Механизм образования временной связи условного рефлекса
Физиологической основой условного рефлекса является процесс замыкания временной связи, проторение новых путей в наиболее реактивных образованиях центральной нервной системы – в высших ее отделах.
Временная связь –это совокупность нейрофизиологических, биохимических и структурных изменений мозга, возникающих в процессе сочетания условного и безусловного раздражителей и формирующих определенные взаимоотношения между различными мозговыми центрами.
В настоящее время нет единой точки зрения на механизмы замыкания временной связи. И.П. Павлов считал, что замыкание временных связей происходит в горизонтальной плоскости на уровне коры больших полушарий между тем нервным центром, который воспринимает условный раздражитель, и корковым представительством безусловного рефлекса. Сигнальный и подкрепляющий раздражители вызывают собственные безусловные реакции: условный сигнал – ориентировочный рефлекс, а подкрепляющий раздражитель – соответствующий безусловный рефлекс. Именно эти рефлексы составляют нейросубстрат условного рефлекса. При многократном сочетании условного и безусловного раздражителей ориентировочная реакция на сигнал угасает.
В процессе формирования условного рефлекса в корковых проекциях сигнального и подкрепляющего раздражителей происходит функциональная перестройка. В результате этого повышается порог собственной безусловно-рефлек-
торной реакции на сигнал, и он начинает вызывать не свойственную ему ранее условную реакцию.
И.П. Павлов сделал вывод, что формирование временных связей является функцией именно коры больших полушарий, так как животные, лишенные проекционных зон коры, вырабатывают только наиболее элементарные, нестойкие условные рефлексы, требующие длительной выработки.
Современные электрофизиологические исследования показали, что замыкание временной связи межу различными корковыми участками происходит не в горизонтальной плоскости, как предполагал Павлов, а в вертикальной, через подкорковые структуры. Установлено, что в условно-рефлекторных реакциях принимают участие корковые отделы, ретикулярная формация и лимбическая система. Если у животных удалены структуры старой и древней коры, то выработка условных рефлексов становится невозможной. Эти факты позволили П.К. Анохину высказать предположение о вертикальном корково-подкорковом характере замыкания временной связи и сформулировать гипотезу конвергентного замыкания условного рефлекса (рис.2).
Кора больших полушарий
А Б
| ||||||||||||||
  
|







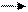 УС рецепторы рецепторы БП
УС рецепторы рецепторы БП
Рис. 2 Схема конвергентного замыкания условного рефлекса



 ЦС – нервный центр условного сигнала; ЦП – нервный центр безусловного подкрепления; – специфические подкорковые центры (таламус); – ретикулярная формация; УС – условный сигнал; БП – безусловное подкрепление;
ЦС – нервный центр условного сигнала; ЦП – нервный центр безусловного подкрепления; – специфические подкорковые центры (таламус); – ретикулярная формация; УС – условный сигнал; БП – безусловное подкрепление;
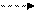

 – нейроны коры больших полушарий, на которых происходит замыкание влияний от условного и безусловного раздражителей; А – специфический путь проведения возбуждения от рецепторов, воспринимающих сигнал, в корковый центр сигнала; Б – специфический путь проведения возбуждения от рецепторов, воспринимающих безусловное подкрепление, в корковый центр безусловных рефлексов; – неспецифический путь проведения возбуждения, вызванного условным сигналом, от ядер ретикулярной формации в кору больших полушарий; – неспецифический путь проведения возбуждения, вызванного безусловным подкреплением, от ядер ретикулярной формации в кору больших полушарий.
– нейроны коры больших полушарий, на которых происходит замыкание влияний от условного и безусловного раздражителей; А – специфический путь проведения возбуждения от рецепторов, воспринимающих сигнал, в корковый центр сигнала; Б – специфический путь проведения возбуждения от рецепторов, воспринимающих безусловное подкрепление, в корковый центр безусловных рефлексов; – неспецифический путь проведения возбуждения, вызванного условным сигналом, от ядер ретикулярной формации в кору больших полушарий; – неспецифический путь проведения возбуждения, вызванного безусловным подкреплением, от ядер ретикулярной формации в кору больших полушарий.
В ее основе лежит способность нейронов отвечать на раздражения разных модальностей, т.е. явление полисенсорной конвергенции. По представлениям П.К. Анохина, условный и безусловный раздражители вызывают распространенную активацию корковых нейронов благодаря включению ретикулярной формации, которая оказывает неспецифическое активирующее влияние на кору. В результате восходящие сигналы (условного и безусловного раздражителей) перекрываются, т.е. происходит встреча (конвергенция) этих возбуждений на одних и тех же корковых нейронах. Условное и безусловное возбуждение, доходя до нейронов, фиксируется в них в виде прочных химических соединений, образование которых и представляет собой механизм замыкания условно рефлекторной связи. В результате конвергенции возбуждений возникают и стабилизируются временные связи между корковыми представительствами условного и безусловного раздражителей.
Импульсы от рецепторов, воспринимающих условный и безусловный раздражители, через специфические подкорковые образования поступают в кору больших полушарий, вызывая в ней формирование двух возбужденных очагов: центра сигнала и центра подкрепления. В то же время возбуждение из подкорковых ядер передается к нейронам ретикулярной формации, которая оказывает восходящее неспецифическое генерализованное влияние на кору. В результате происходит конвергенция условного и безусловного раздражений в одних и тех же нейронах и синхронизация активности пространственно удаленных участков головного мозга, включенных в системную организацию данного условного рефлекса. Общей закономерностью высшей нервной деятельности является образование условных рефлексов с прямыми и обратными связями. Если применять попарное сочетание между собой двух раздражителей, каждый из которых вызывает типичный безусловный рефлекс (например, мигательный или сосудисто-двигательный и пищевой), то вырабатывается условный рефлекс с двусторонней временной связью. При этом каждый из пары сочетаемых раздражителей становится условным сигналом для рефлекса партнерного раздражителя, т.е. вызывает его эффекторное выражение.
В любом условном рефлексе проявляется тенденция к анализу и синтезу. В результате условный рефлекс непрерывно совершенствуется. Возникая как приблизительный и расплывчатый, он постепенно становится точным и определенным. Такое совершенствование условного рефлекса связано с преобладанием сначала его генерализации, а затем специализации. На первых этапах образования условных рефлексов животное отвечает сходными реакциями на сигналы, похожие на условный раздражитель. Так, если условный пищевой рефлекс выработан на ноту ДО, то сначала любая другая нота вызывает этот же рефлекс. Эта стадия, получившая название стадии генерализации условного рефлекса, отражает синтетические процессы в коре больших полушарий. Но, поскольку едой подкрепляется только нота ДО, нес другие неподкрепляемые ноты перестают вызывать условное слюноотделение. Эта стадия получила название стадии специализации условного рефлекса, в основе которой лежат аналитические процессы в коре, обеспечивающие распознавание звуков.
Стадия генерализации характеризуется тем, что все раздражители, близкие по физическим параметрам к условному, приводят к проявлению условного рефлекса; это происходит благодаря свойству иррадиации возбудительного процесса. Физиологический механизм стадии генерализации условного рефлекса заключается в образовании побочных временных связей подкрепляющей реакции с пунктами сигналов, близких к основному.
С явлением генерализации связаны некоторые процессы обобщения в мыслительной деятельности человека. В двигательных условных рефлексах, например при образовании двигательных навыков, феномен генерализации проявляется в участии в двигательном акте значительного числа мышц, сокращение которых не является необходимым.
Стадия специализации характеризуется тем, что в процессе выработки условного рефлекса различные раздражители, кроме условного, становятся неэффективными и не вызывают условного рефлекса. В основе лежит процесс концентрации возбуждения.
При специализации условных рефлексов основной сигнал подкрепляется, а смежные не подкрепляются, т.е. возникают условия для развития дифференцировочного торможения. В результате основная временная связь усиливается, а побочные ослабевают. Следовательно, физиологический механизм стадии специализации условного рефлекса заключается в угашении побочных временных связей на все посторонние раздражители благодаря развитию дифференцировочного торможения. Условное возбуждение на основной сигнал концентрируется точно в его анализаторном пункте.
При образовании условных рефлексов у человека явления генерализации выражены несравненно меньше, чем у животных. Процессы концентрации, проявляющиеся у человека в значительно большей степени, чем у животных, дают возможность сразу же устранить условные рефлексы на воздействия, существенно отличающиеся от раздражителя, на который вырабатывается данный условный рефлекс.
По мере выработки условного рефлекса генерализация активности многих нейронов коры, а также подкорковых структур уменьшается. На конечной стадии формирования стабильного условного рефлекса в этот процесс втягиваются только те нейроны, которые участвуют в осуществлении условно-рефлекторной реакции. В структуру временной связи на корковом уровне будут включены кроме нейронов центра сигнала и центра подкрепления те нейроны, на которых произошла конвергенция условного и безусловного раздражений, переданных через ретикулярную формацию.
По данным электрофизиологических исследований, в процессе образования временной связи непрерывно идущие колебания потенциалов мозга (его фоновая активность) меняют свой характер. В начале выработки условного рефлекса происходит десинхронизация (активация) основного ритма. На стадии генерализации условного рефлекса возникает реакция десинхронизации, которая охватывает обширную территорию коры, включая области проекций и условного, и безусловного раздражителей. При образовании временной связи реакция синхронизации ритмов вначале формируется в корковых проекциях сигнала, затем в зоне подкрепления, а при дальнейшем упрочнении условного рефлекса она идет на убыль в зоне проекции сигнала, но долго сохраняется в зоне проекции подкрепления. При этом усвоение ритма происходит сначала в верхних, а потом в нижних слоях коры. Показатели синхронизации демонстрируют иррадиацию нервных процессов при начальной генерализации условного рефлекса, а по мере его специализации – нарастающую их концентрацию.
Образование условных рефлексов отражается в электрической активности не только коры, но и подкорковых структур. При одновременной регистрации биопотенциалов корковых областей, ядер таламуса и ретикулярной формации отмечалось, что реакции десинхронизации и синхронизации появляются в подкорке (ретикулярной формации) даже раньше, чем в коре.