КОЛЕБАНИЯ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Когда популяция прекращает расти, ее плотность обнаруживает тенденцию к флуктуациям относительно верхнего асимптотического уровня роста. Такие флуктуации могут возникать либо в результате изменений физической среды, вследствие чего повышается или снижается верхний предел численности, либо в результате внутри-популяционных взаимодействий, либо, наконец, в результате взаимодействия с соседними популяциями. После того, как верхний предел численности популяции (К) окажется достигнутым, плотность может некоторое время оставаться на этом уровне или сразу резко упасть (рис. 7.7, кривая 1). Это падение окажется еще резче, если сопротивление среды увеличивается не постепенно, по мере роста популяции, а проявляется внезапно (кривая 2). В таком случае популяция будет реализовы-
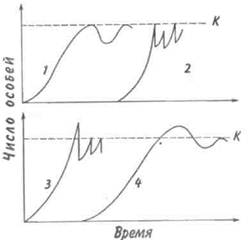
Рис. 7.7. Различные типы флуктуации плотности популяции. Объяснения в тексте.
вать биотический потенциал. Однако экспоненциальный рост не может происходить долго. Когда экспонента достигает парадоксальной точки стремления к бесконечности, как правило, происходит качественный скачок — быстрое увеличение численности сменяется массовым отмиранием клеток или гибелью особей. Пример подобных флуктуации — вспышка размножения насекомых, сменяемая массовой их гибелью; размножение и гибель водорослей («цветение» водоемов). Возможна и такая ситуация, при которой численность популяции «перескакивает» через предельный уровень (кривые 3, 4), если питательные вещества и другие необходимые для жизни факторы накоплены еще до начала роста популяции. Этим, в частности, можно объяснить, почему новые пруды и озера часто богаче рыбой, чем старые.
Существует два основных типа колебаний численности популяций (рис. 7.8). При первом типе периодические нарушения среды, такие как пожары, наводнения, ураганы и засухи, часто приводят к катастрофической, не зависящей от плотности, смертности. Так, численность популяции однолетних растений и насекомых обычно быстро растет весной и летом, а с наступлением холодной погоды резко сокращается. Популяции, рост которых дает регулярные или случайные всплески, называ-
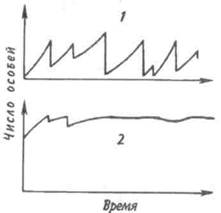
Рис. 7.8. Флуктуация плотности популяции у оппортунистических (1) и равновесных (2) видов.

 ются оппортунистическими (рис. 7.8,1). Другие популяции, так называемые равновесные (свойственные многим позвоночным), обычно находятся в состоянии, близком к состоянию равновесия с ресурсами, а значения их плотности гораздо более устойчивы (рис. 7.8,2).
ются оппортунистическими (рис. 7.8,1). Другие популяции, так называемые равновесные (свойственные многим позвоночным), обычно находятся в состоянии, близком к состоянию равновесия с ресурсами, а значения их плотности гораздо более устойчивы (рис. 7.8,2).
Два выделенных типа популяций представляют собой только крайние точки континуума, однако при сравнении разных популяций подобное разделение часто оказывается полезным. Значение противопоставления оппортунистических популяций равновесным заключается в том, что действующие на них независящие и зависящие от плотности факторы, так же как совершающиеся при этом события, по-разному влияют на естественный отбор и на сами популяции. Мак-Артур и Уилсон (1967) назвали эти противоположные типы отбора г-отбором и К-отбором в соответствии с двумя параметрами логистического уравнения. Некоторые характерные признаки г- и К-отбора приведены в таблице.
Во временном отношении колебания численности популяции бывают непериодическими и периодическими. Последние можно разделить на колебания с периодом в несколько лет и сезонные колебания. Непериодические флуктуации носят непредвиденный характер.
В Тихом океане, особенно в районе Большого барьерного рифа к северо-востоку от Австралии, с 1966 г. наблюдается увеличение численности морской звезды Терновый венец, Acanthaster planci. Терновый венец, будучи ранее малочисленным (менее одной особи на 1 м2), достиг к началу 1970-х годов плотности 1 особь на 1 м2. Морская звезда приносит большой вред коралловым рифам, так как питается полипами, составляющими их живую часть. Она «очистила» 40-километровую полосу рифов у острова Гуам менее чем за три года. Ни одна из гипотез, предложенных для объяснения внезапного увеличения численности морской звезды (исчезновение одного из ее врагов — брюхоногого моллюска Тритоний рог, Charonia tritonis, которого добывают из-за раковин, содержащих перламутр; увеличение содержания в морской воде ДДТ и в связи с этим нарушение естественного равновесия; влияние радиоактивных осадков), не может считаться удовлетворительной.
Пример периодических колебаний численности с периодом в несколько лет дают популяции некоторых арктических млекопитающих и птиц. У зайца-беляка и рыси период колебаний численности равен 9,6 года (рис. 7.9). Как видно из рисунка, максимум численности зайца по
Некоторые характерные признаки г- и /К-отбора (Пианка, 1981)

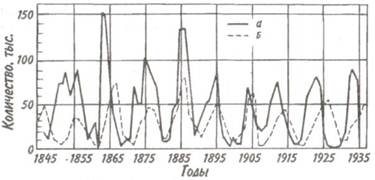
 Рис. 7.9. Периодические колебания
Рис. 7.9. Периодические колебания
популяций зайца-беляка (а) и рыси (б),
установленные по числу шкурок, заготовленных
«Компанией Гудзонова залива» (из Дажо, 1975).
сравнению с численностью рыси обычно сдвинут на 1 -2 года назад. Это вполне понятно: рысь питается зайцами, а потому колебания ее численности должны быть связаны с колебанием численности ее добычи.
Циклические изменения численности со средним периодом в 4 года характерны для обитателей тундры: полярной совы, песца, а также лемминга. По мнению многих ученых, периодичность 9,6-летних циклов у зайца-беляка и рыси определяется явлениями, происходящими в космосе, и так или иначе связана с солнечными циклами. Подобная зависимость отмечается, например, у атлантического канадского лосося, максимум численности которого наблюдается через каждые 9-10 лет.
Причины, вызывающие другие периодические флуктуации численности, хорошо известны. У берегов Перу наблюдается трансгрессия теплых вод к югу. известная под названием Nino. Приблизительно один раз в 7 лет теплые воды вытесняют с поверхности холодные. Температура воды быстро поднимается на 5°С, изменяется соленость, гибнет планктон, насыщая воду продуктами распада. В результате погибает рыба, а за ней морские птицы. Случаи сезонных изменений численности популяций хорошо известны всем. Тучи комаров, большое
количество населяющих леса птиц обычно наблюдаются в определенный период года. В другие сезоны популяции этих видов могут практически исчезать.