рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Экология
- /
- Основы общей экологии
Реферат Курсовая Конспект
Основы общей экологии
Основы общей экологии - раздел Экология, Н.а.воронков Основы Общей Экологии (Общеобразовательный Кур...
Н.А.Воронков
Основы общей экологии
(Общеобразовательный курс)
Издание четвертое доработанное и дополненное
Рекомендовано Министерством образования Российской Федерации в качестве учебника для студентов высших учебных заведений
Допущено Департаментом общего среднего образования Министерства образования Российской Федерации в качестве пособия для учителей
«Агар» «Рандеву-АМ»
Москва 1999
ББК20Я73
Воронков Н.А.
Основы общей экологии: Учебник для студентов высших учебных заведений. Пособие для учителей. - М.: Агар, 1999. - 96 с.
В соответствии с вновь вводимыми стандартами высшего образования, экологию должны изучать студенты всех специальностей в блоке общекультурных дисциплин. Настоящий учебник ставит целью оказать помощь в решении данной весьма актуальной задачи экологического всеобуча. В нем в доступной форме, при максимально возможном сохранении научного уровня, рассматриваются основные вопросы современной экологии. Учебник является результатом обобщения опыта длительной научной работы автора в области экологии и преподавания данной дисциплины студентам всех специальностей (факультетов) Московского государственного открытого педагогического университета.
Предлагаемая читателю первая часть учебника посвящена наиболее значимым вопросам общей, или классической экологии, рассматривающей основные закономерности функционирования природных систем различного ранга (от биосферы до элементарных экосистем и популяций), их устойчивость, энергетику, продуктивность, роль в сохранении жизни и другие свойства.
Во второй части (издается в 1999 году) освещаются проблемы социальной, прикладной и других разделов экологии, связанных с деятельностью человека.
Кроме использования студентами и учителями, будет полезен слушателям университетов повышения квалификации, учащимся старших классов школ, а также всем лицам, интересующимся вопросами экологии и экологического образования.
Автор: Воронков Н. А.
Рецензенты:
Академик Российской академии образования, д.б.н. И.Д.Зверев.
Проректор Московского института повышения квалификации работников образования, действительный член Международной академии наук, профессор, доктор физико-математических наук Ю.Л.Хотунцев.
Кафедра зоологии и экологии МПГУ, доктор биологических наук, профессор Н.М.Чернова.
ISBN 5-89218-098-0 ISBN 5-93290-008-3
© Издательство «Агар», 1999
© Н.А.Воронков
СЛОВО К ЧИТАТЕЛЮ! (вместо предисловия)
Автор учебника, который Вам предлагается, стремится познакомить Вас с началами экологии как науки, пробудить интерес к этой весьма актуальной и интересной отрасли знаний.
В предлагаемой первой части учебника ставится цель показать, по каким закономерностям природа формировалась миллиарды лет и существовала без участия человека; утвердить Вас в мысли, что среда, в которой человек обитает, создана прежде всего живыми организмами и продуктами их жизнедеятельности, и что сбережение этой среды возможно только при непременном условии сохранения всего разнообразия жизни. Из этого следует, что любые попытки человека и человечества решить свои проблемы существования и выживания в одиночку (без сохранения всего разнообразия жизни), даже самыми современными техническими средствами, однозначно не состоятельны.
Современная среда обитания и свойственная ей жизнь на Земле порождены живыми организмами многих геологических эпох, они же являются и условием продолжения жизни. Неверен даже самый гуманный тезис, провозглашенный человеком: все окружающие существа - братья наши меньшие. Человек в существующем мире - самый младший брат, хотя и щедро наделенный старшими братьями разумом и невиданной силой. Свой разум и силу человек должен использовать не для того, чтобы больше взять у старших братьев, а также из их кладовых, наполнявшихся миллиарды лет, а для осознания того факта, что старшие братья существовали миллионы лет без младшего и могут продолжать существование без него. Младший же без старших не проживет и нескольких дней. Поэтому силу и ум, дарованные ему старшими братьями, он ни в коем случае не должен использовать во вред им, часто немощным и беззащитным перед мощью человека, а в помощь, хотя бы для того, чтобы вернуть долги, залечить раны, искупить свою вину и не повторять ошибок.
Тезис «меньших братьев» неверен и потому, что предполагает учение их уму-разуму. На самом же деле человек должен, наконец, осознать, что ему, и прежде всего ему, есть чему учиться у братьев старших, если не у каждого в отдельности, то уж у коллективного разума - обязательно!
Вторая часть пособия («Социальная и прикладная экология») посвящена рассмотрению тех проблем, которые человек создал мгновениями своей жизни в том мире, который многие миллионы и миллиарды лет жил по своим нормам и правилам; каковы масштабы его деятельности и к чему надо стремиться, чтобы стать равным среди равных и не разрушить наш общий дом. К сожалению, пока далеко не все ясно, что конкретно надо делать для исправления сложившейся и усугубляющейся с каждым днем весьма тревожной ситуации. Однако известно, что поиск выхода возможен только совместными усилиями всех и каждого на основе познания себя и мира, в котором мы живем, и прежде всего глубоких экологических знаний. Незнание и равнодушие - дорога в пропасть!
Если этот учебник вызовет у Вас интерес к науке и учебной дисциплине, имя которой «Экология», а знакомство с ее проблемами хоть в какой-то мере заставит Вас задуматься о своем месте и роли в окружающем мире, автор будет глубоко удовлетворен и с благодарностью примет и учтет в дальнейшей работе все Ваши замечания и пожелания, которые можно направлять по адресу: 109004 Москва, ул. Верхняя Радищевская, 18, МГОПУ, биолого-химический факультет. По этому же адресу можно приобрести книгу.
ВВЕДЕНИЕ
Существует образное выражение, что мы живем в эпоху трех «Э»: экономика, энергетика, экология. При этом экология как наука и образ мышления привлекает все более и более пристальное внимание человечества.
Экологию рассматривают как науку и учебную дисциплину, которая призвана изучать взаимоотношения организмов и среды во всем их разнообразии. При этом под средой понимается не только мир неживой природы, а и воздействие одних организмов или их сообществ на другие организмы и сообщества.
Термин «экология» был введен в употребление немецким естествоиспытателем Э. Геккелем в 1866 году и в дословном переводе с греческого обозначает науку о доме (ойкос - дом, жилище; логос - учение).
По этой причине экологию иногда связывают только с учением о среде обитания (доме) или окружающей среде. Последнее в основе правильно с той, однако, существенной поправкой, что среду нельзя рассматривать в отрыве от организмов, как и организмы вне их среды обитания. Это составные части единого функционального целого, что и подчеркивается приведенным выше определением экологии как науки о взаимоотношениях организмов и среды.
Такую двустороннюю связь важно подчеркнуть в связи с тем, что это основополагающее положение часто не доучитывается: экологию сводят только к влиянию среды на организмы. Ошибочность таких положений очевидна, поскольку, как будет показано ниже, именно организмы сформировали современную среду. Им же принадлежит первостепенная роль в нейтрализации тех воздействий на среду, которые происходили и происходят по различным причинам.
Концептуальные основы дисциплины. С момента появления «Экология» развивалась в рамках биологии практически на протяжении целого века - до 60-70-х годов настоящего столетия. Человек в этих системах, как правило, не рассматривался - полагалось, что его взаимоотношения со средой подчиняются не биологическим, а социальным закономерностям и являются объектом общественно-философских наук.
В настоящее время термин «экология» существенно трансформировался. Она стала больше ориентированной на человека в связи с его исключительно масштабным и специфическим влиянием на среду.
Сказанное позволяет дополнить определение «экологии» и назвать задачи, которые она призвана решать в настоящее время. Современную экологию можно рассматривать как науку, занимающуюся изучением взаимоотношений организмов, в том числе и человека, со средой, определением масштабов и допустимых пределов воздействия человеческого общества на среду, возможностей уменьшения этих воздействий или их полной нейтрализации. В стратегическом плане - это наука о выживании человечества и выходе из экологического кризиса, который приобрел (или приобретает) глобальные масштабы - в пределах всей планеты Земля.
Становится все более ясным, что человек очень мало знает о среде, в которой он живет, особенно о механизмах, которые формируют и сохраняют среду. Раскрытие этих механизмов (закономерностей) -одна из важнейших задач современной экологии и экологического образования. Ясно, что она может решаться лишь при условии изучения не только «Дома», но и его обитателей, их образа жизни.
Содержание термина «экология», таким образом, приобрело социально-политический, философский аспект. Она стала проникать практически во все отрасли знаний, с ней связывается гуманизация естественных и технических наук, она активно внедряется в гуманитарные области знаний. Экология при этом рассматривается не только как самостоятельная дисциплина, а как мировоззрение, призванное пронизывать все науки, технологические процессы и сферы деятельности людей.
Признано поэтому, что экологическая подготовка должна идти, по крайней мере, по двум направлениям через изучение специальных интегральных курсов и через экологизацию всей научной, производственной и педагогической деятельности.
Решению этих крайне актуальных вопросов и призван помочь предлагаемый курс. Как отмечено в аннотации, основной целью его является изложение тех основ экологии, с которыми должен быть знаком каждый обучающийся вне зависимости от его специальности. Сказанное не исключает, а полагает, что вопросы, ориентированные на конкретные отрасли знаний, должны рассматриваться в специальных экологических курсах. Ясно, что без основательной общеэкологической подготовки экологизация образования, как и деятельности человека, практически невозможна, а если она и проводится - то либо не достигает цели, либо имеет результат, противоположный ожидаемому, так как базируется на случайных, часто фрагментарных положениях, что недопустимо для системной науки, к рангу которой относится «Экология».
Наряду с экологическим образованием существенное внимание уделяется экологическому воспитанию, с которым связывается бережное отношение к природе, культурному наследию, социальным благам. Без серьезного общеэкологического образования решение этой задачи также весьма проблематично.
Между тем, став в своем роде модной, экология не избежала вульгаризации понимания и содержания. В ряде случаев экология становится разменной монетой в достижении определенных политических целей, положения в обществе.
В разряд экологических нередко возводятся вопросы, относящиеся к отраслям производства, видам и результатам деятельности человека, просто если к ним добавляют модное слово «экология». Так появляются несуразные выражения, в том числе и в печати, типа «хорошая и плохая экология», «чистая и грязная экология», «испорченная экология» и др. Это равнозначно присвоению таких же эпитетов математике, физике, истории, педагогике и т. п.
По этому же принципу ранг экологии присваивается многим разделам гуманитарных (философии, социологии, экономики) и естественных наук (биологии, естествознания, географии).
Несмотря на отмеченные неясности и издержки в понимании объема, содержания и использования термина «экология», несомненным остается факт ее крайней актуальности в настоящее время.
Предлагаемая первая часть учебника («Общая экология») является теоретической базой второй («Социальная и прикладная экология»), которая, в свою очередь, является логическим продолжением первой. В таком случае более частные вопросы рассматриваются в ранге отдельных проблем названных выше двух крупных разделов (частей). Так, например, во второй части пособия рассматриваются экологические проблемы (но не «экологии») промышленности, земельных ресурсов, продовольствия, водных ресурсов, городов, энергетики, народонаселения, состояния среды и здоровья и др.
В обобщенном виде «Общая экология» изучает наиболее общие закономерности взаимоотношений организмов и их сообществ со средой в естественных условиях.
«Социальная экология» рассматривает взаимоотношения в системе «общество - природа», специфическую роль человека в системах различного ранга, отличие этой роли от других живых существ, пути оптимизации взаимоотношений человека со средой, теоретические основы рационального природопользования.
«Прикладная экология» призвана решать конкретные вопросы природопользования, определять допустимые нагрузки на среду, разрабатывать методы управления природными системами (экосистемами) и способы «экологизации» различных видов деятельности человека.
С точки зрения основного содержания предмета «Общая экология» есть не что иное, как экология природных систем и учение о природной среде, а «Социальная и прикладная экология» - экология измененных человеком природных систем и среды, или экология природно-антропогенных систем и учение о природно-антропогенной (иногда техногенной) среде.
Краткий исторический очерк. Общеэкологические подходы к рассмотрению и оценке природных явлений имеют длительную историю. По сути своей в значительной мере экологичными были труды первых ученых-естествоиспытателей, искавших зависимости между свойствами живых существ и условиями обитания: Аристотель (384-322 г. до н. э.), его ученик-ботаник Теофраст (371-280 г. до н. э.). Много ценных материалов поставили исследователи-натуралисты, занимавшиеся описанием и систематизацией растений и животных.
Особо следует выделить труд Ч. Дарвина «Происхождение видов» (1859), в котором большое внимание уделяется приспособлениям (адаптациям) и взаимоотношениям организмов. Э. Геккель, вводя термин «экология», отмечал, что одной из задач данной науки является исследование всех тех взаимоотношений организмов, которые Ч. Дарвин условно обозначил как борьбу за существование.
Из отечественных ученых наиболее существенный вклад в развитие отдельных разделов общей экологии и прежде всего системный взгляд на природные явления внесли исследования почвоведа-географа В. В. Докучаева (1846-1903) и его школы (Г. Ф. Морозов, Г. Н. Высоцкий, В. И. Вернадский и др.). В. В. Докучаев показал тесную взаимосвязь живых организмов и неживой природы на примере почвообразования и выделения природных зон. Г. Ф. Морозов (1867-1920) раскрыл всесторонние связи в лесных сообществах и рассмотрел их как единые системы, включающие весь свойственный им комплекс живых организмов и условий обитания, их средообразовательную роль. В этом же направлении, но применительно к решению конкретных вопросов степного лесоразделения, проводил свои исследования ботаник, почвовед, географ Г.Н.Высоцкий (1865-1940).
В. И. Вернадский (1863-1945) системный подход применил к раскрытию основополагающих геологических явлений и их эволюции, показал определяющую роль живых организмов и продуктов их жизнедеятельности в этих явлениях, стал автором учения о биосфере и закономерностях ее существования, устойчивости и развития.
Оригинальны и интересны исследования В. Н. Сукачева (1880-1967), посвятившего многие годы комплексному изучению лесных систем (сообществ), результатом чего явилось всестороннее рассмотрение единства и взаимообусловленности природных явлений, живой и неживой материи. Им в 1942 г. введен в науку термин «биогеоценоз», раскрыто его содержание.
Несколько раньше (в 1935 г.) подобные идеи сформулировал английский ботаник-эколог А. Тенсли и ввел в науку термин «экосистема», дал его определение. В настоящее время эти понятия являются определяющими для экологии как науки.
В числе других ученых, которые либо развивали, либо обогащали различные аспекты общей экологии как науки (многие из них являются авторами учебников и учебных пособий), следует назвать Д. Н. Кашкарова, Ч. Элтона, Н. П. Наумова, С. С. Шварца, М. С. Гилярова - труды по вопросам экологии животных; А. П. Шенникова, Ф. Клементса, В. Лархера и др. - комплекс работ по экологии растений; Г. Одума, Ю. Одума, Р. Уиттекера, Р. Риклефса, М. Бигона и др., Р. Дажо, Н. М. Чернову, А. М. Былову, В. А. Радкевича, И. Н. Пономареву, И. А. Шилова и др. - учебники и учебные пособия по проблемам общей экологии.
Одно из первых высказываний, относящихся к сфере социальной экологии, принадлежит французскому естествоиспытателю-эволюционисту Жану-Батисту Ламарку (1744-1829). Он, наряду с раскрытием ряда закономерностей влияния среды на организмы, впервые обратил серьезное внимание на специфическую роль человека и ее возможные катастрофические последствия. Он писал: «Можно, пожалуй, сказать, что назначение человека как бы заключается в том, чтобы уничтожить свой род, предварительно сделав земной шар непригодным для обитания». Это высказывание перекликается с «Пророчествами» Леонардо да Винчи (1452-1519), предрекавшего появление существ, результаты деятельности которых «... ничего не оставят ни на земле, ни под водой, что не было бы преследуемо и не подвергалось искоренению...».
Различные аспекты экологии и смежных с ней дисциплин содержатся в трудах и учебниках М. И. Будыко, Н. Н. Моисеева, Н.Ф. Реймерса, А. В. Яблокова, Б. Г. Розанова, Б. Коммонера, а также в переведенных в последнее время на русский язык обстоятельных сводках по вопросам различных проблем экологии Б. Небела, Т. Миллера, П. Ревелля, Ч. Ревелля, Л. Р. Брауна и других авторов. Следует также обратить внимание на оригинальный труд «Проблемы экологии России», авторами которого являются К. С. Лосев, В. Г. Горшков, К. Я. Кондратьев и другие ученые.
В целом основная задача курса сводится к формированию общих основ системного взгляда на природные и техногенные процессы как базы для оптимизации деятельности и поведения человека в окружающем мире с целью поиска путей относительно стабильного, а в дальнейшем и устойчивого развития общества, к чему призвала Конференция ООН по окружающей среде и развитию, состоявшаяся в Рио-де-Жанейро в 1992 году.
На первый взгляд, казалось бы, возможно при знакомстве с экологией как дисциплиной ограничиться ее прикладными аспектами и прежде всего мероприятиями по оздоровлению среды, которые сводятся в конечном счете к определенной системе технологических требований, административных запретов и санкций. Однако такой подход недостаточен и односторонен, поскольку не позволяет видеть глубинные причины сложившейся экологической ситуации и тем более обоснованно прогнозировать возможные и часто труд-нопредсказуемые последствия планируемых или осуществляемых действий, в том числе и с самыми благими намерениями. Поэтому крайне важно рассмотреть основные положения общей экологии, которая, как отмечалось, является теоретической основой для решения проблем рационального природопользования и охраны природы, базовой для социальной и прикладной экологии, а также для других, более частных экологических дисциплин.
I. ОСНОВНЫЕ ПОНЯТИЯ (ТЕРМИНЫ) ЭКОЛОГИИ. ЕЕ СИСТЕМНОСТЬ
I.1. Основные понятия
Основным понятием и основной таксономической единицей в экологии является «экосистема». Этот термин, как упоминалось выше, введен в употребление А. Тенсли в 1935 г., т. е. более полувека спустя после выделения экологии как отрасли научных знаний (1866).
Под экосистемой понимается любая система, состоящая из живых существ и среды их обитания, объединенных в единое функциональное целое. Основные свойства экосистем -способность осуществлять круговорот веществ, противостоять внешним воздействиям, производить биологическую продукцию. Выделяют обычно экосистемы различного ранга: от микроэкосистем (небольшой водоем, труп животного с населяющими его организмами или ствол дерева в стадии разложения, аквариум и даже лужица или капля воды, пока они существуют и в них присутствуют живые организмы, способные осуществлять круговорот веществ); мезоэкосистемы (лес, пруд, река и т.п.); макроэкосистемы (океан, континент, природная зона и т. п.) и глобальная экосистема - биосфера в целом.
Таким образом, более крупные экосистемы включают в себя экосистемы меньшего ранга. Образное (шутливое) определение экосистемы дал географ и писатель Г. К. Ефремов: это любое природное образование - «от кочки до оболочки» (географической).
Близкий по содержанию смысл вкладывается в термин «биогеоценоз», введенный в литературу академиком В. Н. Сукачевым несколько позднее, чем «экосистема» - в 1942 г.
Небольшие различия, которые свойственны этим терминам будут рассмотрены в разд. IV. 1 (часть I).
Экосистемы (биогеоценозы) обычно включают два блока. Первый из них состоит из взаимосвязанных организмов разных видов и носит название «биоценоз» (термин введен немецким зоологом К. Мебиусом в 1877 г.), второй блок составляет среда обитания, которую в данном случае называют «биотоп» или «экотоп».
Каждый биоценоз состоит из множества видов, но виды входят в него не отдельными особями, а популяциями или их частями. Популяция - это относительно обособленная часть вида (состоит из особей одного вида), занимающая определенное пространство и способная к саморегулированию и поддерживанию оптимальной численности особей. Каждый вид в пределах занимаемой территории (ареала), таким образом, распадается на популяции. Размеры их различны. В таком случае можно сказать, что биоценоз - это сумма взаимосвязанных между собой и с условиями среды популяций разных видов.
В экологии часто пользуются также термином «сообщество». Содержание этого термина неоднозначно. Под ним понимается и совокупность взаимосвязанных организмов разных видов (синоним биоценоза), и аналогичная совокупность только растительных (фитоценоз, растительное сообщество), животных (зооценоз) организмов или микробного населения (микробоценоз).
Системность экологии. Экология как наука рассматривает системы, звенья и члены которых находятся в тесной взаимосвязи и взаимозависимости. Из этого вытекает необходимость учета множества факторов при анализе тех или иных экологических явлений и тем более при планировании любых вмешательств в экосистемы. Такой подход, в свою очередь, невозможен без комплексного метода изучения, оценки и решения тех или иных экологических задач. По этим же причинам очевидна тесная связь экологии с другими науками, сведениями из которых необходимо не только располагать, но и уметь их грамотно использовать. К таким наукам относятся: биология, география, почвоведение, гидрология, химия, физика и другие отрасли знаний. Важно также уметь пользоваться необходимой информацией из различных отраслей хозяйства и свойственных им технологических процессов.
Говоря о системных явлениях, важно познакомиться с видами систем, общими положениями теории систем. Обычно различают три вида систем: 1) изолированные, которые не обмениваются с соседними ни веществом, ни энергией, 2) закрытые, которые обмениваются с соседними энергией, но не веществом (например, космический корабль), и 3) открытые, которые обмениваются с соседними и веществом, и энергией. Практически все природные (экологические) системы относятся к типу открытых.
Существование систем немыслимо без связей. Последние делят на прямые и обратные. Прямой называют такую связь, при которой один элемент (А) действует на другой (В) без ответной реакции. Примером такой связи может быть действие древесного яруса леса на случайно выросшее под его пологом травянистое растение или действие солнца на земные процессы. При обратной связи элемент В отвечает на действие элемента А. Обратные связи бывают положительными и отрицательными. И те и другие играют существенную роль в экологических процессах и явлениях.
Положительная обратная связь ведет к усилению процесса в одном направлении. Пример ее - заболачивание территории, например, после вырубки леса. Снятие лесного полога и уплотнение почвы обычно ведет к накоплению воды на ее поверхности. Это, в свою очередь, дает возможность поселяться здесь растениям-влагонакопителям, например сфагновым мхам, содержание воды в которых в 25-30 раз превышает вес их тела. Процесс начинает действовать в одном направлении: увеличение увлажнения - обеднение кислородом - замедление разложения растительных остатков - накопление торфа - дальнейшее усиление заболачивания.
Отрицательная обратная связь действует таким образом, что в ответ на усиление действия элемента А увеличивается противоположная по направлению сила действия элемента В. Такая связь позволяет сохраняться системе в состоянии устойчивого динамического равновесия. Это наиболее распространенный и важный вид связей в природных системах. На них прежде всего базируется устойчивость и стабильность экосистем. Пример такой связи - взаимоотношение между хищником и его жертвой. Увеличение численности жертвы как кормового ресурса, например полевых мышей для лис, создает условия для размножения и увеличения численности последних. Они, в свою очередь, начинают более интенсивно уничтожать жертву и снижают ее численность. В целом численность хищника и жертвы синхронно колеблется в определенных границах. Второй пример. В истории биосферы имели место явления локального увеличения содержания углекислого газа в атмосфере, например, при извержении вулканов. За этим следовало повышение интенсивности фотосинтеза и связывание углекислоты в органическом веществе, а также более интенсивное поглощение ее океаном. Третий пример. В природе закономерны периодические повышения уровней почвенно-грунтовых вод. За этим следует увеличение их контакта с корневыми системами растений, повышение расходования на испарение растительностью (транспирацию) и возвращение уровней грунтовой воды в исходное состояние.
Одно из отрицательных проявлений деятельности человека в природе связано с нарушением этих связей, что может привести к разрушению экосистем или переходу их в другое состояние. Например, умеренное загрязнение водной среды органическими и биогенными (необходимыми для жизнедеятельности организмов) веществами обычно сопровождается интенсификацией деятельности организмов, потребляющих эти вещества, результатом чего является самоочищение водоемов. Перегрузка же среды загрязняющими веществами на определенном этапе ведет к угнетению или уничтожению организмов-санитаров, переводу установившихся обратных связей в прямые, переходу системы на другой уровень. В результате неизбежным становится прогрессирующее загрязнение, обеднение водной среды кислородом и превращение чистых озерных или текущих вод в системы болотного типа.
Универсальное свойство экосистем - их эмерджентность (англ. эмердженс - возникновение, появление нового), заключающееся в том, что свойства системы как целого не являются простой суммой свойств слагающих ее частей или элементов. Например, одно дерево, как и редкий древостой, не составляет леса, поскольку не создает определенной среды (почвенной, гидрологической, метеорологической и т. д.) и свойственных лесу взаимосвязей различных звеньев, обусловливающих новое качество. Недоучет эмерджент-ности может приводить к крупным просчетам при вмешательстве человека в жизнь экосистем или при конструировании систем для выполнения определенных целей. Например, сельскохозяйственные поля (агроценозы) имеют низкий коэффициент эмерджентности и поэтому характеризуются крайне низкой способностью саморегулирования и устойчивости. В них, вследствие бедности видового состава организмов, крайне незначительны взаимосвязи, велика вероятность интенсивного размножения отдельных нежелательных видов (сорняков, вредителей).
Энергетические процессы в экосистемах подчиняются первому и второму началам термодинамики. В соответствии с ними энергия не возникает и не исчезает, она лишь переходит из одной формы в другую (первое начало термодинамики). При этом часть энергии при любых ее превращениях рассеивается (теряется) в виде тепла (второе начало термодинамики). Мерой необратимого рассеивания энергии является энтропия (греч. эн - внутрь, тропе - превращение). Последнюю можно характеризовать и через степень упорядоченности системы. Так, живые организмы и нормально функционирующие экосистемы характеризуются высокой степенью упорядоченности слагающих их элементов. Они сохраняют (поддерживают) определенный уровень энергии и тем самым противостоят энтропии. Мертвый организм характеризуется максимальной неупорядоченностью элементов (структур), в результате чего приходит в равновесие с окружающей его средой (температура его тела выравнивается с температурой среды, составляющие его химические элементы и соединения включаются в процессы круговорота и становятся частью среды). Это значит, что организм как система приходит в состояние полной неупорядоченности, максимальной энтропии. Показатель, противоположный энтропии, носит название негэнтропии. Чем выше организованность системы (упорядоченность), тем значительнее ее негэнтропия. Опасно любое вмешательство в систему, которое ведет к снижению ее негэнтропии, а следовательно, устойчивости и способности противостоять внешним возмущениям.
Основным свойством нормально функционирующих природных экосистем является способность извлекать негэнтропию из внешней среды (солнечную энергию) и тем самым поддерживать свою высокую упорядоченность.
Деятельность человека, если она превышает определенные пределы, ведет к снижению негэнтропии систем, а следовательно, уменьшает их способность поддерживать себя в устойчивом состоянии вплоть до перехода к полной неупорядоченности (максимальной энтропии) и гибели.
Видный американский эколог Б. Коммонер сделал удачную попытку обобщить системность экологии как науки в виде четырех законов. Эти законы в основе своей не новы, но впервые сформулированы в образной простой форме. Их соблюдение - обязательное условие любой экологически обусловленной деятельности человека в природе.
Первый закон Коммонера отражает по сути своей всеобщую связь процессов и явлений в природе и звучит так: «Все связано со всем». Второй закон базируется на положении сохранения вещества и энергии: «Все должно куда-то деваться». Какой бы ни была высокой труба завода, она не может выбрасывать отходы производства за пределы биосферы. В такой же мере загрязнители, попадающие в реки, в конечном счете оказываются в морях и океанах и с их продуктами возвращаются к человеку в виде своего рода «экологического бумеранга». Третий закон ориентирует на действия, согласующиеся с природными процессами, сотрудничество с природой, или коадаптацию (лат. ко - с, вместе; адаптацио - приспособление), вместо покорения человеком природы, подчинения ее своим целям: «Природа знает лучше». Сущность четвертого закона заключается в ориентации человека на то, что любое его действие в природе не остается бесследным, мнимая выгода часто оборачивается ущербом, а охрана природы и рациональное использование природных ресурсов немыслимы без определенных экономических затрат. Звучит этот закон так: «Ничто не дается даром». Дешевому природопользованию не должно быть места. Если не заплатим за него мы, то в многократном размере это должны будут сделать пришедшие нам на смену поколения.
Другие термины и понятия, а также закономерности (правила, принципы) экологии, важные для решения социальных и прикладных ее задач, будут рассмотрены во второй части работы (см.разд.1.2 и 1.3).
I.2. Структура общей экологии
В «Общей экологии» обычно выделяют несколько взаимосвязанных разделов, которые иногда рассматривают как отдельные дисциплины (табл. 1). Это: учение о факторах среды и закономерностях их действия на организмы (факториальная экология); экология на уровне взаимоотношения отдельных организмов и среды (экология организмов, или аутэкология); экология взаимосвязанных и относительно обособленных групп организмов одних и тех же видов (популяционная, или демографическая, экология), экология взаимосвязанных популяций различных видов между собой (учение о биоценозах). Если биоценозы рассматриваются во взаимосвязи со средой обитания (как единая система), то этот раздел выделяется в учение об экосистемах или биогеоценозах. Основополагающим и высшим рангом экологии является учение о биосфере как наиболее крупной (глобальной) экосистеме.
Таблица 1
Структура «Общей экологии»

В настоящем общеобразовательном курсе мы познакомимся с теми основными положениями «Общей экологии», которые составляют базу для понимания наиболее существенных моментов функционирования разных природных экосистем и биосферы в целом, раскрывают роль живых организмов (живого вещества, по В. И. Вернадскому) в создании, сохранении и стабилизации природной среды; рассматривают механизмы, обусловливающие устойчивость природных систем различного ранга, и другие основополагающие проблемы. На этом фоне существенно увеличивается возможность научно обоснованного решения конкретных вопросов прикладной и других разделов экологии, ориентированных на человека, а также осуществление основного требования экологизации природопользования и других видов деятельности человека: «мыслить глобально, действовать локально».
Вопросы и задания
1. Дайте определение экологии как науки. Назовите автора термина.
2. Как трансформировать содержание и задачи экологии в современный период?
3. Какие вопросы и проблемы рассматривает «Общая экология»? Назовите основные ее разделы.
4. Какие вопросы и проблемы являются предметом изучения «Социальной и прикладной экологии»?
5. Почему экологию относят к системным наукам? Назовите основные виды систем и присущие им связи.
6. Дайте определение понятий (терминов) экологии: «экосистема», «биоценоз», «сообщество», «популяция». К какому виду систем относится «экосистема» и почему? Какие для нее присущи связи? Приведите примеры.
7. Раскройте содержание понятий «эмерджентность», «энтропия», «негэнтропия».
8. Назовите экологические законы Б. Коммонера. Раскройте их содержание.
II. СРЕДА ОБИТАНИЯ. ФАКТОРЫ СРЕДЫ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ. СРЕДЫ ЖИЗНИ
II.1. Среда и факторы среды, их классификация
Под средой обитания обычно понимают природные тела и явления, с которыми организм (организмы) находятся в прямых или косвенных взаимоотношениях. Отдельные элементы среды, на которые организмы реагируют приспособительными реакциями (адап-тациями), носят название факторов.
Наряду с термином «среда обитания» используются также понятия «экологическая среда», «местообитание», «окружающая среда», «окружающая природная среда», «окружающая природа» и др. Четких различий между этими терминами нет, но на некоторых из них следует остановиться. В частности, под популярным в последнее время термином «окружающая среда» понимается, как правило, среда, в той или иной (в большинстве случаев в значительной) мере измененная человеком. К ней близки по смыслу «техногенная среда», «антропогенная среда», «промышленная среда».
Природная среда, окружающая природа - это среда, не измененная человеком или измененная в малой степени. С термином «местообитание» обычно связывается та среда жизни организма или вида, в которой осуществляется весь цикл его развития.
В «Общей экологии» речь обычно идет о природной среде, окружающей природе, местообитаниях; в «Прикладной и социальной экологии» - об окружающей среде. Этот термин часто считают неудачным переводом с английского environment, поскольку отсутствует указание на объект, который окружает среда.
Влияние среды на организмы обычно оценивают через отдельные факторы (лат. делающий, производящий). Под экологическими факторами понимается любой элемент или условие среды, на которые организмы реагируют приспособительными реакциями, или адаптациями. За пределами приспособительных реакций лежат летальные (гибельные для организмов) значения факторов.
Классификация факторов:
Чаще всего факторы делят на три группы.
1. Факторы неживой природы (абиотические, или физико-химические). К ним относятся климатические, атмосферные, почвенные (эдафические), геоморфологические (орографические), гидрологические и другие.
2. Факторы живой природы (биотические) - влияние одних организмов или их сообществ на другие. Эти влияния могут быть со стороны растений (фитогенные), животных (зоогенные), микроорганизмов, грибов и т. п.
3. Факторы человеческой деятельности (антропогенные). В их числе различают прямое влияние на организмы (например, промысел) и косвенное - влияние на местообитание (например, загрязнение среды, уничтожение кормовых угодий, строительство плотин на реках и т. п.).
Современные экологические проблемы и возрастающий интерес к экологии связан с действием антропогенных факторов.
Интересна классификация факторов по периодичности и направленности действия, степени адаптации к ним организмов. В этом отношении выделяют факторы, действующие строго периодически (смены времени суток, сезонов года, приливно-отливные явления и т. п.), действующие без строгой периодичности, но повторяющиеся время от времени. Сюда относятся погодные явления, наводнения, ураганы, землетрясения и т. п. Следующая группа - факторы направленного действия, они обычно изменяются в одном направлении (потепление или похолодание климата, зарастание водоемов, заболачивание территорий и т. п.). И последняя группа - факторы неопределенного действия. Сюда относятся антропогенные факторы, наиболее опасные для организмов и их сообществ.
Из перечисленных групп факторов организмы легче всего адаптируются или адаптированы к тем, которые четко изменяются (строго периодические, направленные). Адаптационность к ним такова, что часто становится наследственно обусловленной. И если фактор меняет периодичность, то организм продолжает в течение некоторого времени сохранять адаптации к нему, т. е. действовать в ритме так называемых «биологических часов». Такое явление, в частности, имеет место при смене часовых поясов.
Некоторые трудности характерны для адаптации к нерегулярно-периодическим факторам, но организмы нередко имеют механизмы предчувствия их возможности (землетрясения, ураганы, наводнения и т. п.) и в какой-то мере могут смягчать их отрицательные последствия.
Наибольшие трудности для адаптации представляют факторы, природа которых неопределенна, к ним организм, как правило, не готов, вид не встречался с такими явлениями и в процессе эволюции. Сюда, как отмечалось, относится группа антропогенных факторов. В этом их основная специфика и антиэкологичность. Многие из этих факторов, кроме того, выступают как вредные. Их относят к группе ксенобиотиков (греч. ксенокс - чужой). К последним относятся практически все загрязняющие вещества. В числе быстроизменяющихся факторов большое беспокойство в настоящее время вызывают изменение климата, обусловливаемое так называемым «тепличным, или парниковым, эффектом», изменение водных экосистем в результате преобразования рек, мелиорации и т. п. Только в отдельных случаях по отношению к таким факторам организмы могут использовать механизмы так называемых преадаптаций, т. е. те адаптации, которые выработались по отношению к другим факторам. Так, например, устойчивости растений к загрязнениям воздуха в какой-то мере способствуют те структуры, которые благоприятны для повышения засухоустойчивости: плотные покровные ткани листьев, наличие на них воскового налета, опушенности, меньшее количество устьиц и другие структуры, замедляющие процессы поглощения веществ, а следовательно, и отравление организма. Это необходимо учитывать, в частности, при подборе ассортимента видов для выращивания в районах с высокой промышленной нагрузкой, для озеленения городов, промплощадок и т.п.
II.2. Некоторые общие закономерности действия факторов среды на организмы
В комплексе действия факторов можно выделить некоторые закономерности, которые являются в значительной мере универсальными (общими) по отношению к организмам. К таким закономерностям относятся правило оптимума, правило взаимодействия факторов, правило лимитирующих факторов и некоторые другие.
Правило оптимума. В соответствии с этим правилом для экосистемы, организма или определенной стадии его развития имеется диапазон наиболее благоприятного (оптимального) значения фактора. За пределами зоны оптимума лежат зоны угнетения, переходящие в критические точки, за которыми существование невозможно (рис. 1).К зоне оптимума обычно приурочена максимальная плотность популяции. Зоны оптимума для различных организмов неодинаковы. Для одних они имеют значительный диапазон. Такие организмы относятся к группе эврибионтов (греч. эури - широкий; биос - жизнь). Организмы с узким диапазоном адаптации к факторам называются стенобионтами (греч. стенос - узкий). Важно подчеркнуть, что зоны оптимума по отношению к различным факторам различаются, и поэтому организмы полностью проявляют свои потенциальные возможности в том случае, если весь спектр факторов имеет для них оптимальные значения.
Диапазон значений факторов (между критическими точками) называют экологической валентностью (см. рис.1). Синонимом термина валентность является толерантность (лат. толеранция - терпение), или пластичность (изменчивость). Эти характеристики зависят в значительной мере от среды, в которой обитают организмы. Если она относительно стабильна по своим свойствам (малы амплитуды колебаний отдельных факторов), в ней больше стено-бионтов (например, в водной среде), если динамична, например, наземно-воздушная - в ней больше шансов на выживание имеют эврибионты.

Зона оптимума и экологическая валентность обычно шире у теплокровных организмов, чем у холоднокровных. Надо также иметь в виду, что экологическая валентность для одного и того же вида не остается одинаковой в различных условиях (например, в северных и южных районах в отдельные периоды жизни и т.п.). Молодые и старческие организмы, как правило, требуют более кондиционированных (однородных) условий. Иногда эти требования весьма неоднозначны. Например, по отношению к температуре личинки насекомых обычно стенобионтны (стенотермны), в то время как куколки и взрослые особи могут относиться к эврибионтам (эвритермным).
Правило взаимодействия факторов. Сущность его заключается в том, что одни факторы могут усиливать или смягчать силу действия других факторов. Например, избыток тепла может в какой-то мере смягчаться пониженной влажностью воздуха, недостаток света для фотосинтеза растений - компенсироваться повышенным содержанием углекислого газа в воздухе и т. п. Из этого, однако, не следует, что факторы могут взаимозаменяться. Они не взаимозаменяемы.
Правило лимитирующих факторов. Сущность этого правила заключается в том, что фактор, находящийся в недостатке или избытке (вблизи критических точек) отрицательно влияет на организмы и, кроме того, ограничивает возможность проявления силы действия других факторов, в том числе и находящихся в оптимуме. Например, если в почве имеются в достатке все, кроме одного, необходимые для растения химические элементы, то рост и развитие растения будет обусловливаться тем из них, который находится в недостатке. Все другие элементы при этом не проявляют своего действия. Лимитирующие факторы обычно обусловливают границы распространения видов (популяций), их ареалы. От них зависит продуктивность организмов и сообществ. Поэтому крайне важно своевременно выявлять факторы минимального и избыточного значения, исключать возможности их проявления (например, для растений - сбалансированным внесением удобрений).
Человек своей деятельностью часто нарушает практически все из перечисленных закономерностей действия факторов. Особенно это относится к лимитирующим факторам (разрушение местообитаний, нарушение режима водного и минерального питания растений и т.п.).
Фотопериодизм. Под фотопериодизмом понимают реакцию организма на длину дня (светлого времени суток). При этом длина светового дня выступает и как условие роста и развития, и как фактор-сигнал для наступления каких-то фаз развития или поведения организмов. Применительно к растениям обычно выделяют организмы короткого и длинного дня. Растения короткого дня существуют в низких (южных) широтах, где при длинном периоде вегетации день остается относительно коротким. Растения длинного дня характерны для высоких (северных) широт, где при коротком вегетационном периоде день длиннее, чем в южных широтах, вплоть до круглосуточного. Перемещение растении из одних широт в другие без учета данного явления обычно заканчивается неудачей: растения ненормально развиваются, не вызревают.
Сигнальное свойство фотопериодизма выражается в том, что растительные и животные организмы обычно реагируют на длину дня своим поведением, физиологическими процессами. Например, сокращение продолжительности дня является сигналом для подготовки организмов к зиме. Дня растений это повышение концентрации клеточного сока и т. п. Для животных - накопление жиров, смена накожных покровов, подготовка птиц к перелетам и т. п.
Другие факторы обычно в меньшей мере используются как сигнал (например, температура), поскольку они изменяются не с такой строгой закономерностью, как фотопериод, и могут провоцировать наступление у организмов каких-то фаз или явлений преждевременно или с запозданием. Хотя определенную корректировку в действие фотопериодизма они вносят.
Адаптации к ритмичности природных явлений. Наряду с длиной дня организмы эволюционно адаптировались к другим видам периодических явлений в природе. Прежде всего это относится к суточной и сезонной ритмике, приливно-отливным явлениям, ритмам, обусловливаемым солнечной активностью, лунными фазами и другими явлениями, повторяющимися со строгой периодичностью. Человек может нарушать эту ритмику через изменение среды, перемещением организмов в новые условия и другими действиями.
Ритмичность действия факторов среды, подверженная строгой периодичности, стала физиологически и наследственно обусловленной для многих организмов. Например, к суточной ритмике адаптирована активность многих животных организмов (интенсивность дыхания, частота сердцебиений, деятельность желез внутренней секреции и т п.). Одни организмы очень стойко сохраняют эту ритмику, другие более пластичны. Например, отмечается, что черные крысы более стойки к суточной (или околосуточной) ритмике и поэтому меньше склонны к расселению, держатся в определенных местообитаниях; серые крысы более лобильны по ритмике, легче осваивают новые условия и поэтому являются практически космополитами.
Индивидуальны реакции отдельных людей на изменение суточной ритмики. Например, одни лица относительно легко переносят смену часовых поясов, и для их адаптации в новых условиях требуется непродолжительное время. Другие - переносят такие смены болезненно и приспосабливаются к ним в течение более длительных периодов. Это явление представляет серьезную проблему с физиологической и медицинской точек зрения. В частности, при решении проблем ночных смен работы, пребывания в космосе, перелетах на значительные расстояния и т. п.
Поразительна высокая и разнообразная адаптивность некоторых организмов к подобным природным ритмам. Например, приливно-отливные ритмы морей связаны с солнечными сутками (24 часа), лунными сутками (24 часа 50 минут). Кроме этого, в течение последних имеют место два прилива и два отлива, которые ежедневно смещаются на 50 минут. Сила приливов изменяется также в течение лунного месяца, равного 29,5 солнечным суткам, а приливы дважды в месяц (при новолунии и полнолунии) достигают максимальной величины. Некоторые организмы, обитающие в приливно-отливной зоне (литораль), адаптируются ко всем изменениям водной среды. Например, отдельные рыбы (атерина в Калифорнии) откладывают икринки на границе максимального прилива. К этому же периоду приурочен и выход мальков из икринок.
Многие из ритмов становятся наследственно обусловленными. Например, при перемещении некоторых животных в более северные районы они (животные) продолжают сохранять свою ритмику. В таких случаях нарушается правило приуроченности наиболее ответственных периодов в жизни (размножения) к более благоприятному времени. Так, австралийские страусы в условиях Аскании Нова (Украина) могут откладывать яйца на снег.
Нет оснований доказывать, что ритмичность деятельности организмов должна учитываться человеком при тех или иных изменениях среды и особенно при перемещениях или переселениях организмов, например, при интродукции (перемещении вида в новые условия за пределы его ареала).
II.3. Среды жизни и адаптации к ним организмов
Наряду с понятиями «среда», «местообитание», «природная среда», «окружающая среда» широко используется термин «среда жизни». Все разнообразие условий на Земле объединяют в четыре среды жизни: водную, наземно-воздушную, почвенную и организменную (в последнем случае одни организмы являются средой для других).
Среды жизни выделяются обычно по фактору или комплексу факторов, которые никогда не бывают в недостатке. Эти факторы являются средообразующими и обусловливают свойства сред. Рассмотрим кратко присущие названным средам жизни свойства, лимитирующие факторы и адаптации организмов.
Водная среда. Эта среда наиболее однородна среди других. Она мало изменяется в пространстве, здесь нет четких границ между отдельными экосистемами. Амплитуды значений факторов также невелики. Разница между максимальными и минимальными значениями температуры здесь обычно не превышает 50°С (в наземно-воздушной среде - до 100°С). Среде присуща высокая плотность. Для океанических вод она равна 1,3 г/см3, для пресных - близка к единице. Давление изменяется только в зависимости от глубины: каждый 10-метровый слой воды увеличивает давление на 1 атмосферу.
Лимитирующим фактором часто бывает кислород. Содержание его обычно не превышает 1% от объема. При повышении температуры, обогащении органическим веществом и слабом перемешивании содержание кислорода в воде уменьшается. Малая доступность кислорода для организмов связана также с его слабой диффузией (в воде она в тысячи раз меньше, чем в воздухе). Второй лимитирующий фактор - свет. Освещенность быстро уменьшается с глубиной. В идеально чистых водах свет может проникать до глубины 50-60 м, в сильно загрязненных - только на несколько сантиметров.
В воде мало теплокровных, или гомойотермных (греч. хомой -одинаковый, термо - тепло), организмов. Это результат двух причин: малое колебание температур и недостаток кислорода. Основной адаптационный механизм гомойотермии - противостояние неблагоприятным температурам. В воде такие температуры маловероятны, а в глубинных слоях температура практически постоянна (+4°С). Поддержание постоянной температуры тела обязательно связано с интенсивными процессами обмена веществ, что возможно только при хорошей обеспеченности кислородом. В воде таких условий нет. Теплокровные животные водной среды (киты, тюлени, морские котики и др.) - это бывшие обитатели суши. Их существование невозможно без периодической связи с воздушной средой.
Типичные обитатели водной среды имеют переменную температуру тела и относятся к группе пойкилотермных (греч. пойкиос - разнообразный). Недостаток кислорода они в какой-то мере компенсируют увеличением соприкосновения органов дыхания с водой. Многие обитатели вод (гидробионты) потребляют кислород через все покровы тела. Часто дыхание сочетается с фильтрационным типом питания, при котором через организм пропускается большое количество воды. Некоторые организмы в периоды острого недостатка кислорода способны резко замедлять жизнедеятельность, вплоть до состояния анабиоза (почти полное прекращение обмена веществ).
К высокой плотности воды организмы адаптируются в основном двумя путями. Одни используют ее как опору и находятся в состоянии свободного парения. Плотность (удельный вес) таких организмов обычно мало отличается от плотности воды. Этому способствует полное или почти полное отсутствие скелета, наличие выростов, капелек жира в теле или воздушных полостей. Такие организмы объединяются в группу планктона (греч. планктос -блуждающий). Различают растительный (фито-) и животный (зоо-) планктон. Размеры планктонных организмов обычно невелики. Но на их долю приходится основная масса водных обитателей.
Активно передвигающиеся организмы (пловцы) адаптируются к преодолению высокой плотности воды. Для них характерна продолговатая форма тела, хорошо развитая мускулатура, наличие структур, уменьшающих трение (слизь, чешуя). В целом же высокая плотность воды имеет следствием уменьшение доли скелета в общей массе тела гидробионтов по сравнению с наземными организмами.
В условиях недостатка света или его отсутствия организмы для ориентации используют звук. Он в воде распространяется намного быстрее, чем в воздухе. Для обнаружения различных препятствий используется отраженный звук по типу эхолокации. Для ориентации используются также запаховые явления (в воде запахи ощущаются намного лучше, чем в воздухе). В глубинах вод многие организмы обладают свойством самосвечения (биолюминесценции).
Растения, обитающие в толще воды, используют в процессе фотосинтеза наиболее глубоко проникающие в воду голубые, синие и сине-фиолетовые лучи. Соответственно и цвет растений меняется с глубиной от зеленого к бурому и красному.
Адекватно адаптационным механизмам выделяются следующие группы гидробионтов: отмеченный выше планктон - свободнопарящие, нектон (греч. нектос - плавающий) - активно передвигающиеся, бентос (греч. бентос - глубина) - обитатели дна, пелагос (греч. пелагос - открытое море) - обитатели водной толщи, нейстон - обитатели верхней пленки воды (часть тела может быть в воде, часть - в воздухе).
Воздействие человека на водную среду проявляется в уменьшении прозрачности, изменении химического состава (загрязнении) и температуры (тепловое загрязнение). Следствием этих и других воздействий является обеднение кислородом, снижение продуктивности, смены видового состава и другие отклонения от нормы. Подробнее эти вопросы рассматриваются в ч. II работы (разд.VII, VII.5).
Наземно-воздушная среда. Эта среда относится к наиболее сложной как по свойствам, так и по разнообразию в пространстве. Для нее характерна низкая плотность воздуха, большие колебания температуры (годовые амплитуды до 100°С), высокая подвижность атмосферы. Лимитирующими факторами чаще всего являются недостаток или избыток тепла и влаги. В отдельных случаях, например под пологом леса, недостаток света.
Большие колебания температуры во времени и ее значительная изменчивость в пространстве, а также хорошая обеспеченность кислородом явились побудительными мотивами для появления организмов с постоянной температурой тела (гомойотермных). Гомойотермия позволила обитателям суши существенно расширить место обитания (ареалы видов), но это неизбежно связано с повышенными энергетическими тратами.
Для организмов наземно-воздушной среды типичны три механизма адаптации к температурному фактору: физический, химический, поведенческий. Физический осуществляется регулированием теплоотдачи. Факторами ее являются кожные покровы, жировые отложения, испарение воды (потовыделение у животных, транспирация у растений). Этот путь характерен для пойкилотермных и гомойотермных организмов. Химические адаптации базируются на поддержании определенной температуры тела. Это требует интенсивного обмена веществ. Такие адаптации свойственны гомойотермным и лишь частично пойкилотермным организмам. Поведенческий путь осуществляется посредством выбора организмами предпочтительных положений (открытые солнцу или затененные места, разного вида укрытия и т. п.). Он свойственен обеим группам организмов, но пойкилотермным в большей степени. Растения приспосабливаются к температурному фактору в основном через физические механизмы (покровы, испарение воды) и лишь частично - поведенчески (повороты пластинок листьев относительно солнечных лучей, использование тепла земли и утепляющей роли снежного покрова).
Адаптации к температуре осуществляются также через размеры и форму тела организмов. Для уменьшения теплоотдачи выгоднее крупные размеры (чем крупнее тело, тем меньше его поверхность на единицу массы, а следовательно, и теплоотдача, и наоборот). По этой причине одни и те же виды, обитающие в более холодных условиях (на севере), как правило, крупнее тех, которые обитают в более теплом климате. Эта закономерность называется правилом Бергмана. Регулирование температуры осуществляется также через выступающие части тела (ушные раковины, конечности, органы обоняния). В холодных районах они, как правило, меньше по размерам, чем в более теплых (правило Аллена).
О зависимости теплоотдачи от размеров тела можно судить по количеству кислорода, расходуемого при дыхании на единицу массы различными организмами. Оно тем больше, чем меньше размеры животных. Так, на 1 кг массы потребление кислорода (смУчас) составило: лошадь - 220, кролик - 480, крыса -1800, мышь - 4100.
Регулирование водного баланса организмами. У животных различают три механизма: морфологический - через форму тела, покровы; физиологический - посредством высвобождения воды из жиров, белков и углеводов (метаболическая вода), через испарение и органы выделения; поведенческий - выбор предпочтительного расположения в пространстве.
Растения избегают обезвоживания либо посредством запасания воды в теле и защиты ее от испарения (суккуленты), либо через увеличение доли подземных органов (корневых систем) в общем объеме тела. Уменьшению испарения способствуют также различного рода покровы (волоски, плотная кутикула, восковой налет и др.). При избытке воды механизмы ее экономии слабо выражены. Наоборот, некоторые растения способны выделять избыточную воду через листья, в капельно-жидком виде («плач растений»).
Воздействия человека на наземно-воздушную среду и ее обитателей многообразны. Они рассматриваются во второй части работы.
Почвенная среда. Эта среда имеет свойства, сближающие ее с водной и наземно-воздушной средами.
Многие мелкие организмы живут здесь как гидробионты - в поровых скоплениях свободной воды. Как и в водной среде, в почвах невелики колебания температур. Амплитуды их быстро затухают с глубиной. Существенна вероятность дефицита кислорода, особенно при избытке влаги или углекислоты. Сходство с наземно-воздушной средой проявляется через наличие пор, заполненных воздухом.
К специфическим свойствам, присущим только почве, относится плотное сложение (твердая часть или скелет). В почвах обычно выделяют три фазы (части): твердую, жидкую и газообразную. В. И. Вернадский почву отнес к биокосным телам, подчеркивая этим большую роль в ее образовании и жизни организмов и продуктов их жизнедеятельности. Почва- наиболее насыщенная живыми организмами часть биосферы (почвенная пленка жизни). Поэтому в ней иногда выделяют четвертую фазу - живую.
Есть основание рассматривать почву как среду, которая играла промежуточную роль при выходе организмов из воды на сушу (М. С. Гиляров). Кроме перечисленных выше свойств, сближающих эти среды, в почве организмы находили защиту от жесткого космического излучения (при отсутствии озонового экрана).
В качестве лимитирующих факторов в почве чаще всего выступает недостаток тепла (особенно при вечной мерзлоте), а также недостаток (засушливые условия) или избыток (болота) влаги. Реже лимитирующими бывают недостаток кислорода или избыток угае-кислоты.
Жизнь многих почвенных организмов тесно связана с порами и их размером. Одни организмы в порах свободно передвигаются. Другие (более крупные организмы) при передвижении в порах изменяют форму тела по принципу перетекания, например дождевой червь, или уплотняют стенки пор. Третьи могут передвигаться только разрыхляя почву или выбрасывая на поверхность образующий ее материал (землерои). Из-за отсутствия света многие почвенные организмы лишены органов зрения. Ориентация осуществляется с помощью обоняния или других рецепторов.
Воздействия человека проявляются в разрушении почв (эрозии), загрязнении, изменении химических и физических свойств. Эти вопросы рассматриваются в ч. II работы (ra.VIII).
Организмы как среда обитания. С данной средой связан паразитический и полупаразитический образ жизни. Организмы этих групп получают кондиционированную среду (по температуре, влажности и другим параметрам) и готовую легкоусвояемую пищу. Результатом этого является упрощение всех систем и органов, а также потеря некоторых из них. Наиболее слабое (лимитирующее) звено в жизни паразитов - возможность потери хозяина. Это неизбежно при его смерти. По этой причине паразиты, как правило, не убивают своего хозяина («разумный паразитизм») и имеют приспособления, увеличивающие вероятность выживания в случае потери хозяина. Основной путь сохранения вида (популяции) в таких условиях - большое число зачатков («закон большого числа яиц») в виде долгосохраняющихся цист, спор и т. п. Это увеличивает вероятность встречи с хозяином. Часто используются промежуточные хозяины.
Человек может как увеличивать, так и уменьшать численность паразитов, воздействуя как на среду для организмов-хозяинов, так и непосредственно на последних. Используются различные методы прямого уничтожения или ограничения численности паразитов.
Вопросы и задания
1. В каких сочетаниях используется термин «среда»? В чем отличие этих сочетаний?
2. Что понимается под экологическим фактором? Приведите классификации факторов по двум известным Вам принципам. Какие факторы являются наиболее трудными для адаптации к ним организмов?
3. Перечислите общие закономерности действия факторов среды на организмы. Раскройте их сущность и значение.
4. Перечислите среды жизни и наиболее типичные их свойства. Назовите присущие отдельным средам жизни лимитирующие факторы, адаптации организмов.
III. БИОСФЕРА
III.1. Биосфера как глобальная экосистема
Рассмотрение основополагающих вопросов общей экологии целесообразно начать со знакомства с биосферой как глобальной экосистемой и закономерностями ее функционирования. В таком случае разделы экологии более низкого ранга (популяционный, экосистемный) будут в определенной мере подчинены углубленному раскрытию закономерностей существования биосферы и допустимых пределов вмешательства в нее или ее звенья человека. Другими словами, на уровне экосистем элементарного плана должен осуществляться в основном принцип локальных действий, в то время как биосферный уровень формирует базу для глобального мышления.
В настоящее время становится предельно ясным, что среда, в которой мы живем, сформирована жившими организмами различных геологических эпох. По образному выражению Б. Коммонера, окружающая среда - «... это дом, созданный на Земле жизнью и для жизни». При этом каждое поколение организмов этот дом совершенствовало соответственно изменявшимся условиям и обитающим в нем существам. Эти истины стали понятными людям далеко не сразу. Важнейший вклад в этот раздел современной экологии внесли исследования академика В. И. Вернадского (1863-1945), его учение о биосфере.
Понятие «биосфера». Термин «биосфера» в научную литературу введен в 1875 г. австрийским ученым-геологом Эдуардом Зюссом. К биосфере он отнес все то пространство атмосферы, гидросферы и литосферы (твердой оболочки Земли), где встречаются живые организмы.
Владимир Иванович Вернадский (1863-1945) использовал этот термин и создал науку с аналогичным названием. Если с понятием «биосфера», по Зюссу, связывалось только наличие в трех сферах земной оболочки (твердой, жидкой и газообразной) живых организмов, то, по В. И. Вернадскому, им отводится роль главнейшей геохимической силы. При этом в понятие биосферы включается преобразующая деятельность организмов не только в границах распространения жизни в настоящее время, но и в прошлом. В таком случае под биосферой понимается все пространство (оболочка Земли), где существует или когда-либо существовала жизнь, то есть где встречаются живые организмы или продукты их жизнедеятельности. В. И. Вернадский не только сконкретизировал и очертил границы жизни в биосфере, но, самое главное, всесторонне раскрыл роль живых организмов в процессах планетарного масштаба. Он показал, что в природе нет более мощной геологической (средообразующей) силы, чем живые организмы и продукты их жизнедеятельности.
Учение В. И. Вернадского о биосфере произвело переворот во взглядах на глобальные природные явления, в том числе геологические процессы, причины явлений, их эволюцию. До трудов В. И. Вернадского эти процессы прежде всего связывались с действием физико-химических сил, объединяемых термином «выветривание». В. И. Вернадский показал первостепенную преобразующую роль живых организмов и обусловливаемых ими механизмов образования и разрушения геологических структур, круговорота веществ, изменения твердой (литосферы), водной (гидросферы) и воздушной (атмосферы) оболочек Земли.
Ту часть биосферы, где живые организмы встречаются в настоящее время, обычно называют современной биосферой, или необиосферой, а древние биосферы относят к палеобиосферам, или белым биосферам. В качестве примеров последних можно назвать безжизненные скопления органических веществ (залежи каменных углей, нефти, горючих сланцев и т. п.) или запасы других соединений, образовавшихся при участии живых организмов (известь, мел, соединения кремния, рудные образования и т. п.).
Границы биосферы. По современным представлениям необиосфера в атмосфере простирается примерно до озонового экрана (у полюсов 8-10 км, у экватора - 17-18 км и над остальной поверхностью Земли - 20-25 км). За пределами озонового слоя жизнь невозможна вследствие наличия губительных космических ультрафиолетовых лучей. Гидросфера практически вся, в том числе и самая глубокая впадина (Марианская) Мирового океана (11022 м), занята жизнью. К необиосфере следует относить также и донные отложения, где возможно существование живых организмов. В литосферу жизнь проникает на несколько метров, ограничиваясь в основном почвенным слоем, но по отдельным трещинам и пещерам она распространяется на сотни метров.
Границы палеобиосферы в атмосфере примерно совпадают с необиосферой, под водами к палеобиосфере следует отнести и осадочные породы, которые, по В. И. Вернадскому, практически все претерпели переработку живыми организмами. Это толща от сотен метров до десятков километров. Сказанное относительно осадочных пород применимо и к литосфере, пережившей водную стадию функционирования.
Таким образом, границы биосферы определяются наличием живых организмов или «следами» их жизнедеятельности. В пределах современной, как и былых биосфер, насыщенность жизнью между тем далеко не равномерна. На границах биосферы встречаются лишь случайно занесенные организмы («поле устойчивости жизни», по В. И. Вернадскому). В пределах основной части биосферы организмы присутствуют постоянно («поле существования жизни»), но распределены далеко не равномерно. Очаги повышенной и максимальной концентрации жизни В. И. Вернадский называл пленками и сгущениями жизни. Эти наиболее продуктивные экосистемы являются своего рода каркасом биосферы и требуют повышенного внимания человека. Подробнее о пленках и сгущениях жизни см. разд. IV.5 (ч.1).
III.2. Живое вещество, его средообразующие свойства и функции в биосфере
Живое вещество. Этот термин введен в литературу В. И. Вернадским. Под ним он понимал совокупность всех живых организмов, выраженную через массу, энергию и химический состав.
Вещества неживой природы относятся к косным (например, минералы). В природе, кроме этого, довольно широко представлены биокосные вещества, образование и сложение которых обусловливается живыми и косными составляющими (например, почвы, воды).
Живое вещество - основа биосферы, хотя и составляет крайне незначительную ее часть. Если его выделить в чистом виде и распределить равномерно по поверхности Земли, то это будет слой около 2 см или крайне незначительная доля от объема всей биосферы, толща которой измеряется десятками километров. В чем же причина столь высокой химической активности и геологической роли живого вещества?
Прежде всего это связано с тем, что живые организмы, благодаря биологическим катализаторам (ферментам), совершают, по выражению академика Л. С. Берга, с физико-химической точки зрения что-то невероятное. Например, они способны фиксировать в своем теле молекулярный азот атмосферы при обычных для природной среды значениях температуры и давления. В промышленных условиях связывание атмосферного азота до аммиака требует температуры порядка 500°С и давления 300-500 атмосфер.
В живых организмах на порядок или несколько порядков увеличиваются скорости химических реакций в процессе обмена веществ. В. И. Вернадский в связи с этим живое вещество назвал чрезвычайно активизированной материей.
Свойства живого вещества. К основным уникальным особенностям живого вещества, обусловливающим его крайне высокую средообразующую деятельность, можно отнести следующие:
1. Способность быстро занимать (осваивать) все свободное пространство. В. И. Вернадский назвал это всюдностью жизни. Данное свойство дало основание В. И. Вернадскому сделать вывод, что для определенных геологических периодов количество живого вещества было примерно постоянным (константой). Способность быстрого освоения пространства связана как с интенсивным размножением (некоторые простейшие формы организмов могли бы освоить весь земной шар за несколько часов или дней, если бы не было факторов, сдерживающих их потенциальные возможности размножения), так и со способностью организмов интенсивно увеличивать поверхность своего тела или образуемых ими сообществ. Например, площадь листьев растений, произрастающих на 1 га, составляет 8-10 га и более. То же относится к корневым системам.
2. Движение не только пассивное (под действием силы тяжести, гравитационных сил и т. п.), но и активное. Например, против течения воды, силы тяжести, движения воздушных потоков и т. п.
3. Устойчивость при жизни и быстрое разложение после смерти (включение в круговороты), сохраняя при этом высокую физико-химическую активность.
4. Высокая приспособительная способность (адаптация) к различным условиям и в связи с этим освоение не только всех сред жизни (водной, наземно-воздушной, почвенной, организменной), но и крайне трудных по физико-химическим параметрам условий. Например, некоторые организмы выносят температуры, близкие к значениям абсолютного нуля - 273°С, микроорганизмы встречаются в термальных источниках с температурами до 140°С, в водах атомных реакторов, в бескислородной среде, в ледовых панцирях и т. п.
5. Феноменально высокая скорость протекания реакций. Она на несколько порядков (в сотни, тысячи раз) значительнее, чем в неживом веществе. Об этом свойстве можно судить по скорости переработки вещества организмами в процессе жизнедеятельности. Например, гусеницы некоторых насекомых потребляют за день количество пищи, которое в 100-200 раз больше веса их тела. Особенно активны организмы-грунтоеды. Дождевые черви (масса их тел примерно в 10 раз больше биомассы всего человечества) за 150-200 лет пропускают через свои организмы весь однометровый слой почвы. Такие же явления имеют место в донных отложениях океана. Слой донных отложений здесь может быть представлен продуктами жизнедеятельности кольчатых червей (полихет) и достигать нескольких метров. Колоссальную роль по преобразованию вещества выполняют организмы, для которых характерен фильтрационный тип питания. Они освобождают водные массы от взвесей, склеивая их в небольшие агрегаты и осаждая на дно.
Впечатляют примеры чисто механической деятельности некоторых организмов, например роющих животных (сурков, сусликов и др.), которые в результате переработки больших масс грунта создают своеобразный ландшафт. По представлениям В. И. Вернадского, практически все осадочные породы, а это слой до 3 км, на 95-99% переработаны живыми организмами. Даже такие колоссальные запасы воды, которые имеются в биосфере, разлагаются в процессе фотосинтеза за 5-6 млн. лет, углекислота же проходит через живые организмы в процессе фотосинтеза каждые 6-7 лет.
6. Высокая скорость обновления живого вещества. Подсчитано, что в среднем для биосферы она составляет 8 лет, при этом для суши -14 лет, а для океана, где преобладают организмы с коротким периодом жизни (например, планктон), - 33 дня. В результате высокой скорости обновления за всю историю существования жизни общая масса живого вещества, прошедшего через биосферу, примерно в 12 раз превышает массу Земли. Только небольшая часть его (доли процента) законсервирована в виде органических остатков (по выражению В. И. Вернадского, «ушла в геологию»), остальная же включилась в процессы круговорота.
Все перечисленные и другие свойства живого вещества обусловливаются концентрацией в нем больших запасов энергии. Согласно В. И. Вернадскому, по энергетической насыщенности с живым веществом может соперничать только лава, образующаяся при извержении вулканов.
Средообразующие функции живого вещества. Всю деятельность живых организмов в биосфере можно, с определенной долей условности, свести к нескольким основополагающим функциям, которые позволяют значительно дополнить представление об их преобразующей биосферно-геологической роли.
В. И. Вернадский выделял девять функций живого вещества: газовую, кислородную, окислительную, кальциевую, восстановительную, концентрационную и другие. В настоящее время название этих функций несколько изменено, некоторые из них объединены. Мы приводим их в соответствии с классификацией А. В. Лапо (1987).
1. Энергетическая. Связана с запасанием энергии в процессе фотосинтеза, передачей ее по цепям питания, рассеиванием. Эта функция - одна из важнейших и будет подробнее рассмотрена в разделе IV.4 - энергетика экосистем.
Энергетическая функция живого вещества нашла отражение в двух биогеохимических принципах, сформулированных В.И.Вернадским. В соответствии с первым из них геохимическая биогенная энергия стремится в биосфере к максимальному проявлению. Второй принцип гласит, что в процессе эволюции выживают те организмы, которые своей жизнью увеличивают геохимическую энергию.
2. Газовая - способность изменять и поддерживать определенный газовый состав среды обитания и атмосферы в целом. В частности, включение углерода в процессы фотосинтеза, а затем в цепи питания обусловливало аккумуляцию его в биогенном веществе (органические остатки, известняки и т. п.) В результате этого шло постепенное уменьшение содержания углерода и его соединений, прежде всего двуокиси (СО2) в атмосфере с десятков процентов до современных 0,03%. Это же относится к накоплению в атмосфере кислорода, синтезу озона и другим процессам.
С газовой функцией в настоящее время связывают два переломных периода (точки) в развитии биосферы. Первая из них относится ко времени, когда содержание кислорода в атмосфере достигло примерно 1% от современного уровня (первая точка Пастера). Это обусловило появление первых аэробных организмов (способных жить только в среде, содержащей кислород). С этого времени восстановительные процессы в биосфере стали дополняться окислительными. Это произошло примерно 1,2 млрд. лет назад. Второй переломный период в содержании кислорода связывают со временем, когда концентрация его достигла примерно 10% от современной (вторая точка Пастера). Это создало условия для синтеза озона и образования озонового экрана в верхних слоях атмосферы, что обусловило возможность освоения организмами суши (до этого функцию защиты организмов от губительных ультрафиолетовых лучей выполняла вода, под слоем которой возможна была жизнь).
3. Окислительно-восстановительная. Связана с интенсификацией под влиянием живого вещества процессов как окисления, благодаря обогащению среды кислородом, так и восстановления прежде всего в тех случаях, когда идет разложение органических веществ при дефиците кислорода. Восстановительные процессы обычно сопровождаются образованием и накоплением сероводорода, а также метана. Это, в частности, делает практически безжизненными глубинные слои болот, а также значительные придонные толщи воды (например, в Черном море). Данный процесс в связи с деятельностью человека прогрессирует.
4. Концентрационная - способность организмов концентрировать в своем теле рассеянные химические элементы, повышая их содержание по сравнению с окружающей организмы средой на несколько порядков (по марганцу, например, в теле отдельных организмов - в миллионы раз). Результат концентрационной деятельности - залежи горючих ископаемых, известняки, рудные месторождения и т. п. Эту функцию живого вещества всесторонне изучает наука биоминералогия. Организмы-концентраторы используются для решения конкретных прикладных вопросов, например для обогащения руд интересующими человека химическими элементами или соединениями.
5. Деструктивная - разрушение организмами и продуктами их жизнедеятельности как самих остатков органического вещества, так и косных веществ. Основной механизм этой функции связан с круговоротом веществ. Наиболее существенную роль в этом отношении выполняют низшие формы жизни - грибы, бактерии (деструкторы, редуценты).
6. Транспортная - перенос вещества и энергии в результате активной формы движения организмов. Часто такой перенос осуществляется на колоссальные расстояния, например, при миграциях и кочевках животных. С транспортной функцией в значительной мере связана концентрационная роль сообществ организмов, например, в местах их скопления (птичьи базары и другие колониальные поселения).
7. Средообразующая. Эта функция является в значительной мере интегративной (результат совместного действия других функций). С ней в конечном счете связано преобразование физико-химических параметров среды. Эту функцию можно рассматривать в широком и более узком планах.
В широком понимании результатом данной функции является вся природная среда. Она создана живыми организмами, они же и поддерживают в относительно стабильном состоянии ее параметры практически во всех геосферах.
В более узком плане средообразующая функция живого вещества проявляется, например, в образовании почв. В. И. Вернадский, как отмечалось выше, почву называл биокосным телом, подчеркивая тем самым большую роль живых организмов в ее создании и существовании. Роль живых организмов в образовании почв убедительно показал Ч. Дарвин в работе «Образование растительного слоя земли деятельностью дождевых червей». Известный ученый В. В. Докучаев назвал почву «зеркалом ландшафта», подчеркивая тем самым, что она продукт основного ландшафтообразующего элемента - биоценозов и, прежде всего, растительного покрова.
Локальная средообразующая деятельность живых организмов и особенно их сообществ проявляется также в трансформации ими метеорологических параметров среды. Это прежде всего относится к сообществам с большой массой органического вещества (биомассой). Например, в лесных сообществах микроклимат существенно отличается от открытых (полевых) пространств. Здесь меньше суточные и годовые колебания температур, выше влажность воздуха, ниже содержание углекислоты в атмосфере на уровне полога, насыщенного листьями (результат фотосинтеза), и повышенное ее количество в припочвенном слое (следствие интенсивно идущих процессов разложения органического вещества на почве и в верхних горизонтах почвы).
8. Наряду с концентрационной функцией живого вещества выделяется противоположная ей по результатам - рассеивающая. Она проявляется через трофическую (питательную) и транспортную деятельность организмов. Например, рассеивание вещества при выделении организмами экскрементов, гибели организмов при разного рода перемещениях в пространстве, смене покровов. Железо гемоглобина крови рассеивается, например, кровососущими насекомыми и т. п.
Важна также информационная функция живого вещества, выражающаяся в том, что живые организмы и их сообщества накапливают определенную информацию, закрепляют ее в наследственных структурах и затем передают последующим поколениям. Это одно из проявлений адаптационных механизмов.
В обобщающем виде роль живого вещества сформулирована геохимиком А. Н. Перельманом в виде «Закона биогенной миграции атомов» (В. И. Вернадского): «Миграция химических элементов на земной поверхности и в биосфере в целом осуществляется или при непосредственном участии живого вещества, или же она протекает в среде, геохимические особенности которой обусловлены живым веществом...» В соответствии с этим законом понимание процессов, протекающих в биосфере, невозможно без учета биотических и биогенных факторов. Воздействуя на живое население Земли, люди тем самым изменяют условия миграции атомов, а следовательно, воздействуют на основополагающие геологические процессы.
III.3. Основные свойства биосферы
Биосфере, как и составляющим ее другим экосистемам более низкого ранга, присуща система свойств, которые обеспечивают ее функционирование, саморегулирование, устойчивость и другие параметры. Рассмотрим основные из них.
1. Биосфера - централизованная система. Центральным звеном ее выступают живые организмы (живое вещество). Это свойство всесторонне раскрыто В. И. Вернадским, но, к сожалению, часто не дооценивается человеком и в настоящее время: в центр биосферы или ее звеньев ставится только один вид - человек (антропоцентризм).
2. Биосфера - открытая система. Ее существование немыслимо без поступления энергии извне. Она испытывает воздействие космических сил, прежде всего солнечной активности. Впервые представления о влиянии солнечной активности на живые организмы (гелиобиология) разработаны А. Л. Чижевским (1897-1964), который показал, что многие явления на Земле и в биосфере тесно связаны с активностью солнца. Все больше накапливается данных, свидетельствующих, что резкое увеличение численности отдельных видов или популяций («волны жизни») - результат изменения солнечной активности. Высказываются мнения, что солнечная активность оказывает воздействие на многие геологические процессы (катаклизмы, катастрофы), а также на социальную активность человеческого общества или отдельных его этносов.
В частности, есть сторонники той точки зрения, что серия аномальных явлений, имевших место, например, в 1989 году, связана с высокой солнечной активностью. На протяжении только 1,5-2 месяцев наблюдались такие аномальные явления, как землетрясение на острове Итуруп, авария на продуктопроводе в районе Челябинска, гибель атомной подводной лодки «Комсомолец», события в Тбилиси, активизация военных действий в Нагорном Карабахе и др.
3. Биосфера - саморегулирующаяся система, для которой, как отмечал В. И. Вернадский, характерна организованность. В настоящее время это свойство называют гомеостазом, понимая под ним способность возвращаться в исходное состояние, гасить возникающие возмущения включением ряда механизмов. Гомеостатические механизмы связаны в основном с живым веществом, его свойствами и функциями, рассмотренными выше. Биосфера за свою историю пережила ряд таких возмущений, многие из которых были значительными по масштабам, и справлялась с ними (извержения вулканов, встречи с астероидами, землетрясения, горообразование и т. п.) благодаря действию гомеостатических механизмов и, в частности, принципа, который в настоящее время носит название Ле Шателье-Брауна: при действии на систему сил, выводящих ее из состояния устойчивого равновесия, последнее смещается в том направлении, при котором эффект этого воздействия ослабляется.
Опасность современной экологической ситуации связана прежде всего с тем, что нарушаются многие механизмы гомеостаза и принцип Ле Шателье-Брауна, если не в планетарном, то в крупных региональных планах. Их следствие - региональные кризисы. В стадию глобального кризиса биосфера, к счастью, еще, по-видимому, не вступила. Но отдельные крупные возмущения она уже гасить не в силах. Результатом этого является либо распад экосистем (например, расширяющиеся площади опустыненных земель), либо появление неустойчивых, практически лишенных свойств гомеостаза систем типа агроценозов или урбанизированных (городских) комплексов. Человечеству, к сожалению, отпущен крайне малый промежуток" времени для того, чтобы не произошел глобальный кризис и следующие за ним катастрофы и коллапс (полный и необратимый распад системы).
4. Биосфера - система, характеризующаяся большим разнообразием. Разнообразие - важнейшее свойство всех экосистем. Биосфера как глобальная экосистема характеризуется максимальным среди других систем разнообразием. Последнее обусловливается многими причинами и факторами. Это и разные среды жизни (водная, наземно-воз-душная, почвенная, организменная); и разнообразие природных зон, различающихся по климатическим, гидрологическим, почвенным, биотическим и другим свойствам; и наличие регионов, различающихся по химическому составу (геохимические провинции); и, самое главное, объединение в рамках биосферы большого количества элементарных экосистем со свойственным им видовым разнообразием.
В настоящее время описано около 2 млн. видов (примерно 1,5 млн. животных и 0,5 млн. растений). Полагают, однако, что число видов на Земле в 2-3 раза больше, чем их описано. Не учтены многие насекомые и микроорганизмы, особенно в тропических лесах, глубинных частях океанов и в других малоосвоенных местообитаниях. Кроме этого, современный видовой состав - это лишь небольшая часть видового разнообразия, которое принимало участие в процессах биосферы за период ее существования. Дело в том, что каждый вид имеет определенную продолжительность жизни (10-30 млн. лет), и поэтому с учетом постоянной смены и обновления видов число видов, принимавших участие в становлении биосферы, исчисляется сотнями миллионов. Считается, что к настоящему времени арену биосферы оставили более 95% видов.
Разнообразие биосферы за счет элементарных экосистем по вертикали обусловливается ярусностью или экогоризонтами растительного покрова и связанных с ними животных организмов, а в горизонтальном направлении неравномерностью распределения организмов и их группировок и связанных с ними факторов (увлажнение, микрорельеф, обеспеченность элементами питания и т. п.).
Для любой природной системы разнообразие - одно из важнейших ее свойств. С ним связана возможность дублирования, подстраховки, замены одних звеньев другими (например, на видовом или популяционном уровнях), степень сложности и прочности пищевых и других связей. Поэтому разнообразие рассматривают как основное условие устойчивости любой экосистемы и биосферы в целом. Это свойство настолько универсально, что сформулировано в качестве закона (автор его У. Р. Эшби).
К сожалению, как будет показано во второй части работы, практически вся без исключения деятельность человека подчинена упрощению экосистем любого ранга. Сюда следует отнести и уничтожение отдельньк видов или резкое уменьшение их численности, и создание агроценозов на месте сложных природных систем. Например, полностью исчезли с лица земли степи как тип экосистем и ландшафтов, резко уменьшились площади лесов (до появления человека они занимали примерно 70% суши, а сейчас - не более 20-23%). Идет дальнейшее, невиданное по масштабам уничтожение лесных экосистем в настоящее время, особенно наиболее ценных и сложных тропических, спрямление русел рек, создание промышленных районов и т. п.
Простые экосистемы с малым разнообразием удобны для эксплуатации, они позволяют в короткое время получить значительный объем нужной продукции (например, с сельскохозяйственных полей), но за это приходится рассчитываться снижением устойчивости экосистем, их распадом и деградацией среды.
Не случайно, что биологическое разнообразие отнесено Конференцией ООН по окружающей среде и развитию (1992 г.) к числу трех важнейших экологических проблем, по которым приняты специальные Заявления или Конвенции. Кроме сохранения разнообразия, такие конвенции приняты по сохранению лесов и по предотвращению изменений климата.
Следует подчеркнуть, что значимость разнообразия для природных систем в значительной мере действительна и для социальных структур. Всякое стремление к упрощению социальной структуры общества, перевод ее на однообразие, авторитаризм могут дать кратковременный положительный результат, за которым неминуемо проявляются отрицательные последствия.
Важное свойство биосферы - наличие в ней механизмов, обеспечивающих круговорот веществ и связанную с ним неисчерпаемость отдельных химических элементов и их соединений. При отсутствии круговорота, например, за короткое время был бы исчерпан основной «строительный материал» живого - углерод, который практически единственный способен образовывать межэлементные (углерод-углеродные) связи и создавать огромное количество органических соединений. Только благодаря круговоротам и наличию неисчерпаемого источника солнечной энергии обеспечивается непрерывность процессов в биосфере и ее потенциальное бессмертие. Как отмечал академик-почвовед В. Р. Вильямс, есть единственный способ сделать какой-то процесс бесконечным - пустить его по пути круговоротов. Одно из мощнейших антиэкологических действий человека связано с нарушением и даже разрушением природных круговоротов. Эти вопросы будут рассмотрены во второй части работы (см.разд.II.2).
Вопросы и задания
1. Что понимается под «Биосферой»? Кто является автором термина и автором науки «Биосфера»? Где проходят границы биосферы?
2. Попытайтесь кратко сформулировать сущность учения В. И. Вернадского о биосфере? Какое значение учение о биосфере имеет для понимания и решения современных экологических проблем?
3. Что называется «живым веществом», по В. И. Вернадскому? Какие вещества, кроме живого, В. И. Вернадский выделил в биосфере?
4. Назовите и раскройте основные свойства живого вещества. Сравните их с неживым веществом (косным).
5. Перечислите и раскройте содержание основных функций живого вещества. Каково их значение в процессах планетарного масштаба?
6. Сформулируйте содержание и раскройте сущность закона биогенной миграции атомов (В. И. Вернадского).
7. Назовите и раскройте содержание основных свойств биосферы. Как они связаны с живым веществом?
8. Что является основным условием устойчивости биосферы и других экологических систем?
9. Приведите примеры саморегулирующей способности биосферы.
10. Почему биосфера потенциально бессмертна с энергетической и вещественной точек зрения?
11. Что является центральным звеном биосферы?
IV. ЭКОСИСТЕМНЫЙ УРОВЕНЬ ЖИЗНИ
Экосистемы, как отмечалось в разделе («Основные понятия...»), являются основными звеньями (блоками) биосферы. Там же дается определение термина. На уровне экосистем, особенно элементарном (неделимом), представляется возможным рассмотреть более детально, глубоко и последовательно, чем это сделано на примере биосферы, основные свойства и закономерности их функционирования, важные как в теоретическом плане, так и для решения прикладных задач.
Экосистемы в конечном счете являются основными объектами (ячейками) научно обоснованного природопользования, особенно возобновимыми ресурсами. Их уровень используется для обоснования допустимых объемов изъятия продукции (при сохранении устойчивости), увеличения ее выхода (повышения продуктивности) и для решения других вопросов. В конечномсчете через сохранение отдельных элементарных экосистем решается важнейшая проблема современности - предотвращения или нейтрализации неблагоприятных явлений глобального кризиса, сохранения биосферы в целом.
IV. 1. Организация (структура) экосистем
Для того чтобы экосистемы функционировали (существовали) неограниченно долго и как единое целое, они должны обладать свойствами связывания и высвобождения энергии, а также круговоротом веществ. Экосистема, кроме этого, должна иметь механизмы, позволяющие противостоять внешним воздействиям (возмущениям, помехам), гасить их. Для раскрытия этих механизмов познакомимся с различными видами структур и другими характеристиками (свойствами) экосистем.
Блоковая модель экосистемы. Любая экосистема состоит из двух блоков. Один из них представлен комплексом взаимосвязанных живых организмов - биоценозом, а второй - факторами среды - биотопом или экотопом. В таком случае можно записать: экосистема = биоценоз + биотоп (экотоп). В. Н. Сукачев блоковую модель в ранге биогеоценоза в виде схемы изобразил на рис. 2.
Этот рисунок позволяет наглядно представить, чем отличаются понятия «экосистема» и «биогеоценоз», на что мы обращали внимание в разделе «Основные понятия...». Биогеоценоз, по В. Н. Сукачеву, включает все названные блоки и звенья. Это понятие обычно используют применительно к сухопутным системам. В биогеоценозах обязательно наличие в качестве основного звена растительного сообщества (фитоценоза). Примеры биогеоценозов - однородные участки леса, луга, степи, болота и т. п.
Экосистемы могут и не иметь растительное звено. Таким примером являются системы, формирующиеся на базе разлагающихся органических остатков, гниющих в лесу деревьев, трупов животных и т. п. В них достаточно присутствие зооценоза и микробоценоза или только микробоценоза, способных осуществлять круговорот веществ.
Таким образом, каждый биогеоценоз может быть назван экосистемой, но не каждая экосистема относится к рангу биогеоценоза.
Чтобы снять терминологические неясности, соавтор В. Н. Сукачева по формированию науки биогеоценологии - профессор В. Н. Дылис - образно определил биогеоценоз как экосистему, но только в рамках фитоценоза.
Биогеоценозы и экосистемы могут различаться и по временному фактору (продолжительности существования). Любой биогеоценоз потенциально бессмертен, поскольку все время пополняется энергией за счет деятельности растительных фото- или хемосинтезирующих организмов. В то же время экосистемы без растительного звена заканчивают свое существование одновременно с высвобождением в процессе разложения субстрата всей содержащейся в нем энергии. Надо, однако, иметь в виду, что в настоящее время термины «экосистема» и «биогеоценоз» нередко рассматриваются как синонимы.
Видовая структура экосистем. Под видовой структурой понимается количество видов, образующих экосистему, и соотношение их численностей. Точных данных о количестве видов в экосистемах нет. Это связано с тем, что трудно учесть видовое разнообразие всех мелких организмов (особенно микроорганизмов). Оно исчисляется сотнями и десятками сотен. Видовое разнообразие обычно тем значительнее, чем богаче условия (биотоп) экосистемы. В этом отношении самыми богатыми по видовому разнообразию являются, например, экосистемы дождевых тропических лесов. Только древесные виды исчисляются в них сотнями.
Рис.2
Схема биогеоценоза (экосистемы), по В.Н.Сукачеву

Богатство видов зависит также от возраста экосистем. Молодые экосистемы, возникающие, например, на таком изначально безжизненном субстрате, как отвалы пород, извлекаемые из глубинных слоев земной коры при добыче полезных ископаемых, крайне бедны видами. В дальнейшем по мере развития экосистем их видовое богатство увеличивается. Но в хорошо сформировавшихся экосистемах оно может несколько уменьшаться. К тому времени обычно выделяется один или 2-3 вида, которые явно преобладают по численности особей. Например, в еловом лесу - ель, в смешанном - ель, береза и осина, в степи - ковыль и типчак. Эти виды занимают большую часть пространства, оставляя меньше места для других видов.
Виды, явно преобладающие по численности особей, носят название доминантных (лат. доминантис - господствующий). Наряду с доминантами в экосистемах выделяются виды-эдификаторы (лат. эдификатор - строитель). К ним относят те виды, которые являются основными образователями среды. Обычно вид-доминант одновременно является и эдификатором. Например, ель в еловом лесу наряду с доминантностью обладает высокими эдификаторными свойствами. Они выражаются в ее способности сильно затенять почву, создавать кислую среду своими корневыми выделениями и при разложении мертвого органического вещества, образовывать специфические для кислой среды подзолистые почвы. Вследствие высоких эдификаторных свойств ели под ее пологом могут жить только виды растений, которые способны мириться со скудным освещением (теневыносливые и тенелюбивые). В то же время под пологом елового леса доминантным видом может быть, например, черника, но она не является существенным эдификатором.
Видовое разнообразие - очень важное свойство экосистем. С ним, как отмечалось выше, связана устойчивость систем к неблагоприятным факторам среды. Разнообразие обеспечивает как бы подстраховку, дублирование устойчивости. Вид, который присутствует в числе единичных экземпляров, при неблагоприятных условиях для широко представленного вида, в том числе и доминантного, может резко увеличить свою численность и таким образом заполнить освободившееся пространство (экологическую нишу), сохранив экосистему как единое целое.
Видовую структуру обычно используют для оценки условий местопроизрастания по растениям-индикаторам. Так, для лесной зоны кислица указывает на условия увлажнения, близкие к оптимальным, и значительное богатство почв питательными минеральными веществами; черника - на несколько избыточное увлажнение и некоторый дефицит элементов минерального питания; брусника - на дефицит увлажнения и почвенного плодородия; мхи (кукушкин лен и особенно сфагнум) - на чрезмерно избыточное увлажнение, дефицит минеральных веществ, недостаток кислорода для дыхания корней и наличие процессов торфообразования. Наряду с индикаторами меняется состав и других видов, произрастающих под пологом эдификаторов.
Названия экосистем (биогеоценозов). По растениям-эдификаторам или доминантам и растениям-индикаторам обычно называют биогеоценозы (экосистемы). Лесоводы их определяют как типы леса (например, ельники-кисличники, ельники-черничники, ельнико-сфагновые и др.). По такому же принципу классифицируются и называются другие экосистемы. Например, для степей выделяются типчаково-ковыльные, злаково-разнотравные и другие системы.
Трофическая (функциональная) структура экосистем. Цепи питания. Любая экосистема включает несколько трофических (пищевых) уровней или звеньев. Первый уровень представлен растениями. Их называют автотрофами (греч. аутос - сам; трофо - пища) или продуцентами (лат. продуцена - создающий). Второй и последующие уровни представлены животными. Их называют гетеротрофами (греч. геторос - другой) или консумен-тами. Последний уровень в основном представлен микроорганизмами и грибами, питающимися мертвым веществом. Их называют редуцентами (лат. редуцере - возвращать). Они разлагают органическое вещество до исходных минеральных элементов.
Взаимосвязанный ряд трофических уровней представляет цепь питания, или трофическую цепь (рис. 3). Главное свойство цепи питания - осуществление биологического круговорота веществ и высвобождение запасенной в органическом веществе энергии. Важно подчеркнуть, что цепь питания не всегда может быть полной. В ней могут отсутствовать растения (продуценты). Такая цепь питания характерна, как отмечалось выше, для сообществ, формирующихся на базе разложения животных или растительных остатков, например, накапливающихся в лесах на почве (лесная подстилка).
В цепи питания очень часто отсутствуют или представлены небольшим количеством животные (гетеротрофы). Например, в лесах отмирающие растения или их части (ветви, листья и др.) сразу включаются в звено редуцентов, которые завершают круговорот.
Исходя из положения: разнообразие - синоним устойчивости, можно заключить, что экосистемы с более длинными цепями питания характеризуются повышенной надежностью и более интенсивным круговоротом веществ.
IV.2. Связи организмов в экосистемах
Ни один организм в природе не существует вне связей со средой и другими организмами. Эти связи - основное условие функционирования экосистем. Через них, как было показано выше, осуществляется образование цепей питания, регулирование численности организмов и их популяций, реализация механизмов устойчивости систем и другие явления. В процессе взаимосвязей происходит поглощение и рассеивание энергии и в конечном счете осуществляются средообразующие, средоохранные и средостабилизирующие функции систем.
Подобные экосистемные связи обусловлены всем ходом эволюционного процесса. По этой причине и любое их нарушение не остается бесследным, требует длительного времени для восстановления. В связи с этим экологически обусловленное поведение человека в природе невозможно без знакомства с этими связями и последствиями их нарушения. Целесообразно выделять взаимосвязи и взаимоотношения организмов в природе (экосистемах) как различные понятия.
Взаимосвязи организмов. Взаимосвязи обычно классифицируются по «интересам», на базе которых организмы строят свои отношения.
Самый распространенный тип связей базируется на интересах питания. Такие связи носят название пищевых или трофических (греч. трофо - питание). В данный тип связей выделяется питание одного организма другим или продуктами его жизнедеятельности (например, экскрементами), питание сходной пищей (например, мертвым органическим веществом). Этим типом связей объединяются растения и насекомые, опыляющие их цветки. На базе трофических связей возникают цепи питания.
Связи, основанные на использовании местообитаний, носят название топических (греч. топос - место). Например, топические связи возникают между животными и растениями, которые предоставляют им убежище или местообитание (насекомые, прячущиеся в расщелинах коры деревьев или живущие в гнездах птиц, растения, поселяющиеся на стволах деревьев (но не паразиты). Не только трофическими, но и топическими отношениями связаны паразиты с организмами, на которых они паразитируют.
Рис.3
Трофическая (функциональная) структура экосистемы (цепь питания) и круговорот вещества в ней
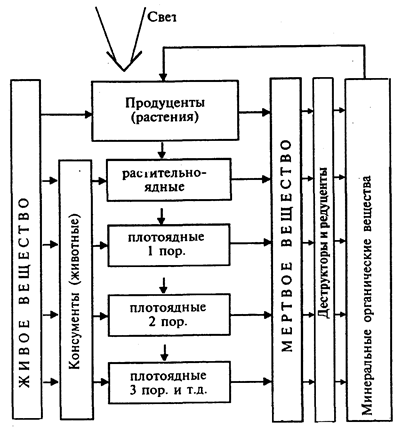
Следующий тип связей носит название форических (лат. форас -наружу, вон). Они возникают в том случае, если одни организмы участвуют в распространении других или их зачатков (семян, плодов, спор). Животными это распространение может осуществляться как на наружных покровах, так и в пищеварительном тракте.
Выделяют также тип связей, которые носят название фабрических (лат. фабрикатио - изготовление). Для них характерно использование одними организмами других или продуктов их жизнедеятельности, частей (например, растений, перьевого покрова, шерсти, пуха) для постройки гнезд, убежищ и т. п.
Взаимоотношения организмов. Данная классификация строится по принципу влияния, которое оказывают одни организмы на другие в процессе взаимных контактов. Эти взаимоотношения можно обозначить математическими значками «+», «–», «0» (положительно, отрицательно, нейтрально).
Если взаимоотношения обоим партнерам выгодны, они обозначаются значками (+,+) и носят название симбиоза или мутуализма. Степень этих связей различна. В ряде случаев организмы настолько тесно связаны, что функционируют как единый организм. Например, лишайники, представляющие симбиоз гриба и водоросли. Водоросль поставляет грибу продукты фотосинтеза, а гриб для водоросли является поставщиком минеральных веществ и, кроме того, субстратом, на котором она живет. В то же время сожительство грибов с корнями растений (микориза) носит хотя и взаимовыгодные, но не в такой степени тесные взаимоотношения. Тип взаимовыгодных отношений широко распространен. Сюда относятся и микроорганизмы, населяющие пищеварительный тракт животных, способствуя усвоению пищи; и, в ряде случаев, травоядные животные. Установлено, что исключение поедания трав животными может иметь следствием оскудение растительных сообществ, снижение ими продуктивности и устойчивости. Даже умеренное объедание листьев древесных растений насекомыми или их гусеницами может быть положительным не только для животных, но и для растений.
Взаимоотношения, которые положительны для одного вида и отрицательны для другого (+,-), характеризуются как хищничество и паразитизм. Хищник и паразит обычно приспосабливаются к использованию других организмов (их жертв и хозяев), а последние, в свою очередь, имеют адаптации, которые сохраняют им жизнь. Эти типы взаимоотношений обычно играют большую роль в регулировании численности организмов. Интенсивное размножение хищников и паразитов обычно имеет следствием уменьшение численности их жертв или хозяев.
В свою очередь, уменьшение численности жертв и хозяев подрывает кормовую базу хищников и паразитов, что ведет к сокращению их численности и т. д. В конечном счете имеет место обычно пульсирующая численность организмов, вступающих в такие типы взаимоотношений.
Хотя взаимоотношения типа хищничества и паразитизма сходны по результатам влияния на численность особей, они резко различаются по образу жизни и адаптациям. Во взаимоотношениях хищник-жертва оба организма постоянно совершенствуются: первый в плане успешности охоты, второй - в отношении самосохранения. И в том и в другом случае требуется быстрая реакция, высокая скорость передвижения, хорошее зрение, обоняние и т. п.
Во втором типе взаимоотношений у паразита адаптации идут по пути специализации структур на использование хозяина как источника пищи и «благоустроенного» местообитания. Результатом этого является упрощение многих органов (пищеварительный тракт, накожные покровы, органы передвижения, чувств и др.). Вместе с тем, поскольку жизнь паразита очень тесно связана с хозяином, он адаптирован на выживание во внешней среде после смерти хозяина. Достигается это за счет большого количества зачатков (семян, спор, цист и т. п.), обычно долго сохраняющихся в среде.
Адаптации хозяина направлены, как правило, на уменьшение вреда от паразита. Это проявляется в выработке активного иммунитета, заключении внутренних паразитов в различного вида капсулы (галлы, цецидии и т. п.).
В ряде случаев адаптации паразитов и хозяев приводят к их взаимовыгодным отношениям типа симбиоза. Есть основание полагать, что в большинстве случаев симбиоз (мутуализм) вырос из паразитизма.
Взаимоотношения, невыгодные обоим партнерам (-,-), носят название конкуренции. Последняя тем сильнее, чем ближе потребности организмов к фактору или условию, за которые они конкурируют. В этом отношении наиболее близки интересы организмов одного вида, и, следовательно, внутривидовая конкуренция рассматривается как более острая по сравнению с межвидовой. Однако данное положение противоречит тому факту, что практически все механизмы существования вида направлены на его выживание. Такое противоречие решается тем, что на внутривидовом уровне есть механизмы, которые позволяют снять остроту конкурентной борьбы, в том числе жертвуя частью особей (см. разд.V.2). Конкуренция и взаимоотношения типа хищник-жертва являются основными в совершенствовании видов, в то время как взаимоотношения типа мутуализма (симбиоза) способствуют оптимизации жизненных процессов, более полному освоению среды.
Менее распространенным типом взаимоотношений является комменсализм (франц. комменсал - сотрапезник) - отношения, положительные для одного и безразличные для другого партнера (+,0), его иногда делят на нахлебничество, когда один организм поедает остатки пищи со «стола» другого (крупного) организма (например, акулы и сопровождающие их мелкие рыбы; львы и гиены) и квартиранство, или синойкийю (греч. синойкос -сожительство), когда одни организмы используют другие как «квартиру», убежище. Например, молодь некоторых морских рыб прячется под зонтик из щупалец медуз, или некоторые насекомые живут в норах животных, гнездах птиц, используя их только для укрытия.
Не часто встречается также аменсализм (лат. аменс - безрассудный, безумный) - отрицательный для одного организма и безразличный для другого (-,0). Например, светолюбивое растение, попавшее под полог леса. Отношения, при которых организмы, занимая сходные местообитания, практически не оказывают влияния друг на друга, носят название нейтрализма (0,0). Например, белки и лоси в лесу. Сохранение разнообразия связей - важнейшее условие устойчивости экосистем.
IV.3. Экологическая ниша
Для понимания различного вида существующих связей в экосистемах и обусловленности механизмов их функционирования важно познакомиться с одним из основополагающих понятий экологии - экологической нишей.
Каждый вид или его части (популяции, группировки различного ранга) занимают определенное место в окружающей их среде. Например, определенньш вид животного не может произвольно менять пищевой рацион или время питания, место размножения, убежища и т. п. Для растений подобная обусловленность условий выражается, например, через светолюбие или тенелюбие, место в вертикальном расчленении сообщества (приуроченность к определенному ярусу), время наиболее активной вегетации. Например, под пологом леса одни растения успевают закончить основной жизненный цикл, завершающийся созреванием семян, до распускания листьев древесного полога (весенние эфемеры). В более позднее время их место занимают другие, более теневыносливые растения. Особая группа растений способна на быстрый захват свободного пространства (растения-пионеры), но отличается низкой конкурентной способностью и поэтому быстро уступает свое место другим (более конкурентоспособным) видам.
Приведенные примеры иллюстрируют экологическую нишу или отдельные ее элементы. Под экологической нишей понимают обычно место организма в природе и весь образ его жизнедеятельности, или, как говорят, жизненный статус, включающий отношение к факторам среды,видам пищи, времени и способам питания, местам размножения, укрытий и т. п. Это понятие значительно объемнее и содержательнее понятия «местообитание». Американский эколог Одум образно назвал местообитание «адресом» организма (вида), а экологическую нишу - его «профессией». На одном местообитании живет, как правило, большое количество организмов разных видов. Например, смешанный лес - это местообитание для сотен видов растений и животных, но у каждого из них своя и только одна «профессия» - экологическая ниша. Так, сходное местообитание, как отмечалось выше, в лесу занимают лось и белка. Но ниши их совершенно разные: белка живет в основном в кронах деревьев, питается семенами и плодами, там же размножается и т. п. Весь жизненный цикл лося связан с подпологовым пространством: питание зелеными растениями или их частями, размножение и укрытие в зарослях и т. п.
Если организмы занимают разные экологические ниши, они не вступают обычно в конкурентные отношения, сферы их деятельности и влияния разделены. В таком случае отношения рассматриваются как нейтральные.
Вместе с тем в каждой экосистеме имеются виды, которые претендуют на одну и ту же нишу или ее элементы (пищу, укрытия и пр.). В таком случае неизбежна конкуренция, борьба за обладание нишей. Эволюционно взаимоотношения сложились так, что виды со сходными требованиями к среде не могут длительно существовать совместно. Эта закономерность не без исключений, но она настолько объективна, что сформулирована в виде положения, которое получило название «правило конкурентного исключения». Автор этого правила эколог Г. Ф. Гаузе. Звучит оно так: если два вида со сходными требованиями к среде (питанию, поведению, местам размножения и т. п.) вступают в конкурентные отношения, то один из них должен погибнуть либо изменить свой образ жизни и занять новую экологическую нишу. Иногда, например, чтобы снять острые конкурентные отношения, одному организму (животному) достаточно изменить время питания, не меняя самого вида пищи (если конкуренция возникает на почке пищевых отношений), или найти новое местообитание (если конкуренция имеет место на почве данного фактора) и т. п.
Из других свойств экологических ниш отметим, что организм (вид) может их менять на протяжении своего жизненного цикла. Наиболее яркий пример в этом отношении - насекомые. Так, экологическая ниша личинок майского жука связана с почвой, питанием корневыми системами растений. В то же время экологическая ниша жуков связана с наземной средой, питанием зелеными частями растений.
Сообщества (биоценозы, экосистемы) формируются по принципу заполнения экологических ниш. В природном сформировавшемся сообществе обычно все ниши заняты. Именно в такие сообщества, например в долгосуществующие (коренные) леса, вероятность внедрения новых видов очень мала. В то же время следует иметь в виду, что занятость экологических ниш в определенной мере понятие относительное. Все ниши обычно освоены теми организмами, которые характерны для данного региона. Но если организм приходит извне (например, заносятся семена или другие зачатки) случайно или преднамеренно, например в результате внедрения человеком новых видов (интродукция, акклиматизация), то он может найти для себя свободную нишу в связи с тем, что на нее не было претендентов из набора существующих видов. В таком случае обычно неизбежно быстрое увеличение численности (вспышка) вида-пришельца, поскольку он находит крайне благоприятные условия (свободную нишу) и, в частности, не имеет врагов (хищников, паразитов или других организмов, которые им питаются). Такие явления не единичны. Например, размножение кроликов, завезенных в Австралию; перемещение ондатры из Азии в европейскую часть; интенсивное продвижение колорадского жука в новые районы.
С экологическими нишами в значительной мере связаны жизненные формы организмов. К последним относят группы видов, часто систематически далеко отстоящие, но выработавшие одинаковые морфологические адаптации в результате существования в сходных условиях. Например, сходством жизненных форм характеризуются дельфины (млекопитающие) и интенсивно передвигающиеся в водной среде хищные рыбы. В условиях степей сходными жизненными формами представлены тушканчики и кенгуру (прыгуны). В растительном мире отдельными жизненными формами представлены многочисленные виды деревьев, занимающие в качестве нити верхний ярус, кустарники, существующие под пологом леса, и травы - в напочвенном покрове.
IV.4. Энергетика экосистем
Живые организмы, входящие в экосистемы, для своего существования должны постоянно пополнять и расходовать энергию. Растения, как известно, способны запасать энергию в химических связях в процессе фотосинтеза или хемосинтеза. При фотосинтезе связывается только энергия с определенными длинами волн -380-710 нм. Эту энергию называют фотосинтетически активной радиацией (ФАР). Она по длинам волн близка к видимой части спектра. На эту радиацию обычно приходится около 40% общей солнечной радиации, достигающей земной поверхности. Остальная часть спектра относится либо к более короткой (ультрафиолетовой), либо к более длинной (инфракрасной) радиации. С последней обычно связан тепловой эффект.
Растения в процессе фотосинтеза связывают лишь небольшую часть солнечной радиации. Даже по отношению к фотосинтетически активной - это в среднем для земного шара менее 1%. Только наиболее продуктивные экосистемы, такие как плантации сахарного тростника, тропические леса, посевы кукурузы, в оптимальных условиях могут связывать до 3-5% ФАР. В опытах с кондиционированными условиями по всем факторам среды за короткие периоды времени удавалось достичь эффективности фотосинтеза по усвоению солнечной энергии порядка 8-10% ФАР.
Растения являются первичными поставщиками энергии для всех других организмов в цепях питания. Существуют определенные закономерности перехода энергии с одного трофического уровня на другой вместе с потребляемой пищей. Основная часть энергии, усвоенной консументом с пищей, расходуется на его жизнеобеспечение (движение, поддержание температуры тела и т. п.). Эту часть энергии рассматривают как траты на дыхание, с которым в конечном счете связаны все возможности ее высвобождения из химических связей органического вещества.
Часть энергии переходит в тело организма-потребителя вместе с увеличивающейся массой (приростом, продукцией). Некоторая доля пищи, а вместе с ней и энергия не усваиваются организмом. Они выводятся в окружающую среду вместе с продуктами жизнедеятельности (экскрементами). В последующем эта энергия высвобождается другими организмами, которые потребляют продукты выделения.
Баланс пищи и энергии для отдельного животного организма можно, таким образом, представить в виде следующего уравнения:
Эп = Эд +Эпр +Эп.в ,
где Эп - энергия потребленной пищи, Эд - энергия дыхания или обеспечения жизнедеятельности организма, включая движение, поддержание температуры тела, сердцебиение и т. п., Эпр - энергия прироста (запасенная в теле организма-потребителя), Эп.в - энергия продуктов выделения (в основном экскрементов).
Количество энергии, расходуемой организмами на различные цели, неоднозначно. В периоды интенсивной жизнедеятельности взрослого организма в теле его может совершенно не фиксироваться энергия. Наоборот, траты ее в ряде случаев превышают поступление (организм теряет вес). В то же время в периоды интенсивного роста организмов, особенно в периоды размножения (беременности), в теле фиксируется значительное количество энергии.
Выделение энергии с экскрементами у плотоядных животных (например, хищников) невелико, у травоядных оно более значительно, а гусеницы некоторых насекомых, питающиеся растениями, выделяют с экскрементами до 70% энергии. Однако при всем разнообразии расходов энергии в среднем максимальны траты на дыхание, которые в сумме с неусвоенной пищей составляют около 90% от потребленной. Поэтому переход энергии с одного трофического уровня на другой в среднем принимается близким к 10% от энергии, потребленной с пищей. Эта закономерность рассматривается обычно как «правило десяти процентов».
Данное правило надо оценивать как относительное, ориентировочное. Вместе с тем из него следует, что цепь питания имеет ограниченное количество уровней, обычно не более 4—5. Пройдя через них, практически вся энергия оказывается рассеянной.
Закономерности потока и рассеивания энергии имеют важные в практическом отношении следствия. Во-первых, с энергетической точки зрения крайне нецелесообразно потребление животной продукции, особенно с высоких уровней цепей питания. Образование этой продукции связано с большими потерями (рассеиванием) энергии. Особенно велики потери энергии при переходе с первого трофического уровня на второй, от растений к травоядным животным.
Часто в экологической литературе рассматривается в качестве примера цепь питания: люцерна-телята-мальчик. Показано, что если бы мальчик весом 48 кг питался только телятиной, то за год ему потребовалось бы для обеспечения жизнедеятельности 4,5 теленка, для питания которых, в свою очередь, необходим урожай люцерны с площади 4 га весом 8211 кг. Такова энергетическая цена животной пищи.
Во-вторых, чтобы сократить вероятность дефицита продуктов питания для интенсивно возрастающей численности населения (по закономерности, близкой к экспоненте), надо, чтобы в рационе людей больший удельный вес занимала растительная пища. Энергетически идеально - вегетарианство.
В-третьих, для увеличения КПД использования пищи при получении животноводческой продукции в условиях культурного хозяйства очень важно уменьшить основную статью нерационального расходования энергии - ее траты на дыхание. Это возможно за счет поддержания оптимального температурного режима в животноводческих помещениях, ограничения подвижности животных и, естественно, сбалансированности кормового рациона по различным элементам питания, а также применения различных биотехнических приемов (умеренные добавки стимуляторов роста, веществ, способствующих улучшению аппетита и т. п.).
Споры о допустимо возможной численности населения с точки зрения обеспечения питанием в значительной мере относительны, если они не учитывают, какой в среднем удельный вес в рационе отводится животной и растительной пище. Если исходить из рациона питания зажиточной части населения, потребляющей мяса 80-100 кг в год на одного человека, то явно невозможно обеспечение таким рационом современной численности населения Земли (около 6 млрд. человек). Если же исходить из необходимости обеспечения минимальных потребностей жизнедеятельности организма, при настоящем производстве продуктов питания возможно исключить голод и, кроме того, прокормить на 3-4 миллиарда населения больше современного. Для этого требует решения вопрос более сбалансированного распределения продуктов питания. Переход на вегетарианство и тем более расширение ассортимента растений, используемых в пищу, может обеспечить жизнедеятельность (с энергетической точки зрения) численности населения в 2-3 раза больше современной. Ясно, однако, что при этом останутся нерешенными многие медико-биологические проблемы здоровья и долголетия, а также допустимые пределы антропогенных нагрузок на экосистемы и биосферу в целом.
IV.5. Продуктивность и биомасса экосистем
Одно из важнейших свойств организмов, их популяций и экосистем в целом - способность создавать органическое вещество, которое называют продукцией. Образование продукции в единицу времени (час, сутки, год) на единице площади (метры квадратные, гектар) или объема (в водных экосистемах) характеризует продуктивность экосистем. Продукция и продуктивность могут определяться для экосистем в целом или для отдельных групп организмов (растений, животных, микроорганизмов) или видов.
Продукцию растений называют первичной, а животных - вторичной. Наряду с продукцией различают биомассу организма, групп организмов или экосистем в целом. Под ней понимают всю живую органическую массу, которая содержится в экосистеме или ее элементах вне зависимости от того, за какой период она образовалась и накопилась. Биомасса и продукция (продуктивность) обычно выражаются через абсолютно сухой вес.
Нетрудно понять, что величина биомассы экосистем или их звеньев во многом зависит не столько от их продуктивности, сколько от продолжительности жизни организмов и экосистем в целом. Например, большая биомасса характерна для лесных экосистем: в тропических лесах она достигает 800-1000 т/га, в лесах умеренной зоны -300-400 т/га, а в травянистых сообществах обычно не выходит за пределы 3-5 т/га. В то же время лесные и травянистые (например, луговые) экосистемы в сходных условиях существования по продуктивности могут мало различаться или различаются в сторону большей продуктивности как лесных, так и травянистых сообществ.
Для экосистем, представленных однолетними организмами, их годичная продуктивность и биомасса практически совпадают. Для древесных сообществ они резко различаются. Вообще соотношение биомассы и годовой продукции экосистем можно выразить формулой:

где Б - биомасса в данный момент времени, П - годовая продукция, Д - дыхание. Под последним применительно к экосистемам понимается вся сумма живого вещества, отчуждаемого на процессы разложения в результате гибели целых организмов (отпад) или их частей - сучьев, коры, листьев, наружных покровов (опад) и потребления гетеротрофами.
Экологические параметры продуктивности. Продукция и биомасса экосистем - это не только ресурс, используемый в пищу или в качестве различных видов сырья (техническое, топливо и т. п.). От этих показателей в прямой зависимости находится средообразующая и средостабилизирующая роль экосистем. Так, с продуктивностью растений и их сообществ тесно связана интенсивность поглощения углекислого газа и выделения кислорода. Для образования одной тонны растительной продукции (абсолютно сухой вес) обычно поглощается 1,5-1,8 т углекислого газа и выделяется 1,2-1,4 т кислорода. Биомасса, в том числе и мертвое органическое вещество, являются основными резервуарами концентрации углерода. На суше это практически единственный фактор вывода углекислого газа из процессов круговорота на длительное время. Часть этого органического вещества и вовсе исключается из круговорота или, как отмечал В. И. Вернадский, «уходит в геологию» (торф, уголь, нефть и т. п.).
Чаще всего в гумидных (влажных) районах фактором, прерывающим круговорот, выступает недостаток кислорода и кислая среда. Здесь основными очагами накопления органики являются болота. На дне глубоких водоемов захоронение органического вещества также обусловливается недостатком кислорода или избытком ядовитых веществ (например, сероводорода). В крайне сухих (аридных) условиях круговорот прерывается чаще всего недостатком влаги.
В связи с тем, что дождевые тропические леса характеризуются максимальной продуктивностью (до 20-25 т/га/год) и биомассой (до 700-1000 т/га), их рассматривают как основные аккумуляторы углерода и обогащения атмосферы кислородом, называя «легкими планеты». В северных лесах, как известно, продуктивность (6-10 т/га/год) и биомасса (300-400 т/га) значительно ниже. Однако на этом основании северным лесам никак нельзя отводить менее значительную роль в положительном балансе кислорода и углекислоты. Наоборот, их роль в этом отношении часто более значительна. Эти вопросы рассмотрены во второй части учебника.
Есть и другие экологические аспекты продуктивности и биомассы экосистем. В частности, чем больше биомасса, тем сильнее ее контакт с окружающей средой и тем значительнее такие средоохранные свойства, как очистка воздуха от пыли и химических агентов, регулирование влагооборотов, гашение шумовых воздействий и т. п.
Продуктивность различных экосистем биосферы. До недавнего времени принималось за аксиому, что основной объем первичной продукции образуется в морях и океанах, на долю которых приходится около 70% поверхности земного шара. Однако по последним данным, полученным в основном в результате осуществления Международной биологической программы (МБП), которая проводилась в 1964-1974 гг., было установлено, что основная масса первичной продукции образуется в экосистемах суши (около 115 млрд. тонн в год) и только около 55 млрд. тонн в год - в экосистемах океана (табл. 2). Дело в том, что внутренние воды океана, расположенные за пределами прибрежной (шельфовой) зоны, по продуктивности близки к пустыням наземных экосистем (10-120 г/м2 за год первичной продукции). Для сравнения отметим, что продуктивность лесов тайги составляет в среднем около 700-800, а влажных тропических лесов - 2000-2200 г/м2 за год.
Второй вопрос, на который важно получить ответ: какие же экосистемы в пределах океана и суши являются наиболее продуктивными?
В. И. Вернадский в свое время выделил очаги наибольшей концентрации жизни, назвав их пленками и сгущениями живого вещества. Под пленками живого вещества понимается его повышенное количество на больших пространствах. В океане обычно выделяют две пленки: поверхностную, или планктонную, и донную, или бентосную. Мощность поверхностной пленки обусловливается в основном эуфотической зоной, то есть тем слоем воды, в котором возможен фотосинтез. Она колеблется от нескольких десятков и сотен метров (в чистых водах) до нескольких сантиметров (в загрязненных водах). Донная пленка образована в основном гетеротрофными экосистемами, и поэтому ее продукция представлена вторичной, а количество ее зависит в основном от поступления органического вещества с поверхностной пленки.
В наземных экосистемах также выделяют две пленки живого вещества. Приземная, заключенная между поверхностью почвы и верхней границей растительного покрова, имеет толщину от нескольких сантиметров (пустыни, тундры, болота и др.) до нескольких десятков метров (леса). Вторая пленка - почвенная. Эта пленка наиболее насыщена жизнью. На 1 м2 почвенного слоя насчитывают миллионы насекомых, десятки и сотни дождевых червей и сотни миллионов микроорганизмов. Толщина данной пленки находится в прямой зависимости от мощности почвенного слоя и его богатства гумусом. В тундрах и пустынях это несколько сантиметров, на черноземах, особенно тучных, - до 2-3 метров.
Повышенные концентрации живого вещества в биосфере обычно приурочены к условиям так называемого «краевого эффекта», или экотонов. Такой эффект возникает на стыках сред жизни или различных экосистем. В приведенных примерах для водных экосистем поверхностная пленка - это зона контакта атмосферы и водной среды, донная - водной толщи и донных отложений, почвенная - атмосферы и литосферы.
Таблица 2
Продуктивность и биомасса экосистем материков и океанов земного шара (Уиттекер, 1980)

Примером повышенной продуктивности на стыках экосистем могут служить переходные экосистемы между лесом и полем («опушечный эффект»), а в водных средах - экосистемы, возникающие в эстуариях рек (места впадения их в моря, океаны и озера и т. п.).
Этими же закономерностями во многом обусловливаются упоминавшиеся выше локальные сгущения больших масс живого вещества (наиболее высокопродуктивные экосистемы).
Обычно в океане выделяют следующие сгущения жизни:
1. Прибрежные. Они располагаются на контакте водной и наземно-воздушной среды. Особенно высокопродуктивны экосистемы эстуариев. Протяженность этих сгущений тем значительнее, чем больше вынос реками органических и минеральных веществ с суши.
2. Коралловые рифы. Высокая продуктивность этих экосистем связана прежде всего с благоприятным температурным режимом, фильтрационным типом питания многих организмов, видовым богатством сообществ, симбиотическими связями и другими факторами. 3. Саргассовые сгущения. Создаются большими массами плавающих водорослей, чаще всего саргассовых (в Саргассовом море) и филлофорных (в Черном море). 4. Апвеллинговые. Эти сгущения приурочены к районам океана, где имеет место восходящее движение водных масс от дна к поверхности (апвеллинг). Они несут много донных органических и минеральных отложений и в результате активного перемешивания хорошо обеспечены кислородом. Эти высокопродуктивные экосистемы являются одним из основных районов промысла рыб и других морепродуктов. 5. Рифтовые глубоководные (абиссальные) сгущения. Эти экосистемы были открыты только в 70-х годах настоящего столетия. Они уникальны по своей природе: существуют на больших глубинах (2-3 тыс. метров). Первичная продукция в них образуется только в результате процессов хемосинтеза за счет высвобождения энергии из сернистых соединений, поступающих из разломов дна (рифтов). Высокая продуктивность здесь обязана прежде всего благоприятным температурным условиям, поскольку разломы одновременно являются очагами выхода из недр подогретых (термальных) вод. Это единственные экосистемы, не использующие солнечную энергию. Они живут за счет энергии недр Земли.
На суше к наиболее высокопродуктивным экосистемам (сгущениям живого вещества) относят: 1) экосистемы берегов морей и океанов в районах, хорошо обеспеченных теплом; 2) экосистемы пойм, периодически заливаемые водами рек, которые откладывают ил, а вместе с ним органические и биогенные вещества, 3) экосистемы небольших внутренних водоемов, богатые питательными веществами, а также 4) экосистемы тропических лесов. Продуктивность других экосистем видна из табл.3. Выше мы уже отмечали, что человек должен стремиться сохранить высокопродуктивные экосистемы - этот мощнейший каркас биосферы. Его разрушение связано с наиболее значительными отрицательными последствиями для всей биосферы.
Что касается вторичной (животной) продукции, то она заметно выше в океане, чем в наземных экосистемах. Это связано с тем, что на суше в звено консументов (травоядных) в среднем включается лишь около 10% первичной продукции, а в океане - до 50%. Поэтому, несмотря на более низкую первичную продуктивность океана, чем суши, по массе вторичной продукции эти экосистемы примерно равны (см. табл. 2).
В наземных экосистемах основную продукцию (до 50%) и особенно биомассу (около 90%) дают лесные экосистемы. Вместе с тем основная масса этой продукции поступает сразу в звено деструкторов и редуцентов. Для таких экосистем характерно преобладание детритных (за счет мертвого органического вещества) цепей питания. В травянистых экосистемах (луга, степи, прерии, саванны), как и в океане, значительно большая часть первичной продукции прижизненно отчуждается фитофагами (травоядными животными). Такие цепи носят название пастбищных или цепей выедання.
IV.6. Экологические пирамиды
Если количество энергии, продукции, биомасс или численности организмов на каждом трофическом уровне изображать в виде прямоугольников в одном и том же масштабе, то их распределение будет иметь вид пирамид.
Таблица 3
Продуктивность основных экосистем земного шара (по Н. Ф. Реймерсу, 1990)
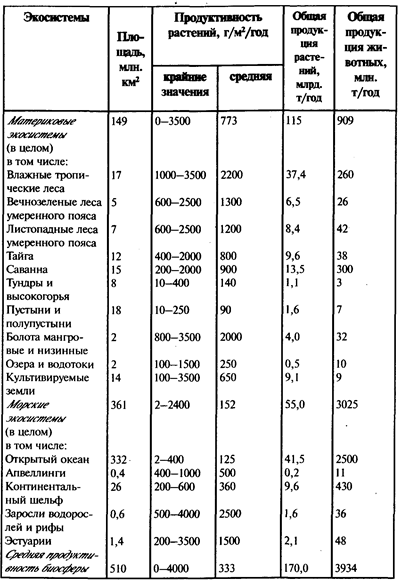
Правило пирамид энергии можно сформулировать следующим образом: количество энергии, содержащейся в организмах на любом последующем трофическом уровне цепи питания, меньше ее значений на предыдущем уровне (рис. 4а).
Рис. 4
Пирамиды энергии и продукции для экосистем суши и океана - а и биомасс для экосистем океана — б
Количество продукции, образующейся в единицу времени на разных трофических уровнях, подчиняется тому же правилу, которое характерно для энергии: на каждом последующем уровне количество продукции меньше, чем на предыдущем (рис. 4а). Более того, суммарное количество вторичной продукции (как и содержащейся в ней энергии), образующейся на разных трофических уровнях, меньше первичной продукции. Эта закономерность абсолютна и легко объясняется исходя из правила передачи энергии в цепях питания. Следует также иметь в виду, что различия в количестве энергии, содержащейся в единице веса (объема) отдельных видов продукции, невелики: 1 г (сухой вес) растительной и животной продукции содержит чаще всего от 3 до 5 калорий энергии.
Пирамиды биомасс сходны с таковыми для энергии и продукции, но только для сухопутных экосистем. Для водных экосистем закономерности соотношения биомасс на различных трофических уровнях имеют свою специфику. Здесь пирамида биомасс как бы перевернута (рис. 4б), то есть биомасса животных, потребляющих растительную продукцию, больше биомассы растительных организмов. Причина этого - резкие различия в продолжительности жизни организмов сравниваемых уровней. Первый уровень (продуценты) представлен в основном фитопланктоном с крайне коротким периодом жизни (несколько дней или часов), второй -более долгоживущими организмами - зоопланктоном или другими животными, питающимися фитопланктоном и зоопланктоном (рыбы, моллюски, киты и т. п.). Они накапливают биомассу годами и десятилетиями.
Пирамида чисел свидетельствует, что количество организмов, как правило, уменьшается от основания к вершине. Это правило не абсолютно и применимо в основном к цепям питания, не включающим редуцентов. Примером может служить пищевая цепь: насекомые и их личинки - насекомоядные животные - хищники.
IV.7. Динамика и развитие экосистем. Сукцессии
Любая экосистема, приспосабливаясь к изменениям внешней среды, находится в состоянии динамики. Эта динамика может касаться как отдельных звеньев экосистем (организмов, популяций, трофических групп), так и системы в целом. При этом динамика может быть связана, с одной стороны, с адаптациями к факторам, которые являются внешними по отношению к экосистеме, а с другой - к факторам, которые создает и изменяет сама экосистема.
Самый простой тип динамики - суточный. Он связан с изменениями в фотосинтезе и транспирации (испарении воды) растений. В еще большей мере эти изменения связаны с поведением животного населения. Одни из них более активны днем, другие - в сумерки, третьи - ночью. Аналогичные примеры можно привести по отношению к сезонным явлениям, с которыми еще больше связана активность жизнедеятельности организмов.
Не остаются неизменными экосистемы и в многолетнем ряду. Если в качестве примера взять лес или луг, то не трудно заметить, что в разные годы этим экосистемам свойственны свои особенности. В одни годы мы можем наблюдать увеличение численности одних видов (на лугах, например, бывают «клеверные годы», годы с резким увеличением злаков и других видов или групп видов). Из этого следует, что каждый вид индивидуален по своим требованиям к среде, и ее изменения для одних видов благоприятны, а на другие, наоборот, оказывают угнетающее влияние. Сказывается также и периодичность в интенсивности размножения.
Эти изменения в одних случаях могут в какой-то мере повторяться, в других же имеют однонаправленный, поступательный характер и обусловливают развитие экосистемы в определенном направлении.
Периодически повторяющуюся динамику называют циклическими изменениями или флуктуациями, а направленную динамику именуют поступательной или развитием экосистем. Для последнего вида динамики характерным является либо внедрение в экосистемы новых видов, либо смена одних видов другими. В конечном же счете происходят смены биоценозов и экосистем в целом. Этот процесс называют сукцессией (лат. сукцессио - преемственность, наследование). Различают обычно первичные и вторичные сукцессии.
Первичные сукцессии. Под первичной обычно понимается сукцессия, развитие которой начинается на изначально безжизненном субстрате. Ход первичной сукцессии рассмотрим на примере наземных экосистем. Если взять участки земной поверхности, например заброшенные песчаные карьеры, в различных географических районах (в лесной, степной зонах либо среди тропических лесов и т. п.), то для всех этих объектов будут характерны как общие, так и специфические изменения в экосистемах.
В качестве общих закономерностей будет иметь место заселение живыми организмами, увеличение их видового разнообразия, постепенное обогащение почв органическим веществом, возрастание их плодородия, усиление связей между различными видами или трофическими группами организмов, уменьшение числа свободных экологических ниш, постепенное формирование все более сложных биоценозов и экосистем, повышение их продуктивности. Более мелкие виды организмов, особенно растительных, при этом, как правило, сменяются более крупными, интенсифицируются процессы круговорота веществ и т. п. В каждом случае при этом можно выделить последовательные стадии сукцессий, под которыми понимается смена одних экосистем другими, а сукцессионные ряды заканчиваются относительно мало изменяющимися экосистемами. Их называют климаксными (греч. климакс - лестница), коренными или узловыми.
Специфические закономерности сукцессий заключаются прежде всего в том, что каждой из них, как и каждой стадии, присущ тот набор видов, которые, во-первых, характерны для данного региона, а во-вторых, наиболее приспособлены к той или иной стадии развития сукцессионного ряда. Различными будут и завершающие (климаксные) сообщества (экосистемы).
Американский эколог Клементс, наиболее полно разработавший учение о сукцессиях, считает, что в любом обширном географическом районе, который по масштабам можно примерно приравнять к природной зоне (лесная, степная, пустынная и т. п.), каждый ряд завершается одной и той же климаксной экосистемой (моноклимаксом). Такой климакс был назван климатическим. Это, однако, не значит, что для любого участка географической зоны (моноклимакса) характерен один и тот же набор видов. Видовой состав климаксных экосистем может существенно различаться. Общим является лишь то, что эти экосистемы объединяет сходство видов-эдификаторов, то есть тех, которые в наибольшей мере создают среду обитания. Например, для степных экосистем эдификаторами являются плотнокустовые злаки (ковыль и типчак). Для тропических лесов в качестве эдификаторов выступает большое количество древесных видов, создающих сильное затенение для других видов своим пологом.
Для лесной зоны северных и срединных регионов Евразии основными эдификаторами выступают ель или пихта. Из набора всех древесных видов они в наибольшей степени изменяют условия место-произрастания: сильно затеняют подпологовое пространство, создают кислую среду почв и обусловливают процессы их оподзоливания (растворение и вымывание из приповерхностного слоя практически всех минералов, кроме кварца). С этими эдификаторами уживаются только те древесные виды, которые не отстают от них в росте и способны первыми захватить пространство. При сочетании таких условий возможно формирование климаксных смешанных елово-лиственных (пихтово-лиственных), чаще всего с березой и осиной, лесов. Последнее наиболее характерно для зоны смешанных лесов. Для таежной (более северной) зоны более типичны климаксные леса с явным преобладанием только эдификаторов (ель, пихта).
Однако прежде чем сформируется климаксное сообщество (экосистема), ему предшествует, как отмечалось выше, ряд промежуточных стадий или серий. Так, в лесной зоне (рис.5) на исходно безжизненном субстрате здесь сначала появляются организмы-пионеры, например, корковые водоросли, накипные лишайники и некоторые малотребовательные к плодородию субстрата семенные растения. За ними следует стадия растительности, представленная в основном травами, а затем кустарниками и деревьями-пионерами (чаще всего березой, осиной, ивой). Последние характеризуются быстрым ростом, но, отличаясь высоким светолюбием, быстро изреживаются (к 40-50-летнему возрасту). В результате этого под их пологом создаются условия для поселения теневыносливой ели, которая постепенно догоняет в росте стареющие лиственные виды деревьев и выходит в первый ярус. На данной стадии и образуется климаксное смешанное елово-лиственное сообщество или чисто еловый лес со свойственным им набором других видов растений и животных.
Рис. 5
Биогеоценотическая сукцессия на примере смен фитоценозов в лесной зоне

Наряду с теорией моноклимакса существует точка зрения, в соответствии с которой в одном и том же географическом районе может формироваться несколько завершающих (климаксных) экосистем (поликлимакс). Например, в лесной зоне наряду с еловыми и елово-лиственными лесами в качестве климаксных рассматривают также луговые экосистемы, сосновые леса. Однако сторонники моноклимакса считают, что луга в лесной зоне могут длительно существовать только в результате их использования (скашивания, выпаса). При прекращении таких воздействий на смену им неизбежно придут лесные сообщества. Что касается сосновых лесов, то длительное существование их связывается с тем, что они занимают обычно крайне бедные (например, песчаные, щебнистые, сильно заболоченные) местообитания, где ель (более сильный эдификатор) не может внедряться и существовать вследствие более значительной требовательности к почвенному плодородию. Но с течением времени по мере накопления в почве органических веществ и необходимых для жизни минеральных элементов и эти «сосновые местообитания», с точки зрения сторонников моноклимакса, будут заняты еловыми лесами, как обладающими более сильной эдификаторной способностью.
Причины сукцессий. Сукцессионные смены обычно связывают с тем, что существующая экосистема (сообщество) создает неблагоприятные условия для наполняющих ее организмов (почвоутомление, неполный круговорот веществ, самоотравление продуктами выделений или разложения и т. п.). Такие явления реальны, но не объясняют всех случаев смен экосистем. Например, в северных лесах внедрение ели под полог лиственных древесных сообществ связано прежде всего с тем, что она использует биологические свойства этих сообществ по слабому притенению почвы. Сами же почвенные условия остаются не только благоприятными для лиственных древостоев, но и постепенно улучшаются для них (идет накопление питательных веществ, уменьшается кислотность почв и т.п.). Следовательно, здесь нет оснований говорить о самоотравлении или других подобных причинах смен.
Не подтверждается безоговорочно и точка зрения о том, что появление ели под пологом лиственных древостоев связано с тем, что в молодом возрасте она требует затенения. Известно, например, что ель и в молодом возрасте прекрасно растет при полном освещении (значительно лучше, чем под пологом других древесных видов). Об этом, в частности, свидетельствуют многочисленные примеры создания культурных фитоценозов ели (посадкой молодых растений или посевом семян) на открытых площадях.
Наряду с природными факторами, причинами динамики экосистем все чаще выступает человек. К настоящему времени им разрушено большинство коренных (климаксных) экосистем. Например, степи почти полностью распаханы (сохранились только на заповедных участках). Преобладающие площади лесов представлены переходными (временными) экосистемами из лиственных древесных пород (береза, осина, реже ива, ольха и др.). Эти леса обычно называют производными или вторичными. Они, как отмечалось выше, являются промежуточными стадиями сукцессий.
К сменам экосистем ведут также такие виды деятельности человека, как осушение болот, чрезмерные нагрузки на леса. Например, в результате отдыха населения (рекреации), химических загрязнений среды, усиленного выпаса скота, пожаров и т. п.
Антропогенные воздействия часто ведут к упрощению экосистем. Такие явления обычно называют дигрессиями (лат. дигрессион - отклонение). Различают, например, пастбищные, рекреационные и другие дигрессии. Смены такого типа обычно завершаются не климаксными экосистемами, для которых характерно усложнение структуры, а стадиями катоценоза (греч. ката - вниз, против; кайнос - общий), которые нередко заканчиваются полным распадом экосистем.
Климаксные экосистемы обычно чувствительны к различным вмешательствам в их жизнь. К подобным воздействиям, кроме хвойных лесов, чувствительны и другие коренные сообщества, например, дубовые леса. Это одна из причин катастрофической гибели дубрав в современный период и замены их, как и хвойных лесов, менее ценными, но более устойчивыми временными экосистемами из березы, осины, кустарников или трав. Последнее особенно типично при разрушении степных и лесостепных дубрав.
Кроме песчаных пространств, первичные сукцессии могут начинаться на горных породах, извлеченных из недр, продуктах извержения вулканов (застывшая лава, отложения пепла) и т. п.
Вторичные н другие сукцессии. Вторичные сукцессии отличаются от первичных тем, что они начинаются обычно не с нулевых значений, а возникают на месте нарушенных или разрушенных экосистем. Например, после вырубок лесов, лесных пожаров, при зарастании площадей, находившихся под сельскохозяйственными угодьями. Основное отличие этих сукцессий заключается в том, что они протекают несравненно быстрее первичных, так как начинаются с промежуточных стадий (трав, кустарников или древесных растений-пионеров) и на фоне более богатых почв. Конечно, вторичная сукцессия возможна только в тех случаях, если человек не будет оказывать сильное и постоянное влияние на развивающиеся экосистемы. В последнем случае, как отмечалось выше, процесс пойдет по схеме дигрессий и завершится стадией катоценоза и опустынивания территорий.
Различают также автотрофные и гетеротрофные сукцессии. Рассмотренные выше примеры сукцессий относятся к автотрофным, поскольку все они протекают в экосистемах, где центральным звеном является растительный покров. С его развитием связаны смены гетеротрофных компонентов. Такие сукцессии потенциально бессмертны, поскольку все время пополняются энергией и веществом, образующимися или фиксирующимися в организмах в процессе фотосинтеза либо хемосинтеза. Завершаются они, как отмечалось, климаксной стадией развития экосистем.
К гетеротрофным относятся те сукцессии, которые протекают в субстратах, где отсутствуют живые растения (продуценты), а участвуют лишь животные (гетеротрофы). Этот вид сукцессий имеет место только до тех пор, пока присутствует запас готового органического вещества, в котором сменяются различные виды организмов-разрушителей. По мере разрушения органического вещества и высвобождения из него энергии сукцессионный ряд заканчивается, система распадается. Таким образом, эта сукцессия по природе своей деструктивна. Примерами гетеротрофных являются сукцессии, имеющие место, например, при разложении мертвого дерева или животного. Так, при разложении мертвого дерева можно выделить несколько стадий смен гетеротрофов. Первыми на мертвом, чаще ослабленном дереве поселяются насекомые-короеды. Далее их сменяют насекомые, питающиеся древесиной (ксилофаги). К ним относятся личинки усачей, златок и др. Одновременно идут смены грибного населения. Они имеют примерно следующую последовательность: грибы-пионеры (обычно окрашивают древесину в разные цвета), грибы-деструкторы, способствующие появлению мягкой гнили, и грибы-гумификаторы, превращающие часть гнилой древесины в гумус. На всех стадиях сукцессий присутствуют также бактерии. В конечном счете органическое вещество в основной массе разлагается до конечных продуктов: минеральных веществ и углекислого газа. Гетеротрофные сукцессии широко осуществляются при разложении детрита (в лесах он представлен лесной подстилкой). Они протекают также в экскрементах животных, в загрязненных водах, в частности, интенсивно идут при биологической очистке вод с использованием активного ила, насыщенного большим количеством организмов.
Общие закономерности сукцессионного процесса. Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса.
1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы. По мере развития сукцессии эти показатели возрастают.
2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Особенно возрастает количество и роль симбиотических отношений. Полнее осваивается среда обитания, усложняются цепи и сети питания.
3. Уменьшается количество свободных экологических ниш, и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме. В связи с этим по мере развития сукцессии уменьшается вероятность вспышек численности отдельных видов.
4. Интенсифицируются процессы круговорота веществ, потока энергии и дыхания экосистем.
5. Скорость сукцессионного процесса в большей мере зависит от продолжительности жизни организмов, играющих основную роль в сложении и функционировании экосистем. В этом отношении наиболее продолжительны сукцессии в лесных экосистемах. Короче они в экосистемах, где автотрофное звено представлено травянистыми растениями, и еще быстрее протекают в водных экосистемах.
6. Неизменяемость завершающих (климаксных) стадий сукцессий относительна. Динамические процессы при этом не приостанавливаются, а лишь замедляются. Продолжаются динамические процессы, обусловливаемые изменениями среды обитания, сменой поколений организмов и другими явлениями. Относительно большой удельный вес занимают динамические процессы циклического (флуктуационного) плана.
7. В зрелой стадии климаксного сообщества (не старческой!) биомасса обычно достигает максимальных или близких к максимальным значений. Неоднозначна продуктивность отдельных сообществ на стадии климакса. Обычно считается, что по мере развития сукцессионного процесса продуктивность увеличивается и достигает максимума на промежуточных стадиях, а затем в климаксном сообществе резко уменьшается. Последнее связывают, во-первых, с тем, что в это время максимум первичной продукции потребляется консументами, а во-вторых, экосистема развивает чрезвычайно большую массу ассимиляционного аппарата, что ведет к дефициту освещенности, следствием чего является снижение интенсивности фотосинтеза при одновременном возрастании потерь продуктов ассимиляции на дыхание самих автотрофов.
Эти положения нельзя распространять на все климаксные сообщества. Например, нет реальных предпосылок для увеличения численности гетеротрофов в хвойных лесах (завершающие стадии сукцессий) по сравнению с лиственными (промежуточные стадии). Скорее, в последних больше потребителей зеленой продукции и вероятнее вспышки численности отдельных видов-фитофагов.
Нет также ни теоретических предпосылок,ни фактических данных, которые бы свидетельствовали, что в зрелой климаксной системе, например в еловых лесах, масса хвои достигает чрезмерно (!) высоких значений.
Весь опыт лесоводства свидетельствует о наиболее высокой продуктивности климаксных лесных сообществ (применительно к лесной зоне хвойных или смешанных хвойно-лиственных лесов). В противном случае, с точки зрения получения продукции (древесины), неизбежен вывод о нецелесообразности ориентации на выращивание и сохранение климаксных стадий лесов.
Применительно к другим экосистемам, например луговым, можно согласиться с тем, что возможности получения продукции на климаксной стадии уменьшаются, однако не потому, что сокращается ее нарастание (прирост, продуктивность), а по той причине, что более значительная ее часть отчуждается гетеротрофами в результате образования устойчивых цепей выедания.
Другими словами, продуктивность экосистем на климаксных стадиях сукцессий высока, как правило, максимальна вследствие более полного освоения пространства. Однако возможности снятия человеком первичной продукции лимитируются (иногда до нулевых значений) вследствие включения ее в цепи питания консументов.
IV.8. Стабильность и устойчивость экосистем
Термины «стабильность» и «устойчивость» в экологии обычно рассматриваются как синонимы, и под ними понимается способность экосистем сохранять свою структуру и функциональные свойства при воздействии внешних факторов.
Более целесообразно, однако, разграничивать эти термины, понимая под «стабильностью» данное выше определение, а под «устойчивостью» - способность экосистемы возвращаться в исходное (или близкое к нему) состояние после воздействия факторов, выводящих ее из равновесия. Кроме этого, для более полной характеристики реакции экосистем на внешние факторы целесообразно пользоваться в дополнение к названным еще двумя терминами: «упругость» и «пластичность».
Упругая система способна воспринимать значительные воздействия, не изменяя существенно своей структуры и свойств. Вместе с тем при определенных (запороговых) воздействиях такая система обычно разрушается или переходит в новое качество.
Пластичная система более чувствительна к воздействиям, но она под их влиянием как бы «прогибается» и затем относительно быстро возвращается в исходное или близкое к исходному состояние при прекращении или уменьшении силы воздействия.
Примером упругих экосистем являются климаксные (например, хвойные леса в лесной зоне, коренные тундровые сообщества, типчаково-ковыльные степи и т. п.). Пластичными экосистемами для лесной зоны являются лиственные леса как промежуточные стадии сукцессий. Они, например, выносят в несколько раз больше рекреационных (связанных с посещением населения) и других (пастьба скота, разного рода загрязнения) нагрузок, чем климаксные экосистемы, в которых эдификаторами выступают хвойные виды.
При рассмотрении стабильности и устойчивости как синонимов, обычно считается, что эти качества тем значительнее, чем разнообразнее экосистемы. Данное положение является настолько универсальным, что формулируется как закон: разнообразие - синоним устойчивости (автор Эшби). С этой точки зрения тундровые и пустынные экосистемы рассматриваются как малоустойчивые (нестабильные), а тропические леса, максимально богатые по видовому составу, - как самые устойчивые (стабильные).
Для экосистем с низкой устойчивостью характерны вспышки численности отдельных видов. Последнее связывается с тем, что в маловидовых экосистемах слабо проявляются силы, уравновешивающие численность различных видов (конкуренция, хищничество, паразитизм). Так, для тундровых экосистем типичны периодические резкие увеличения численности мелких грызунов - леммингов. В качестве результата низкой устойчивости этих экосистем рассматривается легкое разрушение их под влиянием внешних воздействий (перевыпаса, технических нагрузок и т. п.). Так, колеи, образующиеся после прохода тяжелой техники (тракторов, вездеходов), сохраняются десятилетиями.
С этих же позиций к неустойчивым и низкостабильным относят агросистемы, создаваемые человеком и представленные обычно одним преобладающим видом растений, интересующим человека. С этой же точки зрения как неустойчивые и нестабильные следует рассматривать сосновые леса на бедных песчаных или щебнистых почвах. Их древесный ярус представлен в таких условиях одним видом (сосной), беден в них и напочвенный (травяной, моховой) покров.
Однако если экосистемы, приведенные выше в качестве примеров, рассматривать с позиций названных выше различии устойчивости и стабильности, то они попадают в разные категории (табл. 4).
Устойчивость, стабильность и другие параметры экосистем зависят часто не столько от структуры самих сообществ (например, их разнообразия), сколько от биолого-экологических свойств видов-эдификаторов и доминантов, слагающих эти сообщества.
Так, высокая стабильность и значительная устойчивость, как видно из табл. 4, присущи сосновым лесам на бедных песчаных почвах, несмотря на малое видовое разнообразие этих экосистем. Это связано, во-первых, с тем, что сосна довольно пластична, и поэтому на изменение условий, например уплотнение почв, она реагирует снижением продуктивности и редко - распадом экосистемы. Однако и в последнем случае, в силу бедности субстрата питательными веществами и влагой, ее молодое поколение не встречает серьезной конкуренции со стороны других видов, и экосистема довольно быстро вновь восстанавливается в том же виде эдафического (почвенного) климакса.
Таблица 4
Характеристики устойчивости и стабильности отдельных экосистем

* по основным звеньям: фитоценозам и почвам
Иные параметры устойчивости и стабильности характерны для сосняков на богатых почвах, где они могут сменяться еловыми лесами, обладающими более сильными эдификаторными свойствами. Здесь, несмотря на значительное разнообразие (по видовому составу, ярусности, трофической структуре и т. п.), экосистемы сосновых лесов характеризуются низкой стабильностью и низкой устойчивостью. Сосна в данном случае выступает как промежуточная стадия сукцессионного ряда. Ей удается занимать и удерживать какое-то время такие местообитания только в силу каких-то необычных обстоятельств. Например, после пожаров, когда уничтожаются более сильные конкуренты (ель, лиственные древесные породы).
IV.9. Агроценозы и естественные экосистемы
Основное свойство экосистем - способность естественного развития и прежде всего самовозобновления хотя бы в течение одного-двух поколений. С этой точки зрения нет основания рассматривать агроценозы как экосистемы или одну из стадий (начальную, промежуточную) сукцессионного ряда. Агроценозы сельскохозяйственных культур, особенно однолетних, существуют только при условии постоянного вмешательства человека. При прекращении такого вмешательства обычно начинается вторичная сукцессия с той стадии, которую называют сорняками. Но эта стадия уже не имеет прямого отношения к агроценозу.
Другими словами, такой агроценоз - это совершенно чуждое естественным условиям образование (сообщество), поэтому ему не присуще ни одно из свойств экосистемы (см. табл. 4).
Несколько иные свойства характерны для агроценозов, создаваемых из долгоживущих лесных растений. Эти творения человека можно относить к экосистемам если не на протяжении всей жизни, то на определенных стадиях развития. Здесь наиболее типичны два варианта.
Первый из них относится к созданию леса искусственным путем в условиях, где сукцессии не являются четко выраженными. Например, сосняков на бедных песчаных почвах (см. выше). Здесь вмешательство человека по уходу за выращиваемыми растениями требуется только на начальных этапах жизни, когда сосенки еще настолько малы и слабы, что могут не выдержать конкуренции с травами. В дальнейшем (уже с 3-5 лет жизни) сосна начинает создавать сообщество и постепенно занимать позиции доминантного вида, формирующего свою среду. В последующем образуется сообщество со всеми критериями экосистемы. Правда, некоторые свойства такой экосистемы оказываются не вполне реализованными по сравнению с естественными сообществами. В частности, это проявляется в недостаточной жизнеспособности (устойчивости), обусловленной пониженным, по сравнению с естественными экосистемами, разнообразием. Последнее снижается в результате практически абсолютной выравненности растений по возрасту (возрастная структура как таковая отсутствует) и, в какой-то мере, выравненности почвенного фона в результате обработки почвы, предшествующей посадкам или посевам растений. Пониженная устойчивость проявляется через слабую дифференциацию деревьев по росту и, как следствие этого, ослабление, хотя и в разной степени, всех особей в период смыкания крон и острой внутривидовой конкуренции.
Кроме этого, равномерное размещение растений по площади и создание одновидовых древесных фитоценозов из хвойных видов способствует широко распространенному в настоящее время грибному заболеванию - корневой губке. Последняя имеет следствием очаговую, а порой и полную гибель растительных сообществ как систем.
Второй вариант связан с местопроизрастаниями (прежде всего почвами), характеризующимися значительным богатством питательными веществами и влагой. Здесь создание экосистем, минуя промежуточные стадии сукцессий, требует длительного вмешательства человека в их жизнь. Во всяком случае, до тех пор, пока вводимый вид (например, ель или сосна) не сформирует среду, препятствующую видам-конкурентам (осина, береза, ивы и др.) внедриться в сообщество и захватить главенствующие позиции. В большинстве же случаев победа оказывается на стороне естественных процессов развития экосистем (сукцессий). И виды, вводимые человеком, вытесняются конкурентами полностью или до такой степени, что они не способны сформировать полноценную экосистему с точки зрения целей, которые преследовал человек. Если удается сформировать желаемую экосистему (насаждение), минуя промежуточные стадии сукцессий (обязательно ценой больших затрат), то такие системы, как и в первом случае, оказываются недостаточно устойчивыми.
Исключить эти недостатки искусственных экосистем в значительной мере возможно посредством создания многовидовых сообществ, конечно, при постоянной помощи виду, в котором заинтересован человек. Иногда эти поправки удачно вносит природа через внедрение умеренного количества видов промежуточных стадий сукцессий (береза, осина и др.).
Таким образом, попытки создать сразу климаксные сообщества или их подобие, минуя промежуточные, часто обречены на неудачу по разным причинам. Это должен учитывать человек при решении конкретных хозяйственных проблем. Приведенные выше примеры свидетельствуют, насколько разнообразны взаимосвязи в экосистемах, их зависимость от биотических, абиотических и антропогенных факторов, а также насколько обязателен творческий (системный) подход в каждом конкретном случае, даже при том условии, что выявлены какие-то общие (часто основополагающие) закономерности существования экосистем. Возможности моделирования и создания человеком экосистем во многом зависят не только от биологических свойств видов (в приводимых примерах растительных), но и от условий местопроизрастания. Несомненна также относительность и необходимость конкретизации таких основополагающих экологических постулатов, как «разнообразие - синоним устойчивости», неизбежность резкого снижения продуктивности экосистем в климаксной стадии, неоднозначность терминов «устойчивость» и «стабильность» и др.
Вопросы и задания
1. Из каких основных блоков (звеньев) состоит экосистема? Вспомните определение понятия «Экосистема» из разд.1.
2. Что общего и в чем различаются понятия «Экосистема» и «Биогеоценоз»? Почему каждый биогеоценоз можно назвать экосистемой, но не каждую экосистему можно относить к разряду биогеоценоза, рассматривая последний в соответствии с определением В. Н. Сукачева?
3. Приведите примеры организмов доминантов и эдификаторов. Чем они различаются по роли в экосистемах (биогеоценозах)?
4. Перечислите связи и взаимоотношения между организмами в соответствии с существующими классификациями. Какое значение такие связи имеют для существования экосистем?
5. Что называется «экологической нишей»? Чем это понятие отличается от «местообитания»?
6. Что понимается под трофической структурой экосистем? Что называют трофическим (пищевым) звеном и трофической (пищевой) цепью?
7. Какие энергетические процессы происходят в экосистемах? По каким закономерностям энергия рассеивается и передается в цепях питания? Почему «энергетическая цена» животной пищи существенно выше «энергетической цены» растительной пищи?
8. Что называется продуктивностью и биомассой экосистем? Как связаны эти показатели с воздействием экосистем на среду?
9. Назовите наиболее продуктивные экосистемы суши и океана (пленки жизни и сгущения жизни).
10. Что называется сукцессией? Назовите виды сукцессий. Приведите примеры первичных и вторичных автотрофных и гетеротрофных сукцессий.
11. Как изменяются основные параметры и свойства экосистем в сукцессионном ряду? Перечислите основные закономерности сукцессионного процесса.
12. Чем создаваемые человеком агроценозы отличаются от естественных экосистем (по видовому богатству, устойчивости, стабильности, продуктивности)? Могут ли агроценозы существовать без постоянного вмешательства человека, вложения в них энергии?
V. ПОПУЛЯЦИОННЫЙ УРОВЕНЬ ЖИЗНИ
Если экосистемы - это реально существующие элементы (блоки) биосферы, то популяции в изолированном виде нигде не встречаются в природе. Они входят в состав экосистем и являются одним из элементов их структуры. Популяции выделяются как относительно обособленные части отдельных видов, в пределах которых более вероятны скрещивания и передача информации, чем это возможно между различными популяциями данного вида. Важнейшим фактором обособления популяций внутри вида являются различия условий местообитания. Этот же признак лежит в основе выделения экосистем (биогеоценозов). Поэтому границы популяций часто, хотя далеко не всегда, совпадают с границами экосистем.
В конечном счете экосистему можно рассматривать как сумму популяций (или их частей) разных видов, взаимосвязанных между собой и находящихся в тесном единстве с окружающей средой.
Несмотря на абстрактность вычленения популяций из общих систем (биоценозов, экосистем), им свойственен ряд специфических закономерностей функционирования, важных для существования вида и в целом экосистем различного ранга. На уровне популяций происходят основные адаптации, естественный отбор и эволюционные процессы. Разнообразие популяций внутри вида резко увеличивает его приспособительные способности, освоение среды и, в конечном счете, возможности выживания.
Проявляя заботу о сохранении вида, человек должен прежде всего думать о сохранении популяций. Для популяций различных видов существуют допустимые пределы снижения численности особей, за которыми существование популяции становится невозможным. Точных данных о критических значениях численности популяций в литературе нет. Приводимые значения разноречивы. Остается, однако, несомненным факт, что чем мельче особи, тем выше критические значения их численности. Для микроорганизмов это миллионы особей, для насекомых - десятки и сотни тысяч, а для крупных млекопитающих - несколько десятков. Численность не должна уменьшаться ниже тех пределов, за которыми резко снижается вероятность встречи половых партнеров. Критическая численность также зависит от других факторов. Например, для некоторых организмов специфичен групповой образ жизни (колонии, стаи, стада). Группы внутри популяции относительно обособлены. Могут иметь место такие случаи, когда численность популяции в целом еще достаточно велика, а численность отдельных групп уменьшена ниже критических пределов. Например, колония (группа) перуанского баклана должна иметь численность не менее 10 тыс. особей, а стадо северных оленей - 300-400 голов.
V.1. Структура популяций
Для понимания механизмов функционирования и решения вопросов использования популяций важное значение имеют сведения об их структуре.
Различают половую, возрастную, территориальную и другие виды структуры. В теоретическом и прикладном планах наиболее важны данные о возрастной структуре, под которой понимают соотношение особей (часто объединенных в группы) различных возрастов.
Обычно наибольшей жизнеспособностью отличаются популяции, в которых все возраста представлены относительно равномерно. Такие популяции называют нормальными. Если в популяции преобладают старческие особи, это однозначно свидетельствует о наличии отрицательных факторов в ее существовании, нарушающих воспроизводительные функции. Такие популяции рассматривают как регрессивные, или вымирающие. Требуются срочные меры по выявлению причин такого состояния и их исключению. Популяции, представленные в основном молодыми особями, рассматриваются как внедряющиеся, или инвазионные. Жизненность их обычно не вызывает опасений, но велика вероятность вспышек чрезмерно высокой численности особей, поскольку в таких популяциях не сформировались трофические и другие связи. Особенно опасно, если такие популяции представлены видами, которые здесь ранее отсутствовали. В таком случае популяции обычно находят и занимают свободную экологическую нишу и реализуют свой потенциал размножения, интенсивно увеличивая численность.
Если популяция находится в нормальном или близком к нормальному состоянии, человек из нее может изымать то количество особей или биомассу (последний показатель обычно используется применительно к растительным сообществам), которая прирастает за промежуток времени между изъятиями. Ясно, что изыматься должны прежде всего особи послепродуктивного возраста (окончившие размножение). Если преследуется цель получения определенного продукта, то поправки на возраст, пол или другие характеристики популяций корректируются с учетом поставленной задачи.
Эксплуатация популяций растительных сообществ, в частности, с целью получения древесного сырья обычно приурочивается к тому периоду, когда имеет место возрастное замедление накопления продукции (прироста). Этот период обычно совпадает с максимальным накоплением древесной массы на единице площади.
Количество изымаемой продукции и способ ее изъятия сообразуется с биологическими особенностями популяций. Например, у животных, ведущих групповой образ жизни, как отмечалось выше, нельзя уменьшать численность групп до такой степени, которая повлекла бы за собой потерю ими свойств оптимизации жизненных процессов.
Лесоводами применительно к решаемым задачам и сообразуясь с эколого-биологическими свойствами популяций (экосистем) разработаны различные виды рубок. Прежде всего, они делятся на две большие группы: промежуточного и главного пользования. Рубки промежуточного пользования проводятся практически во всех возрастах жизни леса. При их проведении, наряду с изъятием продукции, преследуется цель создания более благоприятных условий для жизнедеятельности и роста остающейся части древостоя. Ими же создаются условия для увеличения доли более ценных с точки зрения целей хозяйства видов (например, хвойных в смешанных хвойно-лиственных сообществах).
При рубках главного пользования убирается весь древостой, достигший возраста спелости. Этот урожай может сниматься единовременно (сплошные рубки) или в несколько приемов (постепенные, выборочные рубки). Изымается при этом такая часть древостоя, которая не нарушила бы жизнедеятельности популяций и экосистем в целом, механизмов их самоподдержания и соморегулирования (гомеостаза). Такой тип ведения хозяйства рассматривают как мягкое управление природными процессами.
С целью создания условий для появления молодого поколения леса при сплошных рубках применяются такие лесоводственные приемы, как вырубка последовательно небольшими площадями (лесосеками). В таком случае остающиеся рядом с вырубкой древостой являются источником семян, умеренного притенения появляющегося молодого поколения леса, препятствием буйному росту конкурентов из трав, кустарников и нежелательных древесных растений. Появлению молодого поколения леса способствует также оставление на вырубках отдельных, как правило, лучших деревьев, которые выполняют роль обсеменителей и носят название семенников.
Однако в обширных лесных массивах Севера и других регионов часто проводятся так называемые концентрированные рубки большими площадями без учета возможностей восстановления их молодым поколением леса. Они проводятся с использованием тяжелой техники, сопровождаются сильным разрушением и уплотнением почвенного покрова. Это, в свою очередь, ведет, как правило, к цепным реакциям природных процессов, в частности, сложившиеся круговороты воды сменяются накоплением застойных вод на поверхности почв с последующей сменой лесных экосистем болотными. В других случаях, например на песчаных почвах, следствием подобного вмешательства в экосистемы является опустынивание или полное разрушение экосистемы. Такой тип ведения хозяйства рассматривается как жесткое вмешательство в природные процессы. Оно не должно иметь места в деятельности человека.
V.2. Динамика популяций. Гомеостаз
К числу важнейших свойств популяций относится динамика свойственной им численности особей и механизмы ее регулирования. Всякое значительное отклонение численности особей в популяциях от оптимальной связано с отрицательными последствиями для ее существования. В связи с этим популяции обычно имеют адаптационные механизмы, способствующие как снижению численности, если она значительно превышает оптимальную, так и ее восстановлению, если она уменьшается ниже оптимальных значений.
Каждой популяции и виду в целом свойственен так называемый биотический потенциал, под которым понимают теоретически возможное потомство от одной пары особей при реализации способности организмов к биологически обусловленному размножению. Обычно биотический потенциал тем выше, чем ниже уровень организации организмов. Так, дрожжевые клетки, размножающиеся простым делением, при наличии условий для реализации биотического потенциала могли бы освоить все пространство земного шара за несколько часов; гриб дождевик, приносящий до 7,5 млрд. спор, уже во втором поколении освоил бы весь земной шар. Крупным организмам с низким потенциалом размножения потребовалось бы для этого несколько десятилетий или столетий.
Однако биотический потенциал реализуется организмами со значительной степенью полноты только в отдельных случаях и в течение коротких промежутков времени. Например, если быстроразмножающиеся организмы (насекомые, микроорганизмы) осваивают какой-либо субстрат или среду, где нет конкурентов. Такие условия создаются, в частности, при освоении экскрементов крупных животных насекомыми, при размножении организмов в средах, богатых питательными веществами, например в загрязненных органическими или биогенными веществами водоемах и т. п. В этом случае увеличение численности идет по j-образной кривой. Такой тип роста носит название экспоненциального. Близкий к экспоненциальному тип роста характерен в настоящее время для популяции человека. Он обусловлен прежде всего резким снижением смертности в детском возрасте. Для человека характерна кривая увеличения численности первого типа (рис. 6).
Рис. 6
Экспоненциальная (А) и логистическая (Б) кривые роста популяций

Для большинства же популяций и видов рост численности характеризуется кривой второго типа, которая отражает высокую смертность молодых особей или их зачатков (яйца, икринки, споры, семена и т. п.). При таком типе выживаемости (смертности) численность популяции обычно выражается s-образнои кривой (см. рис.6). Такая кривая носит название логистической. Но и в этом случае периодические колебания численности особей значительны. Такие отклонения от средней численности имеют сезонный (как у многих насекомых), взрывной (как у некоторых грызунов - лемминги, белки) или сглаженный (как у крупных млекопитающих) характер. Численность при этом может существенно отличаться от средних значений: для насекомых - в 107- 106 раз, для позвоночных животных, например грызунов, - в 105 - 106 раз.
Периоды резкого изменения численности носят название «популяционных волн», «волн жизни», «волн численности». Причины таких колебаний до конца не ясны. В одних случаях их связывают с пищевым фактором, в других - с погодными (климатическими) явлениями (например, для леммингов - с количеством тепла, приносимого Гольфстримом), в третьих - с солнечной активностью или комплексом взаимосвязанных факторов, что наиболее вероятно.
Резкие изменения численности относительно средних значений имеют обычно отрицательные следствия для жизни популяций: при высокой численности - из-за ослабления всех особей в результате недостатка пищи, самоотравления среды, возможных массовых заболеваний и т. п.; при низкой численности - из-за превышения порога ее минимальных значений.
В целом можно выделить динамику популяций, независимую от плотности (численности) ее особей и зависимую от плотности. Для первого типа характерна отмеченная выше экспоненциальная кривая роста. Для второго - логистическая кривая. Соответственно различаются и механизмы (факторы), оказывающие влияние на плотность (численность) особей. При независимом от плотности типе изменения численности последняя обусловливается в основном абиотическими факторами (погодные явления, наличие пищи, различного рода катастрофы и т. п.). Эти факторы могут обеспечивать условия как для неограниченного, хотя и кратковременного роста популяций, так и для снижения их численности до нулевой. Такие факторы обычно называют модифицирующими (лат. модификацио-изменение).
Зависимая от плотности динамика популяций обеспечивается биотическими факторами. Их называют регулирующими. Они «работают» по принципу обратной отрицательной связи: чем значительнее численность. Тем сильнее срабатывают механизмы, обусловливающие ее снижение, и наоборот - при низкой численности сила этих механизмов ослабевает и создаются условия для более полной реализации биотического потенциала. Факторы такого типа лежат в основе популяционного гомеостаза, обеспечивающего поддержание численности в определенных границах значений.
К числу регулирующих факторов относится, в частности, взаимоотношение организмов типа хищник-жертва. Высокая численность жертвы создает условия (пищевые) для размножения хищника. Последний, в свою очередь, увеличив численность, снижает количество жертвы. Численность обоих видов в результате этого носит синхронно-колебательный характер. Регулирующие факторы, в отличие от модифицирующих, никогда не доводят численность популяций до нулевых значений вследствие того, что сила их действия уменьшается по мере уменьшения численности популяций.
Вообще действие регулирующих факторов можно рассматривать на уровне межвидовых и внутривидовых (внутрипопуляционных) взаимоотношений организмов.
К межвидовым механизмам гомеостаза относятся отмеченные выше взаимоотношения типа хищник-жертва. В таком же плане действуют и взаимоотношения паразит- хозяин. При высокой численности создаются условия для увеличения количества паразитов и паразитарных заболеваний как в результате скученности, так и вследствие ослабления организмов. К межвидовым механизмам относится также конкуренция, острота которой находится в прямой зависимости от численности организмов.
Конкуренция лежит и в основе внутрипопуляционного гомеостаза. Она здесь может проявляться в жестких и смягченных формах. Жесткие формы обычно заканчиваются гибелью части особей. В растительном мире это проявляется в явлениях так называемого самоизреживания фитоценозов. Например, на стадии всходов и молодых растений в лесных сообществах на одном гектаре насчитывается до нескольких сотен тысяч древесных растений. К возрасту спелости (100-120 лет для хвойных видов и 50-70 лет для лиственных) число экземпляров обычно не превышает 1000 на 1 га, но чаще исчисляется несколькими сотнями. Остальные погибают в процессе острой конкурентной борьбы (рис. 7). В результате этого, с одной стороны, освобождается пространство для остающихся более сильных особей, а с другой - ослабленные и погибающие особи выполняют положительную роль для сохранения популяции через включение в процессы круговорота, обогащение почвы минеральными веществами и гумусом. Часть ослабленных особей еще при жизни становится донорами для питания более сильных экземпляров. Это возможно в результате имеющего место срастания корней. Частным подтверждением таких явлений служат не единичные случаи нарастания годичных колец на пнях деревьев («растущие пни»).
Рис. 7
Ход самоизреживания соснового леса с возрастом (по Г. Ф. Морозову).
В животном мире результат острой внутривидовой борьбы проявляется часто в форме каннибализма (поедания себе подобных). Такие явления наиболее часты среди хищников. Например, взрослые окуни при высокой численности популяции, особенно в небольших водоемах, начинают питаться мальками своего вида. Явления каннибализма характерны также для некоторых грызунов, личинок насекомых, особенно в случаях существования в ограниченном пространстве. Поедание потомства домашними животными, по-видимому, один из случаев атавизма (лат. атавус - отдаленный предок) данного явления, которое раньше имело место в природных популяциях.
Смягченные формы внутривидовой конкуренции проявляются обычно через ослабление части особей, выключения их из процессов размножения. Случаи гибели особей при таких формах борьбы менее вероятны. К таким механизмам внутрипопуляционного гомеостаза относятся угнетающие (ингибирующие) выделения веществ во внешнюю среду более сильными особями, стрессовые явления, разграничение территорий (территориальность), миграции между популяциями.
Выделения во внешнюю среду характерны как для растительных, так и для животных организмов. Показано, в частности, что молодое поколение леса не появляется под материнским пологом либо находится в сильно угнетенном состоянии не только в результате дефицита светового и других факторов (минеральное питание, влагообеспеченность и т. п.). Опытным путем было установлено, что в этих процессах существенную роль играют ингибирующие выделения корней, а в ряде случаев атмосферные осадки, обогащенные химическими элементами и соединениями, вымывающимися из надземных органов взрослых растений. Препятствием для молодого поколения может также являться мощный слой слаборазложившегося мертвого органического вещества (лесной подстилки), препятствующий прорастанию семян и укоренению всходов. В спелых лесах сила влияния этих факторов обычно уменьшается, и молодое поколение постепенно сменяет теряющих конкурентную способность особей.
На примере лабораторных животных (крысах, мышах) показано, что воздух, подаваемый из помещений, где имеет место перенаселенность, в помещения, где животные свободно размещаются и нормально развиваются, приводит к замедлению роста и угнетению последних. Аналогичные результаты наблюдались в опытах с головастиками лягушек, когда в аквариумы, где они хорошо развивались, добавляли воду из аквариумов, в которых плотность организмов была чрезвычайно высокой.
Явления территориальности наиболее четко выражены в животном мире. Сюда относятся различные способы охраны занимаемых территорий. Например, пение птиц - это прежде всего сигнал о занятости территории в период размножения и последующего выкармливания потомства. У кошачьих и собачьих территориальность проявляется через мечение границ участков выделением желез, мочой или механическими отметинами на деревьях, почве и т. п.
При высокой скученности особей в популяциях регулирующим механизмом численности могут являться стрессовые явления. Они наиболее характерны для млекопитающих. При стрессах обычно часть особей снижает или теряет репродукционные функции (выключается из процессов размножения). Более сильные особи в меньшей степени подвержены стрессу и его следствиям. При ослаблении или прекращении стрессовых явлений организмы обычно восстанавливают функции жизнедеятельности и репродукционного процесса.
Миграции как фактор гомеостаза проявляются чаще всего в двух видах. Первый из них относится к массовому исходу особей из популяции при явлениях перенаселенности (нашествия). Такие явления особенно характерны для леммингов, белок и некоторых других видов с взрывным типом динамики численности. Особи, оставившие популяцию (как правило, молодняк), обычно не возвращаются на прежнее место. Значительное количество их погибает при подобных перемещениях.
Второй вид миграций связан с более постепенным (спокойным) уходом части особей в другие популяции с меньшей плотностью населения.
Вообще и при численности, близкой к оптимальной, популяции обмениваются, хотя и в меньшей степени, особями. Это важно как для исключения или уменьшения вероятности близкородственного скрещивания, так и для обмена информацией, которая имеет свою специфику в разных популяциях.
Неоднозначны реакции популяций на иммигрантов. В периоды высокой численности они препятствуют вселению особей из других популяций. При низкой численности имеет место явление противоположного порядка: резко уменьшается количество особей, оставляющих популяции, снимаются механизмы, препятствующие вселению особей из других популяций (иммигрантов).
Среди насекомых выселительная способность связана часто с появлением специфических особей, выделяемых обычно в отдельную фазу, характеризующихся большей подвижностью, стремлением к перемещениям. У тлей, например, появляется фаза с хорошо развитыми крыльями, у пустынной саранчи, кроме лучшего развития летательного аппарата, подвижность увеличивается за счет более темной окраски и в связи с этим лучшей прогреваемости тела, что для холоднокровных организмов является важнейшим фактором усиления активности.
Ясно, что гомеостаз в полной мере проявляется, если срабатывают все механизмы, лежащие в его основе. Например, не нарушается резко соотношение численностей хищников и жертв, не имеет места действие факторов, ослабляющих популяции (загрязнение, нарушение местообитаний и др.), не превышаются критические пределы численности и т. п.
В настоящее время подобные нарушения гомеостатических механизмов вызываются в большинстве случаев антропогенными факторами. В связи с этим одной из важнейших задач человека является исключение или резкое снижение действия подобных факторов. Решение вопросов такого плана относится прежде всего к области прикладной экологии, знакомству с которой посвящен следующий раздел настоящего пособия.
Вопросы и задания
1. Вспомните и воспроизведите определение популяции. Какие основные критерии используются при расчленении вида на популяции?
2. Назовите основные виды структуры популяций. Покажите прикладное значение возрастной структуры популяций.
3. Что понимается под биотическим потенциалом популяции (вида)? Почему он не реализуется полностью в природных условиях? Какие факторы препятствуют реализации потенциала? Нарисуйте теоретически возможную и реальную кривую роста численности особей в популяциях. Как называются эти кривые?
4. Назовите механизмы, за счет которых регулируется численность особей в популяциях. Перечислите механизмы межвидового и внутрипопуляционного регулирования численности особей в популяциях.
5. Применим ли к популяциям термин «гомеостаз» и в чем он проявляется?
Рекомендуемая литература
1. Вернадский В.И. Биосфера. - М.,1975.
2. Вернадский В.И. Живое вещество. - М., 1978.
3. Лапо А.В. Следы былых биосфер. - М., 1987.
4. Одум Ю. Экология. - Т. 1 и 2. - М.,1986.
5. Пономарева И.Н. Общая экология. - М., 1994.
6. Реймерс Н.Ф. Экология. - М., 1994;
7. Уиттекер Н.М. Сообщества и экосистемы. - М., 1980.
8. Чернова Н.М., Былова А.М. Экология. - М., 1988.
9. Шилов И.А. Экология. - М., 1997.
СОДЕРЖАНИЕ
СЛОВО К ЧИТАТЕЛЮ! (вместо предисловия)....................................................................................................................... 1
ВВЕДЕНИЕ....................................................................................................................................................................................... 2
I. ОСНОВНЫЕ ПОНЯТИЯ (ТЕРМИНЫ) ЭКОЛОГИИ. ЕЕ СИСТЕМНОСТЬ................................................................ 6
I.1. Основные понятия............................................................................................................................................................... 6
I.2. Структура общей экологии.............................................................................................................................................. 9
II. СРЕДА ОБИТАНИЯ. ФАКТОРЫ СРЕДЫ И АДАПТАЦИИ К НИМ ОРГАНИЗМОВ. СРЕДЫ ЖИЗНИ.......... 10
II.1. Среда и факторы среды, их классификация............................................................................................................ 10
II.2. Некоторые общие закономерности действия факторов среды на организмы............................................... 12
II.3. Среды жизни и адаптации к ним организмов.......................................................................................................... 14
III. БИОСФЕРА.............................................................................................................................................................................. 18
III.1. Биосфера как глобальная экосистема..................................................................................................................... 18
III.2. Живое вещество, его средообразующие свойства и функции в биосфере.................................................... 19
III.3. Основные свойства биосферы.................................................................................................................................... 22
IV. ЭКОСИСТЕМНЫЙ УРОВЕНЬ ЖИЗНИ.......................................................................................................................... 25
IV. 1. Организация (структура) экосистем........................................................................................................................ 25
IV.2. Связи организмов в экосистемах............................................................................................................................... 28
IV.3. Экологическая ниша..................................................................................................................................................... 31
IV.4. Энергетика экосистем................................................................................................................................................... 32
IV.5. Продуктивность и биомасса экосистем................................................................................................................... 34
IV.6. Экологические пирамиды............................................................................................................................................ 37
IV.7. Динамика и развитие экосистем. Сукцессии.......................................................................................................... 39
IV.8. Стабильность и устойчивость экосистем............................................................................................................... 44
IV.9. Агроценозы и естественные экосистемы................................................................................................................. 45
V. ПОПУЛЯЦИОННЫЙ УРОВЕНЬ ЖИЗНИ......................................................................................................................... 47
V.1. Структура популяций.................................................................................................................................................... 48
V.2. Динамика популяций. Гомеостаз................................................................................................................................ 49
Рекомендуемая литература....................................................................................................................................................... 54
Воронков Н.А.
Основы общей экологии
(общеобразовательный курс)
Директор В.В.Миненков
Редактор О.А.Блинникова
Корректор Н.В.Миненкова
Оформление обложки Н.Ю.Медведева
Подписано в печать 02.11.99. Формат 69х90/16.
Бумага газетная. Гарнитура «Таймс». Печать офсетная.
Усл. печ. л. 6. Тираж 10000 экз. Заказ № 2849.
Издательство «Агар»
103045, Москва, Костянский пер., д.6
Тел./факс (095) 956-76-15
232-29-90
Лицензия № ЛР №064411 от 22.01.96
Издательство "Равдеву-АМ"
129272, Москва, Олимпийский проспект, 30
Тел. (095) 976-24-44
Факс 900-70-98
Лицензия ЛР № 065494 от 31.10.1997
По вопросу приобретения обращаться в ООО ЦГЛ «РОН»
103 045, а/я 44, Москва, Костянский пер. д.6
(095) 232-29-90,956-76-15
E-mail: ron @ comail.ru
Отпечатано с готовых диапозитивов
в Государственном ордена Октябрьской Революции,
ордена Трудового Красного Знамени Московском
предприятии «Первая Образцовая типография»
Государственного комитета Российской Федерации
по печати. 113054, Москва, Валовая, 28
– Конец работы –
Используемые теги: основы, общей, экологии0.064
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Основы общей экологии
Что будем делать с полученным материалом:
Если этот материал оказался полезным для Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.192 сек.
Новости и инфо для студентов