интеллектуальное выражение осуществляется с помощью словесных символов, для чего требуется высокое развитие новой коры, выражение эмоций может осуществляться через более примитивные нервные механизмы лимбической системы, где роль слов играет некий «язык органов». Он полагает, что «висцеральный мозг» имеет особое значение для эмоциональных процессов, а новая кора осуществляет интеллектуальные операции, требующие участия словесных символов.
ТЕОРИИ ЭМОЦИЙ
Эта теория представляет собой стройную схему, с которой без всяких оговорок согласуются экспериментальные данные, полученные за последние десятилетия, и которая намечает .пути для дальнейших исследований. Нам хотелось бы, однако, добавить .несколько замечаний, которые прежде всего касаются связи между ретикулярной формацией и висцеральным мозгом.
РАССУЖДЕНИЯ ПО ПОВОДУ ТЕОРИИ ЭМОЦИИ
Висцеральный мозг и ретикулярная формация
Согласно теории Мак-Лина, к висцеральному мозгу относится ряд структур — гиппокамп, миндалевидное ядро, поясная извилина, перегородка и 'гипоталамус. Несомненно, что раздражение или разрушение любой из этих структур изменяет функциональное состояние всей системы, а также влияет на состояние новой коры. Наряду с другими результатами это приводит к изменениям эмоциональной реактивности и проявлений эмоционального поведения. Необходимым условием осуществления нормальных и патологических функций висцерального мозга является активность ретикулярной формации. Она обеспечивает тонические импульсы не только для новой коры (в отсутствие этих импульсов возникает коматозное состояние [584]), но также для гипоталамуса и других частей висцерального моэга.
Хотя мы .подчеркиваем эту доминирующую роль ретикулярной формации, мы считаем, что теоретически следует проводить грань между гипоталамусом и ретикулярной формацией. Гипоталамус регулирует функции гипофиза, различные потребности, лежащие в основе инстинктивных форм поведения (голод, жажду, половую активность), интегрирует вегетативную и соматическую активность в характерные проявления эрготропной системы и отвечает за поддержание равновесия между парасимпатической (трофотропной) функцией переднего отдела и симпатической (эрготропной) активностью заднего отдела. Более того, гипоталамус оказывает сильное влияние на реактивность коры, о чем свидетельствуют опыты по разрушению заднего отдела гипоталамуса при
524 ГЛАВА XXV
сохранности ретикулярной формации [549]. Таковы функции, к которым ,не имеет отношения ретикулярная формация, обладающая более широкой, но менее дифференцированной активностью.
В подтверждение этого объяснения приведем цитату из 'рассуждения Хэррика [448] но поводу интеграции вегетативных функций: «Чрезвычайно важно учитывать, что все уровни интеграции переплетаются между собой и ни одни из них никогда не действует .независимо от других. Каждый высший уровень связан происхождением с низшим и может действовать только с помощью средств, предоставляемых в его распоряжение этими низшими уровнями. Тем не менее каждый уровень имеет свои специфические черты и своеобразные законы деятельности».
Поскольку ретикулярная формация и гипоталамус связаны восходящими и нисходящими путями, ясно, что в условиях эксперимента можно получить один и тот же эффект раздражением любой из этих структур [329], однако это не должно заслонять от нас основных функциональных различий между ними.
Реакция пробуждения и эмоции
Гипоталамус и ретикулярная формация тесно связаны между собой как анатомически, так и функционально. Поэтому нет ничего удивительного в том, что их раздражение вызывает пробуждение и эмоциональные реакции, а их разрушение вызывает сонливость и коматозное состояние независимо от того, затрагивают ли они задний отдел гипоталамуса или ретикулярную формацию. Обе структуры являются мультисинаптичесвдми, в результате чего проведение импульсов в них блокируется малыми дозами барбитуратов [264, 304]. Существуют, однако, некоторые данные, которые позволяют полагать, что господствующая тенденция в современной нейрофизиологии и психологии, пытающаяся приписать реакцию пробуждения и эмоции ретикулярной формации, зашла слишком далеко.
Так, исследование влияния фармакологических препаратов на реакцию пробуждения показало, что измене-
ТЕОРИИ ЭМОЦИЙ
ния реактивности ретикулярной формации не обязательно влияют на реакцию пробуждения. Физостигмин вызывает деоинхронизацию в коре (как при реакции пробуждения) , а атропин приводит к Синхронизации (как во время сна), однако на состояние бодрствования эти вещества не влияют [83, 947]. Не влияют они и на порог реакции пробуждения в поведении (открывание глаз, движение ушей, челюстей и т. д.), вызываемой раздражением ретикулярной формации «ли афферентных нервов. Но вместе с тем атропин повышает, а физостигмин понижает порог реакции пробуждения в ЭЭГ при раздражении ретикулярной формации. Таким образом, эти вещества создают условия для расхождения между проявлениями раздражения ретикулярной фор:.мации в поведении и на электроэнцефалограмме. Такое синхронизирующее вещество, как атропин, снижает эффективность влияния раздражения ретикулярной формации на ЭЭГ и ведет себя подобно другим синхронизирующим веществам (барбитураты), но не действует на поведенческие реакции, а физостигмин, который вызывает де-•синхронизацию кортикальных потенциалов (подобно фенамину), не вызывает пробуждения. Последние работы Брэдли и Кея [85, 86] ясно показали, что эффективность изменения потенциалов коры при электрическом раздражении ретикулярной формации может в значительной степени меняться под влиянием атропина и физостиг-мина, без изменений состояния бодрствования (фиг. 52). ^ Это означает, что при некоторых условиях ретикуло- fPpjr кортикальные отношения не определяют состояния бодр- ' т-„ ствования1.
Анатомические исследования показывают, что один f^ из путей идет от покрышки среднего мозга «через медиальный таламус в интраламинарной зоне, а другой идет вентро-латерально и продолжается в ростральном направлении к zona ineerta гипоталамуса» [800]. Исходя из этих данных, можно предложить другое объяснение: ре-
* * * *
1 Важная работа Деомедта [188], показавшего, что действие антихолинэстераз вызывает десиихроиизацию в коре даже после ее отделения от ствола мозга, не опровергает этот вывод.
526 глава xxv
акция пробуждения в первую очередь зависит от прямого и рефлекторного возбуждения гипоталамуса, а также от возбуждения, идущего через ретикулярную формацию и через подходящие к ней афферентные нервы [860, 861]. Таким образом, ^реакция пробуждения и эмоции возникают прежде всего в результате активации -висцерального мозга.
Фиг. 52. Влияние атропина и физостигмина на порог раздражения
ретикулярной формации, вызывающего реакцию пробуждения
в коре [85].
Опыты на кошках, препарат encephale isole. При увеличении дозы атропина (0,1—4,0 мг/кг) порог повышается С), но «реакция пробуждения о поведении» (//) остается неизменной. Стрелкой указано начало введения физостигмина; порог определяется после введения 0.01—0,8 мг/кг. Пунктиром показаны индивидуальные колебания.
Ретикуло-кортикальная система играет важную роль в .модификации состояния бодрствования. Следовательно, сдвиги от состояния покоя к состоянию внимания связаны с кортикофугальными влияниями на ретикулярную формацию, которые приводят к специфической фильтрации афферентных имлульсов [260]. Исчезновение реакции пробуждения при повторных предъявлениях одного и того же звука (привыкание) также зависит от .ретикулярной формации [824].

ТЕОРИИ ЭМОЦИЙ
Мож1но сослаться лишь на немногие физиологические и фармакологические исследования, подтверждающие наше мнение о том, что гипоталамус и висцеральный мозг играют ведущую роль в возникновении реакции пробуждения и эмоций. Рансон [734] рассматривал задний отдел гипоталамуса как центр бодрствования. По данным Наута [677], этот центр подавляется передним отделом гипоталамуса. Раздражение области миндалевидного ядра у кошек прежде всего вызывает реакцию пробуждения (внимание), и лишь затем возникают проявления страха или ярости [900]. Далее, реакция пробуждения, вызываемая присутствием наблюдателя [279, 573], изменяет электрические потенциалы гипоталамуса у подопытного животного. Наконец, некоторые фармакологические препараты, оказывающие глубокое влияние на состояние бодрствования, очевидно, изменяют функции гипоталамуса в большей степени, чем реактивность ретикулярной формации. Брэдли и Кей [86] указывают, что аминазин оказывает минимальное влияние, а ди-этиламид лизергииовой кислоты совсем не действует на реакцию пробуждения, вызываемую раздражением ретикулярной формации. Другие исследователи ясно показали, что реактивность гипоталамуса резко изменяется под влиянием этих веществ [81, 157, 166, 190] и что перерезка ствола мозга перед маммилярными телами устраняет характерные влияния указанных препаратов [459, 593, 684].
Эти данные показывают, что реакцию пробуждения у животного можно вызвать как с висцерального мозга, так и с ретикулярной формации. Существует постепенный переход от реакции пробуждения к эмоциональным реакциям, что согласуется с тесной психологической зависимостью, существующей между некоторыми видами реакций пробуждения и эмоциями. Действительно, трудно решить, является ли реакция кролика на присутствие наблюдателя активацией или эмоцией. В то время как грубые формы реакции пробуждения поддерживаются посредством тонической активности ретикулярной формации и гипоталамуса, а также его влияний .на висцеральный мозг, возникновение более тонких градаций внимания, необходимых для восприятия и сложных пои-
ГЛАВА XXV
хических процессов, очевидно, зависит от взаимодействия новой коры и ствола мозга.
Рефлексы, переживания и эмоции
Общепринято мнение [212, 448] о том, что в .процессе филогенетического развития примитивные эмоции связаны с рефлексами. На более поздней стации эти ощущения индивидуализируются, а <при дальнейшем развитии мозга, которое тесно связано с функциями дистантных рецепторов [828], возникает восприятие окружающей среды. С самого начала сенсорные процессы сопровождаются движениями. Действительно, одна из крупных теоретических школ, представленная Когхиллом [147] и Хэррвком [448], считает, что в «процессе эмбрио- и филогенеза внутренне .возбуждаемые двигательные функции предшествуют реакциям на внешние раздражите-? ли». Эти движения можно .разделить на две группы: движения, приводящие .к устранению от раздражающего объекта, и движения, приводящие к повороту в сторону этого объекта. Первые .направлены на снижение (опасного) контакта с объектом, последние способствуют ему.
В физиологических исследованиях уделяли больше внимания реакции устранения, типичным выражением которой является отдергивание конечности от .контакта с болевым раздражителем, чем второй группе рефлексов, которые обычно вызываются не электрическим раздражением сенсорных органов кожи, а контактом с большими .гладкими поверхностями. Так, давление на подошву вызывает реакцию разгибателей (основа реакции поддержания тела), а такое же раздражение ладони вызывает хватательный рефлекс, осуществляющий соответствующую роль у птиц, обезьян и т. д. Наиболее биологически важным рефлексом подобного типа является оральный рефлекс, который лежит в основе приближения рта к соску и обеспечивает тесный .контакт, необходимый для сосания. Другие известные примеры движений, принадлежащих к этой группе, вызываются поглаживанием и другими подобными воздействиями.
ТЕОРИИ ЭМОЦИИ
Характерно, что рефлексы и действия, .приводящие к движению в направлении объекта, служащего источником раздражения, сопровождаются ощущением благополучия в отличие от тех, которые заключаются в устранении от объекта. Боль и удовольствие, очевидно, возникли на этой основе. Хотя с развитием коры головного мозга эти ощущения стали более сложными, их можно обнаружить в поведении '.младенцев, родившихся без больших полушарий. Так, при поглаживании прекращается плач, а громкие звуки, как было обнаружено, вызывают реакцию съеживания даже при отсутствии полушарий мозга [214].
Прототип неприятных телесных ощущений возникает в связи с рефлекторным устранением от острых, шероховатых или угловатых предметов. При этом двигательные реакции связываются с вегетативными. Так возникает эрготропная система. Ее активация обеспечивает характерное выражение неприятных ощущений, а на более высоких стадиях развития — выражение эмоций, сопровождающих агрессивное поведение и убегание. Напротив, в ответ на раздражения, наносимые округлыми, мягкими и гладкими предметами, возникают Zuwen-dungsreflexe (положительные рефлексы), связанные с приятными ощущениями и образующие основу положительных эмоций [212].. Снижение мышечного тонуса и тенденция к засыпанию при поглаживании позволяют считать, что ощущение благополучия связано с торможением эрготропной системы и повышением тонуса парасимпатического отдела вегетативной нервной системы.
Сложная эмоциональная надстройка возникает на основе диффузных телесных ощущений, связанных первоначально с рефлексами устранения и приближения, обнаруживаемыми у низших видов животных и у детей-анэнцефалов. Тем не менее эти ощущения сохраняют глубокое влияние на эмоциональное и интеллектуальное поведение человека. Мы называем их уже не телесными ощущениями, а настроениями. Вряд ли можно сомневаться, что гипоталамус участвует в их осуществлении [528]. -Можно предположить, что его тоническое состояние является основой настроений, а его фазические реакции связаны с эмоциями.
ГЛАВА XXV
Связь, очевидно существующая между телесными ощущениями и некоторыми рефлексами, с одной стороны, .и иипоталамичеокими функциями и эмоциями, с другой, находит свое отражение в соответствующих формах организации физиологических процессов. Считают, что эмоции связаны с восходящими и нисходящими разрядами в гипоталамусе, первые из которых в своих многообразных формах служат физиологической основой возникновения различных эмоций [301]. Аналогичным образом отмечалось [212], что, хотя большинство висцеральных функций, например функции кровеносной и дыхательной систем, не достигает уровня сознания, прием пищи и выделительные функции связаны соответственно с ощущениями голода и жажды, давления в мочевом пузыре и прямой кишке, доминирующими в сознании и направляющими действие в соответствующих условиях. Существует параллелизм между интенсивностью реакции опорожнения мочевого пузыря и прямой кишки и интенсивностью соответствующих ощущений', которые можно сравнить с отношениями между ноцицептивными рефлексами разной интенсивности и соответствующими степенями эмоционального возбуждения. Сходным образом при аноксии, возникающей в эксперименте или клинике, усиление циркуляторных и дыхательных рефлекторных реакций происходит параллельно с нарастанием ощущения недостатка воздуха. Этот параллелизм иллюстрируется также тем, что когда повышенная при гипоксии возбудимость дыхательного аппарата снижается при глотательном движении, ощущение недостатка воздуха также становится менее выраженным.
Отсюда следует, что в связи с висцеральными рефлексами и развитием механизмов гомеостава (в отношении содержания кислорода, воды, сахара и т. д.) возникли некоторые субъективные явления, (которые в зависимости от степени их сложности обозначаются как телесные ощущения или эмоции. Их первостепенная за-
* * * *
' Эти телесные ощущения отличаются от внешних ощущений. И те и другие возникают при (раздражении рецепторных органов, но последние дают информацию о внешней среде только через дистантные рецепторы.
ТЕОРИИ ЭМОЦИИ 531
дача состоит в усилении и ускорении двигательных реакций, направленных на .восстановление гомеостаза. Тем, кто возражает 'против такой телеологической формулировки, полезно вспомнить следующее высказывание Шер-рингтона: «Физиология занимается анализом реакций тела, рассматриваемых как физические «ли химические явления; однако в дальнейшем она стремится дать обоснованные объяснения дейс.твий организма в связи с их целью и пользой для организма как целого. Это можно назвать телеологической тенденцией, но такая телеология не выходит за рамки науки». То же относится и к эмоциям, которые могут 'Служить поддержкой при нападении у животных. Несомненно, что у человека в связи с возникновением гипоталамо-кортикальных связей эмоции имеют более сложную функцию и приобретают важнейшее значение в его психической деятельности.
Фармакологические препараты, тонус телесных ощущений и настроение
Обширные экспериментальные исследования, проведенные на человеке и животных с применением психофармакологических препаратов, дали ценный материал для изучения физиологических механизмов телесных ощущений и настроений. Хлорпромазин и морфин (а также их производные) выбраны как представители депрес-сирующих препаратов, а амфетамин (фенамин) и другие вещества того же ряда — как представители аналепти-ческих веществ. Хотя между морфинам и транквилизаторами [271] существуют некоторые различия, в общем для всех этих веществ характерно следующее: снижение возбудимости ретикуло-гапоталамичеокой системы, сдвиг функционального равновесия гипоталамуса в сторону преобладания парасимпатического отдела, снижение интенсивности диффузных разрядов, идущих от ретикулярной формации и гипоталамуса к коре, и снижение порога возникновения веретен и реакций вовлечения, вызываемых раздражением таламуса или хвостатого ядра [280]. Вследствие этого в ЭЭГ появляются большие медленные волны, характерные для она, и начинает доминировать трофотропная система. Наоборот, фенамин
ГЛАВА XXV
и подобные ему вещества действуют противоположным образом, приводя к реакции пробуждения (десинхрони-зации) в ЭЗГ, разрядам в симпатическом отделе и повышению общей активности, что указывает на преобладание эртотропной системы в функциях гипоталамуса. Исследования, проведенные на животных, где настроение определялось на основании поведения, показали, что у кошек и обезьян хлордромезин и метамфетамин являются антагонистами: три действии хлорлромазина ощущение удовольствия (которое у кошек определяется по реакциям умывания и мурлыканья, а у обезьян — по игровым, пищевым и .гигиеническим реакциям) усиливается, а оборонительные ,и агрессивные .реакции снижаются; метамфетамин действует противоположным образом [688]. Принципиальные различия в настроении и поведении под влиянием указанных типов фармакологических препаратов отчетливо выступают также в исследованиях на людях, хотя полного совпадения этих результатов с данными экспериментов на животных не наблюдается. Хлорпромазии вызывает дисфорию (ощущение грусти) и успокоение (сонливость), а реакция на морфин и героин у нормальных людей выражается в описанной многими лицами грусти, меланхолии, расслабленности, сонливости, успокоении, головокружении, ощущении неудобства, дрожи, усталости, плохом настроении. Фенамин и другие родственные ему вещества, напротив, вызывают эйфорию (ощущение счастья) и бодрствование [432]. Испытуемые переживают ощущение радостного возбуждения и оптимизма, а со стороны они кажутся напряженными, возбужденными и болтливыми [564], что указывает на доминирование эрготропной системы. Наоборот, часто наблюдаемые после введения морфина и хло.рпромавина сонливость, тошнота, рвота и головокружение указывают на доминирование трофо-тропиой системы. Необходимо добавить, что при малых дозах морфина наблюдались сонливость и ощущение покоя и удовлетворения. Очевидно, это показывает, что незначительные сдвиги в равновесии систем гипоталамуса в сторону преобладания 'парасимпатической системы проявляются в ощущении покоя и удовлетворения, близких к состоянию перед засыпанием, тогда как более
ТЕОРИИ ЭМОЦИИ
выраженные изменения приводят к состоянию депрессии. Сдвиг равновесия в сторону преобладания симпатической системы приводит к бодрствованию и ощущению счастья, тогда как у животных проявляется только усиление агрессивных реакций. Повышение психических функций воспринимается человеком как ощущение успеха. Усиление типоталамо-кортикальных разрядов у животных может быть связано с усилением агрессивности, а у человека с повышением -психомоторной активности и маниакальным поведением.
Кофеин и фенамин ускоряют и усиливают условные реакции [246] и замедляют угашение условных рефлексов, в то время как депресоирующие агенты, такие, как алкоголь и амитал натрия, оказывают противоположное действие.
Эти наблюдения поддерживают гипотезу о том, что депрессия связана с преобладанием трофотропной системы, а подъем—с преобладанием эрготротшой. Ай-зенк [230] считает, что эти состояния связаны с определенными особенностями личности (интроверзия и экстро-верзия), а также с невротическими явлениями.
Нам кажется вероятным, что 'индивидуальные различия реакций на зти препараты зависят от характера равновесия систем .гипоталамуса [919], однако, чтобы доказать правильность этого предположения, необходимы дальнейшие эксперименты.
Значение этих выводов для нейропсихиатрии очевидно. Можно предполагать, что некоторые депрессивные состояния связаны с повышением реактивности центральных парасимпатических структур и снижением активности симпатических механизмов. Вторую часть этого предположения подтверждает проба с мехолилом, однако экспериментальных данных, подтверждающих первую его часть, пока не существует. При состояниях, связанных с агрессивностью и ненавистью, предполагаемое повышение реактивности симпатической системы действительно подтверждается пробой с мехолилом и секрецией норадреналина. С нашей концепцией согласуются также данные о повышении секреции норадренаг •пина при маниакальном состоянии и о ее понижении при Депрессии.
S34
ГЛАВА XXV
Основы качественных различий эмоций
Теорию, которая не раскрывает механизмы, определяющие количественные и качественные различия эмоций, нельзя считать завершенной как в теоретическом, так и в практическом плане. Поэтому мы рассмотрим прежде всего физиологические основы качественных различий эмоций.
Мы строим наш анализ не на центральных процессах, лежащих в основе эмоций, а на сопровождающих их периферических разрядах в вегетативной нервной системе. Еще в 1924 г. Олпорт [17] связывал приятные и неприятные эмоции соответственно с парасимпатической и симпатической системами. Действительно, точно установлено, что вид и запах пищи возбуждают парасимпатические влияния на желудочно-кишечную систему и ее железы и что в состоянии пищевого удовлетворения активность эрготропной системы снижается, а парасимпатической— повышается. Однако не нужно идти далеко, чтобы обнаружить вопиющие противоречия. Печаль сопровождается плачем, возникающим в результате парасимпатической импульсации, а возмущение связано с усилением перистальтики, расширением сосудов слизистых оболочек и прочими симптомами. Не только приятный запак съедобной пищи, но и неприятные запахи стимулируют секрецию, регулируемую парасимпатическими нервами, и могут вызвать рвоту. Вместе с тем усиленная перистальтика мочевого пузыря и прямой кишки, характерные симптомы возбуждения парасимпатической системы сопровождают такие ощущения, как страх и беспокойство.
Со времени работ Кэннона общепризнано, чго испуг стимулирует симпатические или симпато-адреналовые разряды или, по терминологии Гесса, вызывает активацию эрготропной системы. Однако примеры, приведенные здесь и в гл. IV, показывают, что это утверждение в некоторых случаях является чересчур узким, а в других— совсем неверным. Так, тяжелые эмоциональные расстройства связаны и с симпатическими, и с парасимпатическими разрядами, первые из которых обнаруживаются по реакциям сосудов, а вторые — по изменениям
ТЕОРИИ ЭМОЦИИ 535
со стороны внутренних органов (иногда половых). В качестве одного из примеров патологической настройки гипоталамуса [310] укажем, что при неврозах эмоциональный стресс вызывает падение уровня сахара крови (через ваго-инеулиновую систему), а не его повышение [12, 722 а].
Если ограничить наше обсуждение нормальными реакциями, то следует признать, что эмоции умеренной интенсивности, если они приятны, связаны преимущественно с парасимпатическими разрядами, а если они неприятны и болезненны, то с симпатическими. При повышении степени эмоционального возбуждения нисходящие гипоталамические разряды не ограничиваются одним отделом вегетативной нервной системы. Наиболее выразительным примером приятных эмоций высокой интенсивности, вызывающих такой тип разрядов, является оргазм, при .котором сосудистые и соматические изменен.ия указывают на активацию эрготропной системы, а реакция половых органов обнаруживает одновременные парасимпатические разряды. Такое двойное действие отражает двойственную природу процесса: он занимает пограничное место между удовольствием и болью'. Однако в нормальном организме тяжелые (неприятные) эмоции также включают симпато-адреналовую и вато-инсу-линовую системы при преобладании первой. В состояниях предельного испуга и ужаса циркуляторный коллапс и замедление сердечного ритма являются результатом возросшей парасимпатической активности, а расширение зрачка, потоотделение свидетельствуют о повышении симпатических разрядов. В этом случае наблюдается также диссоциация между симпатической активностью и тонусом скелетной мускулатуры2.
Наиболее значительным выводом из этих рассуждений является представление о сложной зависимости эмо-
* * * *
1 Хордотомия (перерезка спино-таламического тракта в передне-латеральном пучке .спинного мозга) вызывает исчезновение оргазма, хотя рефлекторно вызываемая эрекция и эйякуляция сохраняются.
' Напомним также о возникновении тошноты и рвоты при условном сигнале болевого раздражителя у кошки, близкой к невротическому состоянию (си. гл. XVI).
536 ГЛАВА XXV
ций от вегетативной нервной системы, особенно при высокой интенсивности. Хотя в организме, находящемся в состоянии покоя, сдвиг >в равновесии вегетативных систем на уровне гипоталамуса определяется принципам реципрокной иннервация [356], при сильном эмоциональном возбуждении (а также при других условиях, вызывающих интенсивное возбуждение вегетативных центров, например при судорогах, вызванных электрошоком [239, 529]) происходит отклонение от этого правила.
Поскольку изменения вегетативного равновесия в гипоталамусе, вызываемые в эксперименте у животного раздражением, введением фармакологического препара-та или разрушением, приводят к изменениям восходящих и нисходящих разрядов гипоталамуса [59, 310, 549] и поскольку первые связаны с соответствующими «психическими» сдвигами, возникает вопрос о том, каковы характерные отношения, существующие между нисходящими вегетативными разрядами, с одной стороны, и качеством и интенсивностью эмоций — с другой.
Для подтверждения наличия специфических отношений между «неприятными» эмоциями и симпатическими разрядами напомним читателю эксперименты по после дованию ярости: 1) при естественных условиях; 2) в результате раздражения симпатического отдела гипоталамуса различными методами; 3) после высвобождения активности этого отдела путем устранения тормозных влияний со стороны вентро-медиального ядра гипоталамуса [943]. Сходные результаты были получены при разрушениях перегородки [94]. Напротив, отношения между парасимпатической системой и приятными эмоциями, очевидно, выступают в опытах Гесса с раздражением перегородки и переднего отдела гипоталамуса [641]; в этих опытах облизывание и другие поведенческие реакции позволяли предположить возникновение у животных ощущения удовольствия. Подобный эффект удалось получить также с различных частей висцерального мозга, например с гиппокампа (особенно при химическом раздражении [603]). Более того, раздражение гиппокампа сначала сопровождалось периодом разрядов последействия, связанных с признаками возбуждения симпатического отдела (расширение зрачка, пилоэрекция), а
ТЕОРИИ ЭМОЦИИ
затем фазой «благополучия», характеризующейся вылизыванием и другими проявлениями материнского инстинкта по отношению к котенку, к которому до этого животное проявляло агрессивную реакцию [698]. В этом случае вновь сдвиг вегетативного равновесия сопутствует фундаментальным изменениям эмоционального поведения. Тесная связь удовольствия и половых реакций со сниженной реактивностью к болевым раздражителям часто отмечалась в экспериментах с раздражением лим-бической системы [18].
В то время как раздражение миндалевидного ядра вызывает изменения поведения, указывающие на появление страха и ярости [959], удаление миндалевидного комплекса у кошек приводит к гиперсексуальности .и к «усилению .реакций удовольствия на поглаживание и похлопывание» [541]. При этом эмоциональные реакции, связанные с усилением разрядов в симпатическом отделе гипоталамуса, такие, как страж и агрессивное поведение, а также реактивность к болевым раздражителям, оказываются сниженными. Однако эти характерные сдвиги не 'возникают, если перед амигдалэктомией произвести двухстороннее разрушение вентро-медиальных ядер гипоталамуса, или устраняются, если эти ядра' разрушить после амигдалэктомии. Такие животные обнаруживают поведение, характеризующееся одичанием, чго обычно наблюдается у кошек с указанными разрушениями гипоталамуса. Отсюда следует, что амигдалэктомия приводит к повышению активности вентро-медиальных ядер, которые, как известно, тормозят активность симпатического отдела гииоталагмуса. Эти эксперименты показывают, что разрушения висцерального мозга изменяют эмоциональное поведение через гипоталамус1.
1 Эксперименты Всйекранца и Вильсона [936] в соответствии с описанной выше работой показали, что после амигдалэктомии у обезьян наблюдается повышение порога реакции избегания, вызываемой болевым раздражителем. Оперированные животные по-прежнему реагируют па резерпин, подобно нормальным. Оба эти факта можно объяснить на основании данных, приведенных здесь и в гл. III и показывающих, что лимбическая система изменяет возбудимость гипоталамуса и вызывает сдвиг в равновесии его систем и что резерпин действует на промежуточный мозг.
ГЛАВА XXV
Однако «е следует забывать, что экспериментальные даиные по раздражению и разрушению лимбичеокой системы довольно противоречивы. В какой мере несоответствие зависит от видовых различий [40, 545] и локализации разрушения [959], .не вполне ясно. Лабильность этих функций .позволила Глуру [382] предположить, что «миндалевидное ядро ,и другие части обонятельного мозга модулируют активность, интегрированную в подкорковых структурах». Поскольку активность лимбичеокой системы тесно связана с гипоталамусом, если не зависит от него полностью, следует помнить, что уровень реактивности гипоталамуса весьма изменчив и что равновесие его систем варьирует в широких пределах [310].
В пользу предположения о том, что раздражение или повреждение некоторых частей лимбической системы изменяет эмоциональное поведение через сдвиг равновесия систем .гипоталамуса, свидетельствуют эксперименты на крысах, у которых последовательно разрушали перегородку и миндалевидное ядро. Мы уже видели, что раздражение перегородки вызывает положительные реакции н тормозит эртотропную систему, а раздражение миндалевидного ядра вызывает страх и ярость [289]. Наоборот, разрушение миндалевидного ядра снижает эмоциональные реакции (это особенно отчетливо видно на неприрученных, диких крысах, которые становятся ручными после операции [961]), а разрушения перегородки приводят к резкому их повышению [537]. Этот эффект полностью устраняется последующим удалением миндалевидного ядра; в контрольной группе, у животных с разрушениями в области новой коры и поясной извилины, обнаруживается лишь незначительное снижение эмоциональной реактивности. Далее, повышение эмоциональных реакций после разрушений перегородки было значительно менее выражено у крыс с ам.итдалэкто-мией, чем у животных контрольной оперированной группы (фиг. 53).
Если правильно наше утверждение о связи сдвигов в равновесии вегетативных центров гипоталамуса со сдвигами эмоционального состояния, то можно ожидать, что патологические процессы у человека, приводящие к усилению симпатических или парасимпатических разря-
ТЕОРИИ ЭМОЦИИ
дов, должны сопровождаться различными эмоциональными состояниями. Поэтому имеющиеся у «ас сведения об эмоциях и их связи с гипоталамусом, которые до сих пор черпались главным образом из наблюдений над

Фиг. 53. Интенсивность эмоциональных реакций у крыс после разрушения перегородки, миндалевидного ядра и коры [537].
А. Повышение интенсивности эмоциональных реакций после разрушения перегородки, исчезающее после разрушения миндалевидного ядра; 15 крыс были разделены на 2 группы; установив контрольный уровень эмоциональных реакций (/). в обеих группах произвели разрушение перегородки, в результате чего резко усилились эмоциональные реакции (//). Затем у 10 крыс (сплошная линия) произвели разрушение миндалевидного ядра (///) — уровень их эмоциональных реакций восстановился. У других 5 крыс (пунктирная линия) были произведены разрушения поясной извилины и новой коры для контроля влияния самого оперативного вмешательства; у них обнаружилось лишь незначительное и постепенное снижение эмоциональных реакций.
Б. 1 — контроль; // — отсутствие изменений эмоциональных реакций после разрушений коры (новая кора или поясная извилина) или миндалевидного ядра; /// — крысы с разрушениями перегородки после повреждения коры обнаруживают повышение эмоциональных реакций, сравнимое с эффектом разрушения одной только перегородки, в то время как аналогичные разрушения перегородки после амигдалэктомии вызывают значительно менее выраженное усиление эмоциональных реакций. На оси абсцисс обозначены дни после операции.
эмоциональным поведением животных, должны пополниться за счет словесных отчетов больных. Особый интерес представляют наблюдения над больными с цирку-ляторными нарушениями диэнцефапьного происхождения. Как показано в гл. XV, у таких больных бывают приступы крайнего возбуждения системы блуждающего нерва (замедление ритма или полная остановка сердца и падение кровяного давления) или симпатической системы (максимальное ускорение ритма и повышение
ГЛАВА XXV
кровяного давления). Поведение этих двух групп больных существенно различается [112]. Больные с сосудистыми приступами парасимпатического происхождения обычно тихие, довольно спокойные, они жалуются на слабость, 'головокружение и приступы in отер и сознания, но не производят 'впечатление людей, взволнованных своим состоянием. Больные, страдающие от приступов симпатического происхождения, наоборот, выражают крайнее беспокойство, сильно напуганы (Vernichtungs-gefiihl), хотя объективные данные указывают на относительную безопасность их состояния. Для первой группы характерно состояние, близкое к расслаблению и сну, для второй — состояние максимального эмоционального возбуждения, хотя у больных первой труппы значительно больше оснований для беспокойства. Связь между парасимпатическими разрядами .в системе гипоталамуса и покоем, с одной стороны, и между симпатическими разрядами и эмоциональным возбуждением, с другой, подтверждается в экспериментах, где подобные сдвиги активности вегетативной системы вызывали у человека рефлекторно путем введения норадреналина и мехолила соответственно [311]. Введение норадреналина приводило к рефлекторному повышению парасимпатической активности и успокоению, а у некоторых больных вызывало сон, тогда как вызванные мехолилом симпа-гичеокие разряды сопровождались ощущением напряжения in раздражительностью. Эти наблюдения подтверждают нашу гипотезу о том, что центральные (гипоталамические) разряды, зависящие, очевидно, от разных частей гипоталамуса, приводят к принципиально различным восходящим и нисходящим влияниям. Предполагается, что характер восходящего разряда, в значительной степени определяющий характер эмоции, зависит не только от того, какая область гипоталамуса возбуждена, но и от обратной связи, идущей от иннервируе-мых вегетативной нервной системой органов. Только в этом смысле можно говорить о том, что периферические вегетативные реакции влияют на состояние гипоталамуса и таким образом модулируют эмоции.
Кроме того, эти рассуждения показывают, что различные эмоции нельзя объяснять только на основании
ТЕОРИИ ЭМОЦИИ 541
фактора интенсивности, как это делает активационная теория. В .настоящее время .кажется, что качественные различия .между эмоциями, которые на основе психологического анализа требуют трехмерного изображения [802], физиологически зависят от парасимпатических, симпатических .и смешанных разрядов, связанных с соответствующими дифференцированными формами активации висцерального мозга и .новой коры. Далее, следует помнить, что эмоциональное возбуждение по крайней .мере при наиболее высоких уровнях ведет к высвобождению яорадреналина, адреналина и инсулина, а также к регулируемой гипотала1мусом секреторной активности гипофиза. Весьма вероятно, что на изменение характера гипоталамо-корт.икальных разрядов влияют не только адреналин и норадреналин, действие которых на ретикуло-шпоталамическую систему было подробно изучено [768], но и другие гормоны, высвобождаемые в результате активности •гипоталамо-гипофизарной системы [960]. С повышением степени возбуждения гипоталамуса возникают изменения качества эмоций, например наблюдается переход от страха ,к ярости [289]. Предполагается, что это изменение представляет собой не только результат .возрастающей частоты разрядов и увеличения числа разряжающихся нейронов, но также следствие изменения конфигурации разрядов, связанных со сдвигами тонуса и активности поперечнополосатых мышц. Обратная связь от соматической нервной системы при страхе и ужасе иная, чем при ярости; роль проприоцептивной импульсации для поддержания возбудимости типотала.муса и интенсивности типотала.мо-кор-тикальных разрядов была показана в гл. XVII. Этим объясняется тот факт, что легкая кураризация может привести .к превращению эмоциональной реакции ярости в реакцию страха вследствие снижения проприоцептивной импульсации в расслабленных мышцах [311].
О значении интенсивности эмоций
До сих пор мы подчеркивали качественные изменения периферических вегетативных разрядов, сопровождающих различные виды эмоционального возбуждения,
ГЛАВА XXV
однако количественный аспект не менее важен. Линдслей [579], а также другие авторы отметили наличие непрерывного перехода от она' к эмоциональному возбуждению, которое выражается сдвигами в электроэнцефалограмме и указывает на возрастание степени активности ретикуло-гипоталамической системы. В то время как но мере перехода от «настороженного внимания» к сильным эмоциям, таким, как страх или ярость, степень активации как бы постепенно нарастает, это постепенное нарастание не распространяется на .все поведенческие, вегетативные и электроэнцефалографические изменения. Так, при эмоциональном возбуждении у человека внимание на определенном объекте (раздражителе) концентрируется с трудом, реактивность к раздражителям симпатической системы снижается (возможно, на гипо-таламическом уровне), а спонтанная активность возрастает (фиг. 54). Эти явления, изучавшиеся у человека в опытах с кожно-тальваническим рефлексом, очевидно,

Фиг. 54. Теоретические отношения между степенью активации (/) и реакцией на электрическое раздражение (//) [151].
КГР - кожно-гальваничсский рефлекс; каждая стрелка указывает момент нанесения специфического стандартного раздражения а отклонения соответствуют сдвигу кожного сопротивления. Иа трех первых стадиях активации кожно-гальванический рефлекс возникает в ответ на ««ирую, ;ее Раздражу ние. причем его амплитуда прямо зависит от степени активации Однако в состояниях возбуждения и паники становятся все более частыми «понтаи ны» сдвигиГКГ'Г> (неизвестные раздражители), а интенсивность реакции на стандартный раздражитель снижается.
ТЕОРИИ ЭМОЦИЙ
связаны с усилением спонтанных симпатических разрядов, которые можно обнаружить ло сокращению третьего века у подопытного животного после внутривенного ГЗЗЗ] или интратипоталамичеокого введения метразола [325]. Очевидно, состояние резко повышенного симпатического возбуждения гипоталамуса характеризуется повторными «спонтанными» симпатическими разрядами.
Предполагается, что при «сильных» эмоциях гипоталамус дает почти максимальные разряды лод влиянием разрядов от ретикулярной формации и органов чувств, причем импульсы от последних поступают на ретикулярную формацию и гипоталамус не только непосредственно, но также через новую и димбическую кору. При этом дифференциация процесса активации и 'функций, принадлежащих различным кортикальным областям в нормальных физиологических условиях, снижается [581]. Возникающая вследствие этого «функциональная» декортикация не является результатом «кортикального конфликта», ведущего к высвобождению подкорки, как предполагает Дарроу [163], а зависит от чрезмерного возбуждения системы гипоталамуса, которое несовместимо с дифференцированным действием коры, необходимым для внимания [7] и высших психических процессов.
При дальнейшем усилении возбуждения гипоталамуса возникают невротические явления.
Гипоталамическая система
при экспериментальных неврозах
Так называемым экспериментальным неврозам предшествуют и их сопровождают глубокие изменения эмоционального поведения и вегетативной активности. Бели применяются относительно мягкие средства воздействия и длительное невротическое состояние не развивается, условный раздражитель вызывает сначала типичные симптомы возбуждения симпатической системы, затем возникают отчетливые парасимпатические явления, например рвота [510]. В гл. XVI было показано, что три таких явлениях, как тошнота .и рвота, доминирует пара-
544 ГЛАВА XXV
симпатическая система, а эрготропная система тормозится. Этот сдвиг вегетативного равновесия, сопровождающий тяжелые эмоциональные расстройства, можно рассматривать как а-омеастатичеокую реакцию: снижается реактивность в отношении всех окружающих раздражителей, и особенно тех, которые вызывают серьезные сдвиги.
Если условный раздражитель перестает подкрепляться, рвота -исчезает и восстанавливается нормальное равновесие в вегетативной системе. Вместе с тем, если на некоторое время насильственно подавляется соответствующая реакция на условный пищевой раздражитель или если используются другие способы индукции невроза, то возникают патологические условные рефлексы в форме общего возбуждения или торможения. При этих условиях обычно возникают 'расстройства вегетативных функций; нарушения сердечного ритма могут сохраняться годами [285]. Очевидно, при развитии у животного тяжелого эмоционального расстройства возникают либо очень сильные симпатические реакции, либо парасимпатические. В любом случае все поведение в целом становится патологическим.
Важно подчеркнуть, что внешние факторы, вызывающие значительное повышение реактивности симпатической системы, такие, как электрошок, инсулиновая кома и метразоловые судороги, ведут к сходным изменениям условных рефлексов. Несколько таких воздействий может привести к восстановлению ранее заторможенных условных рефлексов [355, 463], но серия из 10 или более приводит к устранению условной реакции страха [495] или к парадоксальным изменениям реакций на условные и дифференцировочные раздражители [764]. Исходя из этого, мы полагаем, что патологическое поведение возникает вследствие вегетативных нарушений на гипота-ламичееком уровне. Они характеризуются чрезмерно сильными симпатическими разрядами или нарушениями вегетативного баланса, возникающими под влиянием конфликтной ситуации или шоковой «терапии». Можно считаггь, что эти процессы причинно связаны и что изменения поведения в значительной степени зависят от изменения гипоталамо-кортвкальных разрядов.
ТЕОРИИ ЭМОЦИЙ
ПРОЦЕССЫ ВОЗБУЖДЕНИЯ И ТОРМОЖЕНИЯ В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ
Наше стремление подчеркнуть важное значение гипоталамуса и висцерального мозга для эмоций не исключает рассмотрения активации нервного аппарата в более широком аспекте.
Важные открытия последних лет показывают, что некоторые части центральной нервной системы оказывают диффузное возбуждающее или тормозное влияние на спинной и головной мозг. Восходящая активирующая ретикулярная система, описанная Мэтуном и другими [582, 584, 860, 861], которая при раздражении вызывает бодрствование и десинхронизацию в ЭЭГ, может служить классическим примером возбуждающей системы. Сходные эффекты можно получить с заднего отдела гипоталамуса ,и различных частей висцерального мозга. Рефлекторное возбуждение гипоталамуса через болевые и проириоцептивиые нервы [59] или высвобождение заднего отдела гипоталамуса из-под ограничивающих влияний барорецепторов синуса аорты аналогичным образом вызывает реакцию пробуждения в ее поведенческом и электроэнцефалографическом выражении [674]. Диффузная активация .коры, сопровождаемая реакцией пробуждения, возникает также при раздражении медиальных ядер таламуса и хвостатого ядра, если частота раздражения достаточно высока (100 имп/сек). Поскольку эти структуры усиливают также движения, вызываемые раздражением моторной коры, и повышают спинномозговые рефлексы [702], можно сказать, что в мозге существует возбуждающая система, к которой относятся корковые и подкорковые структуры и которая активирует сенсорные и моторные системы. Это в равной мере относится к реакции пробуждения и к эмоциональным реакциям: они могут быть вызваны с корковых и подкорковых структур и усиливают движения [671] и сенсорные процессы (восприятие [307]).
Напротив, существует тормозная система, снижающая двигательную активность и уровень активности коры. К ней относится ретикулярная формация продолговатого мозга, оказывающая тормозное влияние на спин-
18-517
ГЛАВА XXV
|номозго1вые рефлексы [617]. Кроме того, известно, что некоторые ядра мозга, например интраламинарные ядра ташамуса [186] и хвостатое ядро [830], оказывают синхронизирующее влияние на кору больших полушарий и подкорковые структуры, особенно при низкочастотном I раздражении или раздражении одиночными ударами. При этих условиях раздражение таламуса вызывает
(«реакцию остановки» (подобную временной потере сознания во время эпилептического припадка); раздражение хвостатого ядра снижает спонтанную активность [9] и подавляет движения, вызванные раздражением коры [640]. Раздражение гиппокамиа током низкого напряжения [507] и переднего отдела гипоталамуса [915] также может вызвать синхронизацию в коре. Последние работы показали еще более широкое распространение тормозной системы в мозге. Исследования ЭЭГ животных после перерезки ствола мозга кпереди от места вхождения V пары черепномозговых нервов (претригеми-нальный оредиемостовой препарат) показали, что синхронизирующие влияния могут возникать в каудальной части ствола мозга [156, 624], возможно в тормозном центре продолговатого мозга, по терминологии Мэгуна и Райнеса [617].
Значение этой работы очевидно из того факта, что сон можно вызнать не только раздражением гипниген-ной зоны Гесса, тесно связанной с интраламинарными ядрами таламуса, но также и переднего отдела гипоталамуса [34] и даже ретикулярной формации среднего мозга [139]. Интересно, что раздражение одного и того же участка в ретикулярной формации вызывает у обезьян зевоту и сон при напряжении 0,2 в, пробуждение при 0,4 в и выраженную паническую реакцию с признаками максимального возбуждения симпатической системы при 0,65—0,9 в [727]. Характер раздражения часто более важен, чем место его нанесения. Так, низкочастотное раздражение хвостатого ядра и интраламииарных ядер таламуса вызывает в коре синхронизацию, а высо-. кочастотпое — десинхронизацию.
Интересно, что процессы возбуждения сопровождаются признаками активации симпатической системы, тогда как тормозные состояния связаны с признаками
ТЕОРИИ ЭМОЦИИ
повышенной парасимпатической активности [394, '„926]. Эти данные показывают, что в функциональном отношении (Представления об эрго- и трофотропной системах могут быть расширены за пределы гипоталамуса. Несомненно, смена сна и бодрствования, а также различные эмоции сопровождаются сдвигом равновесия между возбуждающей и тормозной системами, поскольку между передним >и задним отделами гипоталамуса существуют реципрокные отношения, причем приятные эмоции в основном, очевидно, связаны с передним отделом гипоталамуса, а неприятные главным образом с задним. Яркой иллюстрацией этого может служить тот факт, что у кошки поглаживание по спине, вызывающее приятные ощущения, сопровождается синхронизацией в коре (подобно тому что имеет место при раздражении переднего отдела гипоталамуса), повышением возбудимости переднего отдела гипоталамуса и торможением тонических разрядов гамма-эфферентной системы [916]. Эти три явления объяснимы как результат парасимпатической настройки гипоталамуса и сопутствующего реципрокно-го торможения его симпатического отдела.
Поскольку эмоциональные расстройства часто приводят к грубой патологии, возникает вопрос, не ограничивает ли тормозная система: активность возбуждающей системы, и наоборот? Чтобы ответить на этот вопрос, надо проанализировать реактивность тормозной системы при разных уровнях активности возбуждающей системы.
Тормозная система при различных уровнях активности гипоталамуса и ретикулярной формации)
Ценными показателями реактивности тормозной системы (см. [482]) служат пороговая величина одиночного электрического раздражения, вызывающего разряд веретена (как во время сна), и порог низкочастотного раздражения, вызывающего реакцию вовлечения (появление и исчезновение синхронизированных корковых потенциалов большой амплитуды). Сходным образом чувствительность диффузной возбуждающей системы можно is*
ГЛАВА XXV
оценить по порогу раздражения, активирующего ретикулярную формацию или задний отдел гипоталамуса. Эта диффузная система дает разряды возрастающей интенсивности ,по мере перехода организма от сна к бодрствованию, .вниманию и, наконец, к эмоциональному возбуждению [579]. При определении порога возникновения веретен на фоне возрастающего уровня активности гипоталамуса и ретикулярной формации обнаруживается, что дорог возрастает параллельно (фиг. 55 и 56). Напротив, при снижении уровня активности заднего отдела гипоталамуса локальными разрушениями или инъекциями барбитуратов в эту структуру порог вызова веретен снижается.
Усиление гилоталамо-кортикальных разрядов под влиянием СОг или подпороговых доз метразола повышает порог вызова веретен. Снижение этих разрядов при введении таких фармакологических препаратов, как барбитураты (фиг. 57) .и хлорпромазин, или .под действием гипотермии понижает порог вызова веретен [892]. Эти результаты весьма постоянны независимо от того, используется ли в качестве индикатора состояние тормозной системы веретен или реакция вовлечения. При этом несущественно, производится ли проба на хвостатом ядре [891, 892] или «а интраламинарных ядрах та-ламуса [535], неважно также, чем вызвано изменение состояния заднего отдела гипоталамуса или ретикулярной формации —прямым раздражением, болевыми воздействиями или изменением внутренней среды (вдыхание углекислого газа). О роли состояния заднего отдела гипоталамуса свидетельствует тот факт, что значительное снижение порога раздражения хвостатого ядра при понижении температуры тела полностью исчезало после локальных разрушений заднего отдела гипоталамуса [325].
Эксперименты показывают, что тормозная система, представленная хвостатым ядром, которое при раздражении дает эффект успокоения [9], устраняет агрессивность [177], снижает или тормозит движения, вызываемые раздражением коры [640], и даже судороги [898], не оказывает ограничивающих влияний на возбуждение ретикуло-гипоталамо-кортикальной системы.
А

О 2 h 6 8 Ш
Напряжение тока. раздражающего гипоталамус,в
Фиг. 55. Повышение порога раздражения хвостатого ядра (одиночные импульсы длительностью 1 мсек) на фоне раздражения заднего гипоталамуса (0,5 .мсек, 120 имп/сек) "1891].
/ — раздражение заднего отдела ипсилатераль-
иого гипоталамуса. // — раздражение заднего
отдела контралатсральибго гипоталамуса.

-О I 2 3 4 ,
Напряжение тока, раздражающего ретикулярную формацию, в
Фиг. 56. Повышение порога раздражены» хвостатого ядра (одиночные импульсы длительностью 1 мсек) на фоне раздражения ретикулярной формации (0,5 мсек. 120 имп/сек) [891].
ГЛАВА XXV
Причина этого факта, очевидно, заключается в близкой связи между реакцией пробуждения и эмоциями. Мы знаем, что между сном и бодрствованием, так же как между передним и задним отделами гипоталамуса,

Ьбв */45а иЗв Ьй,5о
А

U3 в Tjjg
б _
Фиг. 57. Электроэнцефалограммы, иллюстрирующие влияние пенто-тала на порог раздражения хвостатого ядра [892]. Опыты на курарезироваиной кошке (вес 4,1 кг). А. Контрольный порог 14.5 в. Б, Порог 11 в после двух инъекций пеитотала (каждая по 2 мг/кг) зя 5,5 и 3,5 мин до записи. 1 — левая передняя сигмовидная извилина; 2— левая краевая извилина; 3 — левая средняя супра-сильвиева извилина.
с которых можно вызвать эти состояния, существуют рещшрокные отношения. Более того, синхронизация кортикальных потенциалов во время сна сопровождается снижением активности ретикулярной формации [931]. Эти реципрокные отношения между возбуждающей и тормозной системами сохраняются при различных уровнях активности гипоталамуса и ретикулярной формации, что обнаруживается по прогрессивно снижающемуся тормозному влиянию (синхронизация) хвостатого ядра на фоне возрастания возбуждения ретикуло-типотала.мической системы, л наоборот [891].
теории эмоции
gsj
Неспособность тормозной системы ограничивать чрезмерное эмоциональное возбуждение можно объяснить с телеологических позиций, если .принять во внимание биологическое значение эмоций.
Воды эмоций, при которых усилена активность заднего отдела гипоталамуса, прежде всего связаны с нападением и бегством, и любой гомеостатический механизм, ■который бы имел тенденцию к ограничению этих действий, представлял бы опасность для организма. Напротив, р этих условиях наблюдается эрготроплая реакция, характеризующаяся синэргичным действием соматической нервной системы и симпатического отдела гипоталамуса. Движения, вызванные раздражением коры, усиливаются гипоталамусом [671], причем это облегчение осуществляется как на кортикальном, так и на спинальном уровнях [314, 671, 702]. Кроме того, -гормон мозгового .вещества надпочечников замедляет (Наступление мышечного утомления.
Тем не менее торможение играет важную роль в некоторых эмоциональных .процессах. При реакциях страха и ужаса тонус скелетной мускулатуры снижается; человек в этих условиях не способен двигаться, а животное . как бы «застывает»; однако признаки симпатической активности совершенно очевидны. Подобный синдром — со-четание симпатического возбуждения с расслаблением 1 мышц — можно вызвать раздражением миндалевидного I ядра [906], а также лоясной извилины [848]; при удалении , этой области реакция страха исчезает [929]. Однако это I трудно объяснить возбуждением заднего отдела гипоталамуса, приводящего к зрготропной активности, т. е. к симпатическому возбуждению и активации скелетной мускулатуры. На основании анализа можно предположить, что относительная роль нейрогенных и гормональных (адреномедуллярцых) компонентов симпатического возбуждения при разных эмоциональных состояниях различна. При реакциях, подобных ярости («внешняя ярость» «anger out» [271]), преобладает нейрогенная активность, регулируемая гипоталамусом, тогда как при «внутренней ярости» господствует гормональная фаза, регулируемая ,в основном продолговатым мозгом [323]. Этим объясняется тот факт, что циркуляторные реак- |
5S2 ГЛАВА XXV
i ции и выделение катехоламинов указывают на преобла' дание норадреналина при ярости и адреналина — при таком эмоциональном состоянии, как ужас [150].
АНТАГОНИЗМ МЕЖДУ НОВОЙ И ДРЕВНЕЙ КОРОЙ
* * * * 1 Подробности первых работ, свидетельствующих о снижении порога «ярости» при… ТЕОРИИ ЭМОЦИЙ 553L
5G8
ГЛАВА XXVI
5. Влияние вызванных реакций гнева и тревоги (которые, как известно, изменяют реакцию на мехолил [150]) на действие норадреналина.
Дельта-волны в ЭЭГ и вегетативная нервная система
Моррис [667] показал, что появление медленных волн в ЭЭГ после гипервентиляции более выражено у лиц с положительным глазо-сердечным рефлексом, чем у тех, у кого этот парасимпатический рефлекс отсутствует. Известно, далее, что дельта-волны чаще появляются при гипервентиляции у лиц, относящихся к группе III (по мехолилу). Хотя мы не располагаем обширным материалом, результаты указывают на .более выраженную тенденцию к выявлению медленных волн в коре у. лиц с низкой симпатической и высокой парасимпатической реактивностью.
Мы предполагаем повторить эти важные наблюдения на большой группе здоровых людей одного возраста, предварительно проведя у них пробы с мехолилом и нор-адреналином [354, 682].
Проба с норадреналином также легко поддается количественной оценке, как и проба с мехолилом, и более удобна, чем рефлекс торможения сердца. Предлагаемые серии экспериментов позволят выявить, усиливается ли проявление медленных волн при пробе на гипервентиляцию на фоне повышения парасимпатической реактивности (определяемой по пробе с норадреналином) и(или) при снижении симпатической реактивности (определяемой по пробе с мехолилом). Теоретически можно ожидать усиления тенденции к появлению медленных волн у лиц, у которых равновесие вегетативной системы сдвинуто в сторону преобладания парасимпатической системы (значительное замедление частоты сердечных сокращений для данного уровня кровяного давления при пробе с норадреналином и принадлежность к группе III при пробе с мехолилом). В этих экспериментах необходимо следить за постоянством уровня сахара крови.
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 569
Фармакологические факторы, влияющие на вегетативное равновесие
Далее мы предлагаем изучить сдвиг равновесия вегетативных систем з симпатическую или парасимпатическую сторону с помощью таких веществ, как амфетамин (фенамин) и хлорпромазин (аминазин) или пентотал. В этих условиях можно исследовать значение принципа реципрокной иннервации путем применения проб с ме-холилом и норадреналином. Для количественной оценки этих влияний у психически больных можно воспользоваться пробами с мехолиХяом и норадреналином, а также холодовой пробой и кожно-гальваническим рефлексом.
В целях выяснения корреляций между вегетативными сдвигами и психическими реакциями рекомендуется провести с мехолилом и норадреналином до и во время применения мескалина и LSD. Ринкель и др. [760] уже отметили «несколько менее выраженную реакцию парасимпатической системы на мехолил» при действии LSD; это, очевидно, означает, что снижение кровяного давления было менее выраженным и что (с нашей точки зрения) произошел ожидаемый сдвиг в сторону преобладания симпатической системы. Как изменяется парасимпатическая реактивность, неизвестно. Можно ожидать, что эти исследования помогут выяснить физиологический смысл известного утверждения о том, что личность испытуемого играет важную роль в определении его реактивности на транквилизаторы и психоактивные вещества.
Рефлексы с барорецепторов и восприятие
Ранее было показано, что повышение активности заднего отдела гипоталамуса сопровождается повышением реактивности специфических сенсорных проекционных зон по отношению к афферентным раздражителям [301, 307]. Этим, возможно, объясняется тот факт, что в условиях раздражения гипоталамуса наблюдается реакция на подпороговые сенсорные раздражители [310]. Можно ожидать противоположных результатов, если реактивность заднего отдела гипоталамуса и, следовательно,
ГЛАВА XXVI
интенсивность гипоталамо-кортикальных разрядов снижена. Это можно получить у экспериментального жи-вотного путем введения норадреиалина, который повышает тормозное действие рефлекторных влияний с барорецепторов на задний отдел гипоталамуса [76, 674]. Было бы интересно исследовать, нельзя ли по этому механизму вызвать у человека изменения кортикальных функций, связанные с восприятием. Предлагается исследовать следующие влияния внутримышечного введения норадреиалина:
1. На сенсорное восприятие. Для этой цели следует определить порог ощущения боли и порог восприятия звука, а также критическую частоту слияния мельканий.
2. На болевую реакцию, оцениваемую по латентному периоду и величине кожно-гальваиического рефлекса. В качестве болевого раздражителя можно использовать электрическое раздражение кожи кисти. Холодовая проба также может быть использована для определения симпатической реакции на болевой раздражитель. Надо проверить, подтверждают ли эксперименты, перечисленные в пп. 1 и 2, предположение об уменьшении восходящих и нисходящих разрядов гипоталамуса во время фазы повышения кровяного давления (усиление рефлексов с барорецепторов).
Айзенк, Грейнджер и Бренгельманн [234] показали, что некоторые процессы восприятия количественно различны в норме, при невротических и психических заболеваниях. Весьма вероятно, что различия в эмоциональной реактивности играют ведущую роль б этих явлениях.
ПСИХОНЕВРОЛОГИЧЕСКИЕ ПРОБЛЕМЫ ВЕГЕТАТИВНОЙ РЕАКТИВНОСТИ И РАВНОВЕСИЯ
Если правильно предположение о том, что пробы с мехолилом и норадреналином отражают уровень симпатической и парасимпатической реактивности и характеризуют состояние вегетативного равновесия на уровне гипоталамуса, то систематическое использование этих проб должно помочь решению многих клинических проблем. Мы рекомендуем использовать их в сочетании с электромиографией, холодовой пробой и кожно-гальза-
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 571
ническим рефлексом. Мы уже писали в своих статьях и книгах [296, 300, 306, 310, 313] о том, что было бы чрезвычайно интересно сопоставить вегетативные параметры, полученные при обследовании здоровых людей и больных различными формами неврозов и психозов, а также провести параллели между психическим и физиологическим статусом при изменениях клинического состояния, происходящих спонтанно или под влиянием терапевтических воздействий.
Мы предлагаем рассчитанный на длительное время план обследования, который должен послужить созданию профилактического направления в психиатрии. У тысячи или более студентов надо провести пробы с мехолилом и норадреналином, а также ряд психологических тестов. Через соответствующие периоды времени следует проверять состояние их психики. При таком исследовании, возможно, удастся выявить вегетативные сдвиги, предшествующие возникновению психических расстройств.
Кроме того, следует изучить вегетативные сдвиги, наблюдаемые при острых и хронических формах шизофрении. Прежние исследования Шнейдера и др. [804] показали, что депрессанты снижают симпатическую реактивность у лиц с гиперактивностыо симпатической системы (реакции на мехолил соответственно группе I) и повышают ее у лиц с гипоактивностью симпатической системы (реакция на мехолил соответственно группе III). Этот результат противоречит физиологическим исслет дованиям, в которых было показано, что проба с мехолилом выявляет сниженную симпатическую реактивность при введении депрессантов в кровь или непосредственно в гипоталамус [356, 357]. Можно ожидать, что при введении пентотала или хлорпромазина произойдет сдвиг-реакции на мехолил в сторону снижения симпатической реактивности, что, возможно, будет связано с повышением парасимпатической реактивности, обнаруживаемым пробой с норадреналином. Противоположный результат ожидается при использовании веществ, стимулирующих симпатическую систему (амфетамин, мескалин, LSD). Более слабая реакция на LSD у больных хронической шизофренией по сравнению с нормой —это факт, кото-
ГЛАВА XXVI
рый должен лечь в основу систематического исследования для выяснения вопроса, изменена ли в той же мере подвижность вегетативной системы (парасимпатической и симпатической) или между психологическим и физиологическим эффектами имеется расхождение. Весьма возможно, что методы, предложенные для воздействия на вегетативные центры здоровых людей (вдыхание СОг, инсулиновая гипогликемия), позволят обнаружить более тонкие различия между нормальными людьми и больными с острыми и хроническими психозами.
Мескалин, LSD и вегетативная нервная система
В гл. X мы указывали, что психотические реакции вызывает только сам LSD и те его производные, которые оказывают возбуждающее действие на симпатическую нервную систему. Влияние LSD на настроение (эйфория у здоровых людей и состояние тревоги у больных шизофренией [481]), тот факт, что хлорпромазин полностью снимает психотические реакции, вызванные LSD и мес-калином, а также появление вегетативных симптомов, обычно предшествующих психическим расстройствам, подтверждают наше представление о том, что в основе психотических симптомов, вызываемых этими веществами, лежит возбуждение симпатического отдела гипоталамуса, а следовательно, количественные и качественные "изменения гипоталамо-кортикальных разрядов. Если эта точка зрения правильна, можно ожидать, что указанные симптомы зависят от равновесия и реактивности систем гипоталамуса. Поэтому представляет интерес исследование корреляции результатов проб с мехолилом и норадреналином с типом изменений, вызываемых LSD и мескалином у здоровых людей и психически больных, или, конкретнее, установить, как реагируют на LSD и мескалин лица, относящиеся к разным группам по реакции на мехолил.
Различный характер изменений психики у одного и того же лица при повторных пробах [131, 481], возможно, объясняется сдвигами эмоционального состояния и реактивности систем гипоталамуса. Поэтому пробы с
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ
LSD и мескалином следует проводить в условиях направленного изменения эмоционального состояния, как это делали Коэн и Сильвермен [150], которые показали, что выделение норадреналина и реакция на мехолил изменяются параллельно. Эти эксперименты должны показать, какие психические сдвиги возникают под влиянием LSD при условии, когда активность симпатической системы изменена в результате реакции тревоги или агрессии.
Хох [481] наблюдал, что для того чтобы вызвать психические сдвиги у больных хронической шизофренией, нужны более высокие дозы указанных веществ, чем при острой шизофрении и у здоровых людей. Очевидно, такие различия аналогичным образом проявляются и на физиологическом уровне, поскольку LSD повышает секрецию норадреналина у больных, находящихся в состоянии депрессии, но не оказывает подобного влияния при хронической шизофрении [222]. На этом основании мы высказываем предположение о том, что при хронической шизофрении реактивность симпатической системы на уровне гипоталамуса снижена. Предполагается изучить корреляцию между влиянием амфетамина и пенто-тала на пробу с мехолилом и эффективностью LSD в вызове психических сдвигов.
Сущность седативного порога
Шагасс и сотр. [818—823] в ряде исследований установили, что стабильным показателем состояния мозга может служить седативный порог, который позволяет выявить характерные различия между здоровыми людьми, невротиками и психически больными. Первоначально этот порог определялся путем введения с постоянной скоростью амобарбитала натрия и установления того момента, когда речь становилась смазанной, а на ЭЭГ лобной области появлялись характерные изменения в амплитуде быстрых (15—30 в 1 сек) потенциалов. Позднее был принят более простой показатель — возникновение сна. При внутривенном введении пентотала порог определяли количеством препарата, которое необходимо ввести со скоростью 0,5 мг/кг за 30 сек, чтобы исчезла
ГЛАВА XXVI
реактивность в отношении словесных раздражителей. С помощью этого метода было показано, что сдвиги в эмоциональном состоянии сопровождаются изменением этого порога. Гнев, тревога и тоска повышают этот порог. Характерные различия при разных видах невроза показаны на фиг. 58. Для истерии характерен минимальный седативный порог, а для состояния тревоги — максимальный.
Применение плетизмографии для исследования состояния вегетативной нервной системы в норме, при истерии и при состояниях тревоги [903] выявляет аналогичные соотношения. Состояние тревоги, очевидно, свя-

Фиг. 58. Соотношение высоких (4 мг/кг и более) и низких (3,5 мг/кг и менее) порогов успокоения у больных с различными психоневрологическими заболеваниями [819].
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 575
зано с преобладанием симпатической системы, а истерия— с преобладанием парасимпатической.
Характерные различия были обнаружены между невротическими и психическими депрессивными состояниями— в первом случае седативный порог повышен, во втором понижен [551]. Седативный порог повышается по мере увеличения числа сеансов электрошока. Шагасс представил экспериментальные доказательства гипотезы о том, что седативный порог является показателем уровня возбудимости мозга.
Возникает вопрос: чем объясняются различия в возбудимости мозга при различных клинических состояниях? По-видимому, седативный порог меняется при эмоциональных сдвигах. Он возрастает при состоянии гнева и тревоги и понижается при наступлении покоя. Эти данные показывают, что состояние гипоталамуса и особенно уровень гипотала.мо-кортикальных разрядов связаны с величиной седативного порога. В пользу такого объяснения свидетельствуют и следующие данные: 1) повторные сеансы электрошока повышают седативный порог; 2) вероятность терапевтического эффекта электрошока снижается, если седативный порог возрастает [819]; это согласуется с нашим мнением, согласно которому электрошок — процедура, возбуждающая в основном центральные симпатические структуры, — может помочь лишь больным с пониженной (но не с повышенной) симпатической реактивностью; 3) при депрессивных состояниях электрошок приводит к облегчению состояния, но повышает седативный порог тогда, как при состояниях тревоги успешная психотерапия и нормализация сопровождаются снижением этого порога [819]. Это позволяет считать, что низкая симпатическая реактивность при депрессивных состояниях под влиянием электрошока повышается до нормального уровня, тогда как патологически высокая реактивность симпатических центров при возбужденных состояниях под влиянием седативной терапии и психотерапии снижается до нормального уровня. Напомним читателю, что нарушения равновесия систем гипоталамус: ведут к параллельно возникающим изменениям восходящих и нисходящих разрядов.
576 ГЛАВА XXVI
Мы указали в гл. XXV, что реактивность центральной тормозной системы, измеряемая порогом возникновения одиночного веретена или реакции вовлечения при раздражении таламуса или хвостатого ядра, зависит от степени активности возбуждающей системы, представленной в основном ретикулярной формацией и гипоталамусом. Нам кажется, что методом Шагасса можно выявить и состояние этих систем. Синхронивацию (и сон) можно вызвать инъекцией барбитуратов или одиночным электрическим раздражением хвостатого ядра, но в любом случае результат определяется уровнем ретикуло-гипоталамо-кортикальных разрядов. Бели это допущение правильно, то можно ожидать следующих отношений:
1. У психически здоровых людей одного возраста с различной реакцией на мехолил (соответствующей группам I и III) должен наблюдаться соответственно высокий и низкий седативный порог. Этого следует ожидать, так как эксперименты на животных, описанные в гл. V, показали, что реакция на мехолил, соответствующая группе III, указывает на снижение симпатической реактивности гипоталамуса и соответствующее снижение интенсивности кортикальных разрядов.
2. У больных с депрессивными состояниями, которые под влиянием лечения электрошоком переходят из группы III по мехолилу в группу II, седативный порог должен после лечения повыситься'.
3. При обследовании одного и того же лица в состоянии покоя и в агрессивном состоянии в последнем случае должна быть обнаружена усиленная секреция норадреналина, повышение седативного порога и сдвиг реактивности на мехолил, свидетельствующий о повы-* * * *
1 Шагасс [819] упомянул о там, что он и Слоан сравнивали величину седативного порога я результаты пробы с мехолилом у 30 больных и не обнаружили корреляции между этими тестами. Тем >не менее такое исследование нужно позторить (включив в него пробу с норадреналином как показатель парасимпатической реактивиости и холодовую пробу и другие пробы, отражающие состояние симпатических центров). Следует взять одинаковое число обследованных каждой группы и количественно оценить результаты, воспользовавшись разработанными ;методами [354].
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 577
шении возбудимости симпатической системы (т. е. переход из группы III в группу II или I или из группы II в группу I). Аналогичный сдзиг можно наблюдать и в том случае, если первая проба проводится при нормальном уровне сахара крови, а последующая при умеренной степени инсулиновой гипогликемии, поскольку в последнем случае центральная симпатическая реактивность возрастает, а некоторые данные Хилла и др. [463] показывают, что гипоталамо-кортикальные разряды в этих условиях оказываются повышенными.
Проведение такого обследования у здоровых людей, при неврозах и у психически больных, принадлежащих к разным возрастным группам, поможет выяснить, не происходит ли диссоциации между интенсивностью восходящих и нисходящих гипоталамических разрядов при физиологических (возраст?) или патологических (хронические психозы?) условиях.
Механизм пробы с пентоталом
Гольдман [387] воспользовался пентоталом, чтобы отдифференцировать больных шизофренией от других психически больных и от здоровых людей. Он обнаружил, что при внутривенном введении этого вещества у больных шизофренией появляются быстрые бета-волны и асимметричные разряды'. Поскольку записи показывают выраженные различия в электрической активности, для получения количественных данных и объективизации результатов следует использовать интегратор. Ряд фактов свидетельствует о тесной связи между результатами пробы с пентоталом и седативным порогом Шагасса и о том, что повышенная электрическая активность зависит от интенсивности гипоталамо-кортикаль-ных разрядов.
Правильность такой интерпретации подтверждается следующими клиническими фактами:
1. Клиническое улучшение, наступающее у больных шизофренией под влиянием резерпина или фенотиазина,
* * * *
1 В этом случае тест считается положительным.
19-517
ГЛАВА XXVI
сопровождается менее выраженной реакцией па введение пентотала; высокая активность коры соответствует клиническому ухудшению, которое может быть спонтанным или наступать в результате прекращения упомянутого выше успешного медикаментозного лечения.
2. Если при применении тофраиила у больных с маниакально-депрессивным психозом депрессивная фаза сменяется маниакальной, то проба с пентоталом оказывается положительной.
3. При применении антидепрессантов, относящихся к стимуляторам симпатических центров (как мы указывали в гл. XXIII), проба с пентоталом становится положительной.
Эти факты отчетливо показывают, что проба с пентоталом становится положительной при повышении реактивности симпатических центров. Следовательно, при этом должен происходить сдвиг реакций на мехолил к типу, характерному для группы I. Исходя из этого, мы рекомендуем определить, при каких условиях сдвиг равновесия вегетативных центров гипоталамуса в сторону преобладания симпатического отдела (о чем указывает соответствующее изменение реакции на мехолил) сопровождается положительной пентоталовой пробой. В случае если указанное уточнение оценки результатов пробы с помощью электронного интегратора окажется успешным, надо установить следующее:
1. Обнаруживаются ли у здоровых лиц одного возраста, принадлежащих к различным группам по мехо-лилу, соответствующие различия при пробе с пентоталом.
2. Не сопровождается ли наступающий под влиянием резерпина или фенотиазина переход из группы I по ме-холилу в группы II и III одновременной сменой положительной пробы с пентоталом на отрицательную.
Снятие депрессии путем соответствующих фармакологических воздействий или терапии электрошоком сопровождается параллельными изменениями реакции на пентотал и мехолил: усиливается положительный эффект пробы с пентоталом и наблюдается переход в группу I по мехолилу. Можно ожидать также противоположных сдвигов при пробе с норадреналином.
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 579
Дополнительные физиологические эксперименты на животных
Мы полагаем, что седативный порог, .проба Годьдма-на с пентоталом и появление дельта-волн при гипервентиляции имеют один общий момент — они зависят от интенсивности гипоталамо-кортикальных разрядов. Чтобы исследовать правильность этой гипотезы, предлагается провести эксперименты на кошках с применением эфирного наркоза, местной анестезии и кураре (искусственное дыхание). Предлагается регистрировать электрокардиограмму и кровяное давление при введении раздражающих электродов в передний или задний отделы гипоталамуса, как это было сделано в работе Гель-горна и др. [310, 358, 674]. Влияние определенного уровня гипервентиляции на ЭЭГ в контрольных условиях следует сравнить с тем же эффектом после повышения активности заднего отдела гипоталамуса (и, следовательно, интенсивности гипоталамо-кортикальных разрядов) путем введения в гипоталамус стрихнина (или метразола) или после ее снижения введением пентотала (или разрушением высокочастотным током). Следует также испытать влияние односторонних инъекций и разрушений, поскольку они могут привести к односторонним изменениям реактивности гипоталамо-кортикальной системы, причем интактная сторона будет служить контролем [549]. Можно ожидать, что воздействия, снижающие активность заднего отдела гипоталамуса, ускорят появление дельта-волн при гипервентиляции и усилят их. Повреждения переднего отдела гипоталамуса или вент-ро-медиального ядра таламуса, напротив, должны путем высвобождения заднего отдела гипоталамуса замедлить ч ослабить появление медленных волн при гипервентиляции.
Весьма вероятно, что эти воздействия приведут также к соответствующим изменениям влияния пентотала на потенциалы коры. В качестве критерия вместо словесных ответов можно использовать сдвиг частоты'электроэнцефалограммы, оцениваемый по интегратору. Можно ожидать, что при постоянной скорости введения пентотала максимальная величина медленных волн будет
19*
ГЛАВА XXVI
достигнута быстрее при снижении активности заднего отдела гипоталамуса, и наоборот. Интересно также установить следующее:
1. Характер влияния вдыхания возрастающих концентраций СОг (3—10%) у .кошек, находящихся под поверхностным наркозом (см. [302, 303, 313, 339]), на синхронизирующее ЭЭГ действие пентотала, вводимого с постоянной скоростью. (Можно ожидать, что на фоне вдыхания С02 для изменения ЭЭГ потребуется больше пентотала.)
2. Влияние повреждений переднего и заднего отделов гипоталамуса на указанное действие пентотала.
Если наша гипотеза правильна, то при разрушении переднего отдела гипоталамуса порог действия пентотала повысится, а при разрушении заднего отдела гипоталамуса понизится. Необходимо, конечно, проследить за тем, чтобы в контроле влияния гипервентиляции и внутривенного введения пентотала на ЭЭГ были воспроизводимыми и постоянными.
Вспомогательные проблемы
Теоретические соображения и гипотезы, изложенные в данной главе, приложимы к многочисленным проблемам. Читатель, который следил за нашими рассуждениями, сможет сам выбрать проблему и поставить соответствующие эксперименты. Поэтому мы ограничимся здесь перечислением некоторых направлений исследования, которые кажутся нам особенно многообещающими.
1. Исследования влияния лишения сна [32] показывают, что первоначально возникает фаза повышенной раздражительности, связанная с повышением частоты колебаний на ЭЭГ [897]. На более поздних стадиях отмечается ухудшение проводимости кожи, уменьшение тонуса мышц и падение температуры тела, что указывает на снижение реактивности эрготропной системы. Пробы с мехолилом и норадреналином позволяют выяснить характер изменений равновесия вегетативных систем и их влияний на активность коры в этих условиях.
2. Предлагается расширить изучение проб с мехолилом и норадреналином: а) поставить их у пожилых
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ
людей и выяснить корреляцию с психическим состоянием для того, чтобы определить не только характер физиологического равновесия вегетативных систем в старости, но также его возможное значение для возникновения старческих изменений психики; б) поставить эти пробы у больных эпилепсией, умственно отсталых, алкоголиков и лиц, совершивших сексуальные преступления; в) изучить влияние менструации и полового созревания на равновесие вегетативных систем с помощью этих проб.
3. Хотя Мориарти [664] убедительно показал, что лечение СО,2 наиболее эффективно при повышенной симпатической реактивности (группа I по мехолилу), не известно, связано ли это улучшение со снижением симпатической реактивности. Нет также данных относительно состояния парасимпатической системы в этих условиях. Применение проб с мехолилом и норадреналином до и после лечения поможет ответить на эти вопросы.
4. Применение проб с мехолилом и норадреналином вскоре после первого сеанса терапии электрошоком и по окончании курса лечения и их сравнение с результатами проб до лечения поможет выяснить природу физиологических сдвигов, происходящих под влиянием лечения. Эти пробы позволят выявить реактивность вегетативной системы в различных клинических группах в острых и хронических случаях. Аналогичные исследования следует провести на больных, леченных инсулиновой комой, чтобы выяснить физиологическую основу отмеченных клиницистами различий между этими двумя формами шока Г14].
5. В литературе имеются отрывочные сведения, указывающие на возможность сдвига равновесия вегетативных систем и возрастания парасимпатической реактивности, аналогичного состоянию «парасимпатической настройки», описанному в гл. V. Перечислим кратко эти наблюдения: а) у психоневротиков состояние эмоционального возбуждения сопровождается повышением кожной температуры, тогда как в норме наблюдается ее снижение [509]; б) при холодовой пробе в некоторых случаях шизофрении происходит снижение кровяного давления вместо повышения [409]; в) при депрессивных состояниях и шизофрении уровень сахара крови после электро-
ГЛАВА XXVI
шока повышается; если же пробу повторить на фоне поверхностного наркоза, то при шизофрении отмечается гипогликемия, а при депрессивных состояниях — незначительная гипергликемия [424]; г) при неврозах, сопровождающихся апатией, утомляемостью и т. д., отмечается тенденция к спонтанной гипогликемии [722а].
Было бы также интересно установить посредством проб с мехолилом и норадреналином, связаны ли парасимпатические реакции в этих случаях со сдвигом вегетативного равновесия. Пробы с мехолилом и норадреналином позволяют лишь отчасти выполнить основную цель данной книги, состоящую в изучении уровня возбудимости гипоталамуса и вегетативного равновесия. Более полное представление о состоянии гипоталамуса можно получить, если дополнительно определять концентрацию различных гормонов, зависящих от гипота-ламо-гипофизарного механизма, а также концентрацию норадреналина и адреналина в крови и моче. Например, мы до сих пор не знаем, сопровождаются ли изменения секреции стероидов, характерные для двух фаз маниакально-депрессивного психоза [121], соответствующим сдвигом равновесия вегетативных систем. Кроме того, было бы чрезвычайно важно исследовать влияние вегетативного равновесия( как описано выше) на выделение гормонов и неирогормонов, особенно при хронических психозах, и сравнить результаты с данными, полученными в аналогичных условиях в норме и при острых психозах.
Следует учитывать, что перечисленные выше клинические исследования преследуют две различные цели.
1. Проведение массовых проб с применением мехо-лила и норадреналина для установления корреляции между вегетативными функциями, с одной стороны, и характером заболевания' и вариациями психического поведения — с другой. Можно было бы примирить многочисленные противоречия, имеющиеся в литературе, подбирая группы, однородные по возрастному составу, тща-* * * *
1 Значительные различия реактивности на мехолил, очевидно, существуют при состояниях тревоги, при депрессии и гебефрснии [20а].
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 583
тельно учитывая условия, обеспечивающие четкую воспроизводимость результатов и количественную их оценку. Мы пока не можем ответить даже на сравнительно простые вопросы, такие, как влияние электрошока на вегетативную систему (на основании проб с мехолилом и норадреналином). Наличие корреляции между клиническим улучшением и изменением реактивности вегетативной системы признают одни авторы и отрицают другие. Со стороны ясно, что по крайней мере часть этих противоречий обусловлена недостаточной тщательностью проведения проб и клинического анализа.
2. Дальнейшее выяснение многочисленных специальных вопросов путем применения проб с мехолилом и норадреналином в экспериментах, исследующих психофизиологические зависимости, как в работах Лейси и Лейси [558, 559].
ПРОБЛЕМЫ ИЗ ОБЛАСТИ ВНУТРЕННИХ БОЛЕЗНЕЙ
Гипертония и функции вегетативной нервной системы
В гл. XIII мы показали, что эмоциональные расстройства, ведущие к длительным симпатическим разрядам, играют ведущую роль в возникновении гипертонии. Вместе с тем имеются наблюдения, которые, по-видимому, противоречат этому утверждению: 1) при резком наклоне головы (на 75°) секреция норадреналина у здоровых людей усиливается, а у больных гипертонией нет [877а]; 2) падение кровяного давления при резком наклоне выражено у больных гипертонией сильнее, чем у здоровых людей [877а]; 3) для гипертонии характерно длительное снижение кровяного давления при инъекции мехолила (группа III). Эти 3 наблюдения заставляют предположить, что реактивность симпатических центров снижена.
Мы примирили бы эти противоречия, если бы нам удалось показать, что при гипертонии ослабляются барорецепторные рефлексы. Снижение этих рефлексов способствует развитию гипертонии, ослабляет гомеоста-тическую регуляцию кровяного давления и, следовательно, объясняет эффекты, указанные в пп. 1 и 3.
Глава xxvi
Для проверки этой гипотезы предлагаются следующие исследования:
А. Обследование лиц с гипертонией (ранняя стадия) и лиц с нормальным давлением, предрасположенных к гипертонии, судя по семейному анамнезу.
Следует отобрать две группы молодых людей одного возраста: группу предрасположенных к гипертонии или с начальной стадией гипертонии и контрольную группу — и провести ряд следующих проб.
1. Проба с мехолилом для выявления частоты повышенной реактивности симпатической системы в обеих группах.
2. Комбинация проб с норадреналином и с мехолилом для выяснения различий в состоянии вегетативного равновесия между этими двумя группами.
3. Холодовая проба с количественной оценкой кровяного давления и частоты сердечных сокращений, а также влияния на интегрированные мышечные токи действия и кожно-гальванический рефлекс.
Можно ожидать, что эти пробы выявят повышенную реактивность эрготропной системы в первой группе (по сравнению с контрольной).
Б. Обследование лиц с гипертонией.
В этой группе следует сравнить результаты обследования лиц с умеренной гипертонией, длящейся в течение нескольких лет, с данными обследования людей с нормальным давлением того же возраста, применив пробы с мехолилом, норадреналином и холодовую пробу.
Можно ожидать, что в контрольной группе выявится параллелизм между результатами пробы с мехолилом и холодовой пробы. Наоборот, при гипертонии должна быть обнаружена выраженная симпатическая реакция эрготропной системы на болевое раздражение, применяемое в холодовой пробе, а проба с мехолилом должна соответствовать группе III. Такое расхождение результатов двух проб может служить подтверждением предположения о том, что для гипертоний характерно усиление центральных симпатических разрядов при снижении барорецепторных рефлексов. Далее, весьма вероятно, что при гипертонии прессорная реакция на норадреналин будет повышена вследствие сниженного
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 585
гомеостатического действия барорецепторцых рефлексов [385].
Гипоталамус и гипертония
В гл. XIII было показано, что эмоциональные расстройства способствуют развитию гипертонии у человека и что по крайней мере в одном исследовании [811] эти факторы привели к гипертонии и сердечно-сосудистым нарушениям у экспериментальных животных. Развитие методики хронического раздражения [216] и стереотак-сического метода [578] позволяет сопоставить влияние хронического раздражения заднего отдела гипоталамуса кровяное давление с результатами классических экспериментов Гольдблата и др. [384] по почечной гипертонии.
Язвенная болезнь
В гл. XIV мы привели данные, свидетельствующие о том, что язва желудка у человека и экспериментальных животных связана с повышенной активностью блуждающего нерва и сдвигом гипоталамического равновесия в сторону относительного преобладания парасимпатической системы. Далее было показано, что сильная активация симпатического отдела гипоталамуса связана с появлением агрессивных тенденций и секрецией нор-адреналина, а меньшая степень возбуждения гипоталамуса сопровождается появлением состояния тревоги и секрецией адреналина (гл. VII). Проведенные недавно исследования показали, что у больных с язвой желудка чаще наблюдается состояние тревоги и понижение секреции иорадреналииа [152]. Этого и следовало ожидать, если признать, что язвенная болезнь зависит от снижения активности симпатического отдела гипоталамуса. Эти важные исследования необходимо дополнить проведением проб с норадреналином и мехолилом. Можно ожидать, что при психическом стрессе, ведущем к повышению желудочной секреции, парасимпатическая реактивность гипоталамуса будет повышена, а симпатическая—понижена, т. е. проба с норадреналином даст высокий показатель, а реакция на мехолил будет соответствовать группе III.
ГЛАВА XXVI
Прочие проблемы
Добавим краткие замечания по поводу некоторых других проблем, касающихся лечения внутренних болезней. Мы лишь приведем примеры, а читатель сможет воспользоваться ими при разработке других, интересующих его проблем.
1. Уже давно установлено, что между уровнем активности щитовидной железы и симпатической нервной системы существует корреляция, в основе которой лежит экспериментально доказанная зависимость секреторной активности щитовидной железы от гипоталамуса [427, 428]. Проведение проб с мехолилом и иорадреналином при гипо- и гипертиреозе поможет нам глубже понять состояние равновесия вегетативных систем при этих патологических состояниях. Эти пробы помогут объяснить, почему у лиц с гипертиреозом внутривенное введение глюкозы вызывает более выраженную гипергликемию, чем у здоровых людей [45]. Не связана ли эта пониженная толерантность к глюкозе с гипореактивностыо парасимпатической (ваго-инсулииовой) системы? Ответ на этот вопрос может дать проба с иорадреналином.
2. В гл. XXIV упоминалось, что вегетативные изменения связаны с физическими явлениями, характерными для циклических заболеваний. И при этих заболеваниях целесообразно воспользоваться вегетативными пробами.
3. Доказано, что острая блокада каротидного синуса новокаином ведет к извращению действия инсулина: вместо гипогликемии возникает гипергликемия [532]. Эти наблюдения позволяют сделать предположение, что подобное извращение реакции на инсулин связано со сдвигом равновесия вегетативных центров вследствие блокады нервов каротидного синуса. Для дальнейшего анализа предлагается провести подобные эксперименты на неиаркотизированных кроликах или собаках, удалив у них каротидный синус. Если эти эксперименты подтвердят наблюдения, сделанные на людях, то можно будет установить, какое влияние окажет на действие инсулина перерезка чревного нерва, удаление мозгового вещества надпочечников или симпатическая блокада.
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ
5S7
4. Известно, что при аддисоновой болезни и ревматоидном полиартрите реакция на мехолил повышена [704]. Точно известно, что аддисонова болезнь связана с гипофункцией коры надпочечников; возможно, что это нарушение имеет место и при артрите, поскольку на таких больных благотворно действует введение гормона коры надпочечников. По этой причине при постановке пробы с мехолилом следует исключать заболевания, связанные с гипофункцией коры надпочечников. Если учесть, что введение адренокортикоидов вызывает у больных артритом психические расстройства [704], то есть все основания предположить, что эти гормоны оказывают сильное влияние на реактивность вегетативных центров. По-видимому, при пониженной их секреции симпатическая реактивность гипоталамуса снижается, а при избыточной — повышается. Проба с мехолилом, проведенная до и после лечения, должна выявить переход из группы III в группу II или I, что может предшествовать появлению психических расстройств и служить ценным объективным показателем изменения состояния мозга. Интересно было бы выяснить, имеются ли при этом другие признаки симпатической реактивности (холодовая проба, кож-но-гальванический рефлекс) и изменяется ли одновременно возбудимость парасимпатической системы (проба с норадреналином).
Эти клинические наблюдения следует дополнить экспериментами на животных с вживленными в гипоталамус электродами. У этих животных рекомендуется исследовать порог реакции ярости перед адреналэктомией, после этой операции, когда животному вводится 3%-ный NaCl, и после введения соответствующего и избыточного количества адренокортикоидов.
Ниже приводятся некоторые дополнительные данные, касающиеся физиологии гипоталамуса.
Барорецепторные рефлексы и гипоталамус
Критерием реактивности и равновесия систем гипоталамуса в значительной части наших исследований служило действие барорецепторных рефлексов. Ослабление этих рефлексов достигалось понижением кровя-
5ti>
ГЛАВА XXVI
ного давления при помощи ацстилхолина и других гипотензивных средств, а для их усиления применяли нор-адреналин и адреналин. Различный характер действия этих препаратов до и после денервации сино-аорталь-ной зоны свидетельствует о первостепенной роли сино-аортальных барорецепторных рефлексов в механизме действия этих препаратов. Аналогичный характер действия на гипоталамус различных гипотензивных препаратов также говорит не о специфическом действии фармакологических веществ, а о роли буферных рефлексов.
Желательно, однако, повторить эту работу в условиях строгого физиологического эксперимента. Изменение давления в области синуса у ненаркотизированной собаки можно вызвать путем выведения наружу сонной артерии, как это делал Кох [547]. Он утверждал, что повышение давления в области синуса вызывает сон. Хотя эту работу не удалось точно воспроизвести (см. [458]), тем не менее было установлено, что повышение давления в области синуса ведет к появлению медленных (характерных для сна) потенциалов в ЭЭГ, а падение давления в области синуса вызывает противоположный эффект. Было показано, что это действие не зависит от изменений общего кровяного давления [76]. Аналогичные изменения потенциалов коры и гипоталамуса возникали при изменении давления в синусе под влиянием гипо- и гипертензивных препаратов [674].
Рекомендуется исследовать влияние изменений давления в синусе на реактивность гипоталамуса. Предварительная операция состоит в имплантации электродов в передний и задний отделы гипоталамуса и выведении наружу сонной артерии. В заключительном эксперименте надо исследовать возбудимость переднего и заднего отделов гипоталамуса при повышении давления в сонной артерии. Следует ожидать, что в этих условиях тонус парасимпатической системы повысится, а тонус симпатической понизится. Это должно проявиться в сужении зрачка, смыкании век, расслаблении третьего века и потере мышечного тонуса. Порог электрического раздражения заднего отдела гипоталамуса должен быть повышен.
При регистрации потенциалов коры и гипоталамуса, при повышении давления в синусе можно ожидать
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ 589
появления медленных потенциалов в этих структурах. Пропуская ток через решетку; на которой стоит животное, можно определить болевой порог и влияние боли на потенциалы гипоталамуса и коры [59]. Следует ожидать, что с повышением давления в синусе болевой порог повысится, а эффективность надпороговых раздражений уменьшится.
Вегетативные индикаторы и гипоталамус
Многие исследователи, изучавшие реактивность вегетативной нервной системы в клинических условиях или стремившиеся исследовать влияние эмоций на центральную нервную систему, применяли для определения реактивности парасимпатической системы такие вещества, как пилокарпин и атропин. Поскольку при применении малых доз эти вещества действуют главным образом на периферические рецепторы, они не могут иметь значения для изучения реактивности вегетативных центров. При применении больших доз атропин вызывает центральное (психомоторное) возбуждение.
Рефлексы, вызывающие изменения функций вегетативной нервной системы, в различной степени связаны с функциями спинного мозга и супраспинальных структур. Их можно использовать в качестве показателя эмоционального состояния, если доказано, что данный рефлекс отражает изменения в состоянии систем гипоталамуса. Исходя из этого, мы рекомендуем пробы с мехо-лилом и норадреналином в качестве индикаторов реактивности гипоталамуса.
Было бы полезно увеличить число индикаторов реактивности гипоталамуса, пригодных для использования при обследовании здоровых и больных людей. Примером таких индикаторов, позволяющих получить ценные данные о состоянии парасимпатической и симпатической систем у человека, могут служить циркуляторные реакции, использованные Везлером [941]. Мы указывали выше, что по этим индикаторам можно дифференцировать ваготонию и симпатотонию (гл. XV). Интересно выяснить, в какой степени минутный объем сердца, эластическое сопротивление артерий, артерио-венозная раз-
ГЛАВА XXVI
ница по кислороду и другие гемодинамические параметры зависят от состояния гипоталамуса. Его активность можно изменить не только рефлекторным путем, но и непосредственно, путем повреждения или введения фармакологических препаратов (см. [310]).
Гипогликемия и гипоталамус
Интересно изучить влияние гипогликемии на порог реакции ярости у кошек с электродами, вживленными в задний отдел гипоталамуса и миндалевидное ядро. Можно предположить, что незначительная гипогликемия приведет к снижению порога реакции ярости при раздражении заднего отдела гипоталамуса. Если это так, то было бы интересно выяснить характер влияния гипогликемии на порог реакции ярости, вызванной раздражением миндалевидного ядра, и, кроме того, установить, снимается ли подобный эффект (понижение порога под влиянием гипогликемии) при незначительных повреждениях заднего отдела гипоталамуса (см. [244, 296, 536, 537, 541]).
Многократные сеансы электрошока и гипоталамус
Хотя влияние электрошока на условные рефлексы, а также на симпатические рефлексы было предметом многочисленных исследований, по-видимому, соответствующие опыты на животных с вживленными электродами не ставились. Интересно было бы изучить влияние возрастающего числа сеансов электрошока на реактивность гипоталамуса, определяемую через различные интервалы после шока. Это помогло бы установить, сопровождается ли повышенная симпатическая реактивность, обнаруженная рядом авторов [360], понижением порога гипоталамической реакции ярости. Одновременно можно было бы исследовать возможность воспроизведения в эксперименте изменений, описанных в ЭЭГ человека [767], и сопоставить их с изменениями реактивности гипоталамуса.
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ
Экспериментальные неврозы
Анализируя физиологические основы экспериментальных неврозов, мы подчеркивали роль гипоталамуса и равновесия его систем. На основе различных данных, имеющихся в специальной литературе, мы высказали предположение, что возбудительную форму экспериментального невроза легче вызвать у животных с повышенной реактивностью симпатического отдела гипоталамуса, а тормозную форму — у животных с преобладанием парасимпатического отдела. Это предположение можно подвергнуть прямой проверке. Большую группу животных— кошек или крыс — делят на три подгруппы: 1) контрольную (неоперированные или с мнимым оперативным вмешательством); 2) подгруппу животных с незначительными повреждениями переднего отдела или (у кошек) вентро-медиалыюго ядра гипоталамуса; 3) подгруппу животных с незначительными повреждениями заднего отдела гипоталамуса.
По всей вероятности, возбудительная форма невроза будет преобладать во 2-й, а тормозная форма—в 3-й подгруппах; 1-я подгруппа займет промежуточное положение.
Можно использовать различные методы воспроизведения невроза. Предневротическое поведение, вызванное незначительной конфликтной ситуацией (легкий воздушный удар во время кормления [510]), может также послужить источником ценной информации, поскольку появление рвоты в этих условиях предполагает сдвиг вегетативного равновесия в сторону преобладания парасимпатической нервной системы (см. гл X). Этот сдвиг равновесия можно использовать для дифференцирования подгрупп 1—3.
Было бы интересно определить относительную эффективность положительных (подкрепляемых) условных раздражителей при выработке условных рефлексов и скорость угашения реакции для этих трех подгрупп; во 2-й подгруппе можно ожидать более быстрого образования условного рефлекса, в 3-й — более быстрого угашения.
Очевидно, это исследование можно распространить и на изучение повреждений лимбического мозга, которые,
ГЛАВА XXVI
как известно, изменяют эмоциональную реактивность и образование условных рефлексов (см. 1Л. XXV).
ИЗУЧЕНИЕ ЭКСПЕРИМЕНТАЛЬНЫХ СУДОРОГ
Подкорковый судорожный разряд при асфиксии
Гельгорн, Баллин и Риггл [330] показали, что после внутривенной инъекции стрихнина длительная асфиксия вызывает вслед за исчезновением корковых потенциалов двухсторонние судорожные разряды, которые охватывают подкорковые структуры, а также кору головного мозга и мозжечок. Важная роль подкорки очевидна из того факта, что местная инъекция стрихнина в задний отдел гипоталамуса во время асфиксии вызывает такие же разряды. Несколько экспериментов, проведенных после стрихнинизации ядер таламуса и хвостатого ядра (в одном случае), также дали положительные результаты. Учитывая значение гипоталамуса для эмоций, а также возникновение общих судорог при эмоциональном возбуждении или при раздражении заднего отдела гипоталамуса, рекомендуется определить значение гипоталамуса для этого генерализованного подкоркового судорожного разряда.
После получения описанного Гельгорном типичного судорожного разряда, вызванного инъекцией стрихнина, интересно повторить опыт с асфиксией после двухстороннего и одностороннего повреждения ретикулярной формации. Целесообразно также поставить опыт с асфиксией после введения стрихнина в ретикулярную формацию и затем повторить его после повреждения заднего отдела гипоталамуса. Мы полагаем, что асфиксия вызовет судорожный разряд после введения стрихнина в ретикулярную формацию. Подобный эксперимент еще никогда не ставился. Если его результаты окажутся отрицательными, то можно изучить роль ретикулярной формации в возникновении судорожного разряда под влиянием асфиксии на фойе внутривенного введения стрихнина и установить, препятствует ли повреждение ретикулярной формации возникновению такого разряда. Сочетая эти пробы с другими экспери-
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ
ментальными повреждениями, можно определить относительную роль гипоталамуса и ретикулярной формации в судорожном разряде при асфиксии. Более того, можно установить значение различных подкорковых структур для распространения судорожной активности. В исследовании Гельгорна отмечалось, что эзерин способствует облегчению подобного судорожного разряда. Поскольку было установлено, что в диффузной активации коры со стороны ретикулярной формации холинэргическая и адренэргическая передача играет известную роль, интересно было бы исследовать, не происходит ли облегчение судорог под влиянием адреналина. Редкие случаи возникновения диффузного судорожного разряда не во время самой асфиксии, а после восстановления доступа воздуха можно считать показателем роли адреномедуллярного гормона, секреция которого в этот момент сильно возрастает [320]. Для подтверждения этого предположения рекомендуется установить: 1) влияет ли на подкорковый судорожный разряд временная перевязка вен надпочечников; 2) облегчается ли этот судорожный разряд при внутривенном введении адреналина или норадреналина.
Роль тормозной системы при экспериментальных судорогах
В ряде работ [891] было показано, что раздражение хвостатого ядра или интраламииарных ядер таламу-са с низкой частотой (5 в 1 сек) вызывает синхронизированные потенциалы коры, а раздражение этих же структур, а также заднего отдела гипоталамуса или ретикулярной формации с высокой частотой (100 в 1 сек) вызывает десинхронизацию в ЭЭГ. Далее было показано [321], что слабое раздражение заднего отдела гипоталамуса и ретикулярной формации вызывает подавление разрядов, вызванных нанесением стрихнина на кору, а применение сильных раздражителей усиливает эти судорожные явления (при частоте в обоих случаях 100 в 1 сек). Поскольку считается, что судорожные разряды являются результатом гиперсинхронизации, было бы интересно исследовать влияние тормоз-
594 глава xxvi
ной системы на судорожные разряды. Можно рекомендовать следующие эксперименты:
I. Исследование судорожных разрядов, вызванных нанесением стрихнина на кору. Условия эксперимента подробно описаны в работе Гельгорна [321].
1. При раздражении гипоталамуса или ретикулярной формации с частотой 100 в 1 сек в течение 5 сек установить порог (напряжение) для:
а) подавления или ослабления;
б) возрастания интенсивности и частоты судорожных разрядов.
2. Повторить опыт на фоне раздражения с низкой частотой хвостатого ядра или интраламинарных ядер, которое вызывает появление веретен в обычной ЗЭГ (без применения стрихнина). Определить, усиливаются или ослабляются эффекты, полученные в опыте 1, а и б.
II. Исследование судорожных корковых разрядов, вызванных электрическим раздражением.
Использовать условия эксперимента Ганглоффа и Монье [280] на ненаркотизированном кролике, у которого судорожные разряды вызываются электрическим раздражением коры, и определить, влияет ли на эти судорожные разряды раздражение хвостатого ядра с низкой частотой, вызывающее веретена в контрольных условиях.
ДРУГИЕ ПРОБЛЕМЫ НЕЙРОФИЗИОЛОГИИ
Влияние хвостатого ядра на сенсорную кору
Хотя известно, что раздражение хвостатого ядр оказывает тормозное влияние на вызванные раздра) нием коры движения [640] и судороги [290а] и привод* к возникновению веретен в ЭЭГ [891], систематичес* исследование влияния хвостатого ядра на сенсорпх кору не производилось. Рекомендуется провести и дующие исследования:
1. Изучить влияние низкочастотного раздражения хвостатого ядра на стрихнинные разряды, вызванные локальным нанесением стрихнина на сенсорные проекционные зоны коры.
I
ДАЛЬНЕЙШИЕ НАПРАВЛЕНИЯ ИССЛЕДОВАНИЯ
2. Изучить влияние раздражения хвостатого ядра на специфические потенциалы, вызванные в соответствующей проекционной зояе зрительными или слуховыми раздражениями; подобный эксперимент можно провести также после предварительного локального нанесения стрихнина; следует ожидать, что в этих условиях судорожная активность ограничится соответствующей проекционной зоной.
3. Поскольку показано [347, 349], что раздражение гипоталамуса или болевое раздражение облегчает реакцию сенсорной проекционной зоны на слуховые и зрительные раздражения, интересно проверить, затормаживается ли этот эффект во время низкочастотного раздражения хвостатого ядра.
4. Интересно исследовать влияние высокочастотного (100 в 1 сек) раздражения хвостатого ядра в условиях, перечисленных в пп. 1—3. Известно, что этот вид раздражения вызывает диффузное возбуждение коры головного мозга, такое же, как при раздражении заднего отдела гипоталамуса или ретикулярной формации. Можно ожидать, что этот вид раздражения приведет к усилению сенсорных реакций.
О функциях заднего отдела гипоталамуса а ретикулярной формации
1. Влияние на стрихнинные разряды. Указывалось [329], что при раздражении ретикулярной формации слабым током стрихнинные разряды коры подавляются, а… 596 ГЛАВА XXVI 2. В опубликованных ранее наших работах [347, 349] отмечалось, что взаимодействие специфических афферентных…– Конец работы –
Используемые теги: Физиология, рефлексов0.05
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Физиология рефлексов
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.179 сек.
Новости и инфо для студентов