Репликация ДНК
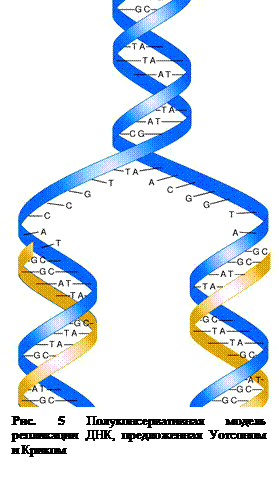
Согласно предложенной в 1953 г Уотсоном и Криком схеме репликацииспиралевидная двухцепочная ДНК сначала расплетается, и цепи расходятся (рис. 5). При этом к нуклеотидам каждой цепи присоединяются комплементарныенуклеотиды, которые с помощью ферментов ДНК-полимераз связываются в новые полинуклеотидные цепи. В результате из одной образуются две дочерние двухцепочечные молекулы ДНК (рис. 5). Таким образом, каждая реплицированная дочерняя молекула ДНК состоит из одной «старой» и одной «новой» цепей, вот почему такой способ репликации получил название - полуконсервативный.
 В 1958 г М. Мезелсон и Ф. Сталь в блестящем эксперименте убедительно доказали именно полукон-сервативный характер репликации ДНК предсказанный Уотсоном и Криком.
В 1958 г М. Мезелсон и Ф. Сталь в блестящем эксперименте убедительно доказали именно полукон-сервативный характер репликации ДНК предсказанный Уотсоном и Криком.
Дальнейшие исследования показали, что процесс полуконсервативной репликациимолекул ДНК начинается в определенной точке инициации (ori). В хромосомах эукариот имеется по нескольку таких точек. Цепи ДНК в точке инициации репликации разъединяются (раскручиваются) под влиянием ферментагеликазы (рис. 6). Возникает репликационная вилка с одноцепочечными участками ДНК, которые становятся матрицами для репликации (рис. 6). Эти участки связываются с белками SSB (single-stranded binding), которые не позволяют им вновь соединиться в двойную спираль. Возникающая суперскрученность и напряжение в нераскрученной части ДНК репликациооной вилки (рис. 6) снимает ферментный комплекс топоизомераза (ДНК-гираза у прокариот).
ДНК-полимераза, осуществляет процесс репликации в направлении 5'-3', она способна присоединять нуклеотиды только к 3'-ОН группе предыдущего нуклеотида и для синтеза новой цепи ей требуется затравка (праймер) со свободным 3'-концом. Поэтому сначала на ДНК-матрице с помощьюпраймазы (РНК-полимеразы) синтезируется короткий (~10 нуклеотидов) фрагмент РНК. Именно к такому РНК-праймеру ДНК-полимераза присоединяет дезоксинуклеотиды, синтезируя новую цепь (рис. 6). Затем РНК-праймер вырезается, замещаясь фрагментом ДНК.
Репликацияначинаетсяна материнской цепи, идущей от точки инициации в направлении 3'-5' и идет непрерывно в виде сплошной линии. Эта цепь называется лидирующей(рис. 6). Синтез на второй цепи идет в обратном направлении в виде отдельных коротких (200-2000 нуклеотидов) фрагментов Оказаки, названных так по имени открывшего их ученого. Эта цепь получила название запаздывающей. После завершения синтеза РНК праймеры в составе фрагментов Оказакизаменяются на ДНК и все фрагменты соединяются при помощи фермента лигазы в общую полинуклеотидную цепочку. В результате репликации образуются две идентичные молекулы ДНК.
