КРАТКАЯ ТЕОРИЯ
Живые ткани являются источником биопотенциалов. Регистрация биопотенциалов тканей и органов с диагностической целью называется электрографией. Более конкретные диагностические методы: электрокардиография (ЭКГ) – регистрация биопотенциалов, возникающих в сердечной мышце при ее возбуждении; электроэнцефалография (ЭЭГ) – метод регистрации биоэлектрической активности головного мозга; электромиография – метод регистрации биоэлектрической активности мышц и др.
В большинстве случаев биопотенциалы регистрируются не непосредственно на органе (сердце, головной мозг и др.), а на «соседних» тканях, в которых электрическое поле этим органом создается. Физический подход к электрографии заключается в создании (выборе) модели электрического генератора, которая соответствует картине регистрируемых потенциалов.
Физическую основу метода электрокардиографии представляет теория отведений Эйнтховена. Согласно теории Эйнтховена сердце есть токовый диполь с дипольным моментом Рс, который изменяет свое положение и точку приложения за время сердечного цикла. Этот диполь находится в центре равностороннего треугольника в электропроводящей среде и создает вокруг себя электрическое поле. Дипольный момент Рс пропорционален потенциалу электрического поля, созданного диполем. Рс~j или Рс~Dj. Эйнтховен предложил «снимать» разность биопотенциалов сердца между вершинами равностороннего треугольника. В этом случае мы регистрируем величины, пропорциональные проекциям сердечного вектора Рс на стороны треугольника: Dj1 ~Рс1;. Dj2 ~Рс2; Dj3 ~Рс3.
Разность биопотенциалов, регистрируемая между двумя точками тела, называют отведением. Формируются три стандартных отведения Эйнтховена, причем, используя понятие эквипотенциальных поверхностей (поверхностей равных потенциалов), подключать электроды можно к конечностям, считая их точками, потенциалы в которых равны потенциалам в вершинах равностороннего треугольника.
По существу, сердце представляет собой электрический мультипольный генератор – это некоторая пространственная совокупность источников электрических полей (совокупность истоков и стоков токов). Поэтому в методе электрографии следует использовать принцип суперпозиции полей (наложения поле). Если поле создается несколькими источниками, то напряженность поля равна векторной сумме напряженностей полей каждого источника 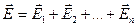 , а потенциал поля равен алгебраической сумме потенциалов источников
, а потенциал поля равен алгебраической сумме потенциалов источников 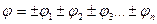 (считаем, что отрицательный заряд создает поле отрицательного потенциала, а положительный – положительного).
(считаем, что отрицательный заряд создает поле отрицательного потенциала, а положительный – положительного).
Элементарным источником поля (ЭДС) можно считать клетку, на поверхности мембраны которой формируется потенциал возбуждения. Процесс формирования разности потенциалов на поверхности одиночного мышечного волокна и генез электрограммы можно проследить на примере формирования зубцов электрокардиограммы.
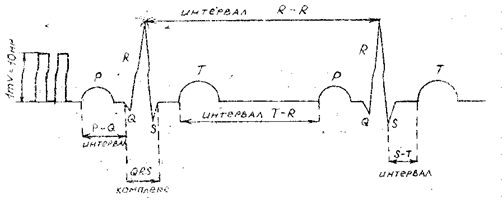
Электрокардиограмма - это график зависимости колебаний разности потенциалов электрического поля сердца при возбуждении сердечной мышцы во времени. ЭКГ имеет вид повторяющихся, согласно сердечному циклу, комплексов, зубцов и интервалов. Отдельные части кривой, обозначаемые буквами P, Q, R, S, T, связаны с возбуждением сердца и (при отсутствии нарушения ритма) повторяются во время каждого цикла, сохраняя неизменной форму при сохранении условий измерения. Физический механизм возникновения отдельных зубцов иллюстрирует следующая схема (риc. 1).
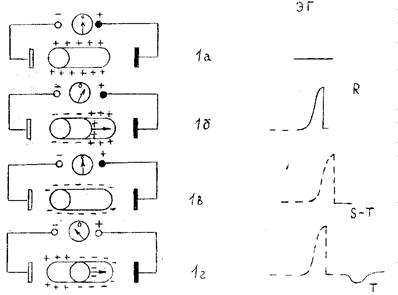
Рис. 1. Процесс формирования разности потенциалов на поверхности одиночного мышечного волокна и генез электрограммы (ЭГ) волокна (стрелки обозначают направление движения волны деполяризации и реполяризации).
В состоянии покоя вся наружная поверхность клеточной мембраны заряжена положительно. Между любыми двумя точками этой поверхности разность потенциалов равна 0. На ЭГ одиночного мышечного волокна, зарегистрированной с помощью двух электродов, расположенных на поверхности клетки, записывается горизонтальная нулевая (изоэлектрическая) линия (рис. 1а). При возбуждении миокардиального волокна (рис. 1б) наружная поверхность деполяризованного участка заряжается отрицательно по отношению к поверхности участка, находящегося еще в состоянии покоя (поляризации); между ними появляется разность потенциалов, которая и может быть зарегистрирована на ЭГ в виде положительного отклонения, направленного вверх от изолинии - зубца R на ЭКГ. Когда все волокно окажется в состоянии возбуждения (рис. 1в) и вся его поверхность будет заряжена отрицательно, разность потенциалов между электродами снова окажется равной нулю, и на ЭГ будет записываться изолиния. На ЭКГ это сегмент S-T. Процесс быстрой конечной реполяризации одиночного мышечного волокна начинается в том же участке, что и волна деполяризации (рис. 1г). При этом поверхность реполяризованного участка заряжается положительно, и между двумя электродами, расположенными на поверхности волокна, вновь возникает разность потенциалов, которая на ЭГ проявляется новым отклонением от изолинии - зубцом Т ЭГ. Поскольку к электроду, соединенному с "+" электрокардиографа, теперь обращена поверхность с отрицательным, а не с положительным зарядом, как при распространении волны деполяризации, на ЭГ будет регистрироваться не положительный, а отрицательный зубец Т. Кроме того, в связи с тем, что скорость распространения процесса реполяризации значительно меньше скорости перемещения фронта деполяризации, продолжительность зубца Т на ЭГ больше таковой зубца R, а амплитуда меньше.
Следует отметить, что на форму зубцов ЭГ влияет не только электрическая активность самого мышечного волокна, но и место расположения положительного и отрицательного электродов отведения, с помощью которого регистрируется ЭГ.
На рис. 2 изображена форма нормальной ЭКГ (II стандартное отведение) и прямоугольный импульс калибровочного сигнала амплитудой 1 мВ.