Трансляция
Трансляция наряду с репликацией и транскрипцией является еще одним важнейшим этапом реализации генетической информации в клетке. В самом общем виде трансляция – это процесс биосинтеза белка по матрице иРНК, который протекает на рибосомах. В этом процессе кроме информационной РНК принимают активное участие еще два типа РНК – транспортные и рибосомные.
 Транспортные РНК. Роль тРНК заключается в том, что они переносят аминокислоты к рибосомам и участвуют в процессе синтеза белка. Транспортные РНК, которых насчитывается более 60-ти, состоят из 75-90 нуклеотидов и имеют структуру в виде клеверного листа (рис. 10). Каждая аминокислота присоединяется к определенной тРНК. На одном конце тРНК находится акцепторный триплет ЦЦА, к аденину которого присоединяется специфическая аминокислота. На другом конце (в антикодонной петле) каждой тРНК находится антикодон– специфический триплет, с помощью которого тРНК «узнает» соответствующий комплементарный кодон в иРНК, и тем самым определяет место, куда должна быть поставлена данная аминокислота в синтезируемой молекуле белка. Боковые петли тРНК, по-видимому, используются для связывания с рибосомой и со специфической аминоацил-тРНК-синтетазой.
Транспортные РНК. Роль тРНК заключается в том, что они переносят аминокислоты к рибосомам и участвуют в процессе синтеза белка. Транспортные РНК, которых насчитывается более 60-ти, состоят из 75-90 нуклеотидов и имеют структуру в виде клеверного листа (рис. 10). Каждая аминокислота присоединяется к определенной тРНК. На одном конце тРНК находится акцепторный триплет ЦЦА, к аденину которого присоединяется специфическая аминокислота. На другом конце (в антикодонной петле) каждой тРНК находится антикодон– специфический триплет, с помощью которого тРНК «узнает» соответствующий комплементарный кодон в иРНК, и тем самым определяет место, куда должна быть поставлена данная аминокислота в синтезируемой молекуле белка. Боковые петли тРНК, по-видимому, используются для связывания с рибосомой и со специфической аминоацил-тРНК-синтетазой.
Рибосомные РНК. Размер рибосомных рРНК составляет 120–3100 нуклеотидов. Они служат каркасом рибосом и способствуют первоначальному связыванию иРНК с рибосомой в ходе биосинтеза белка.
Рибосомы являются клеточными органеллами, на которых протекает процесс биосинтеза белка. В период синтеза белка рибосомы могут объединяться в полисомы. Рибосомы состоят из двух субъединиц разного размера и формы. Размер эукариотической рибосомы составляет 80S (S – ед. Сведберга, характеризующая скорость седиментации при центрифуги-ровании). Большая субъединица величиной 60S состоит из рРНКтрех типов - 28S, 5Sи 5,8S и 50 белков, а малая величиной 40S - из 18SрРНК и 33 белков. У прокариот рибосома имеет величину 70S и состоит из большой (50S)субъединицы, в состав которой входит 23S и 5SрРНК, а также34 белка и малой (30S),состоящей из16S рРНК и 21 белка.
 |
Как уже отмечалось, трансляция заключается в том, что последовательность расположения кодонов в иРНК переводится в строго упорядоченную последовательность расположения аминокислот в молекуле синтезируемого белка (рис.11). Процесс трансляции включает два этапа: активирование аминокислот и присоединение их к «своим» тРНК и непосредственно синтез белковой молекулы. В свою очередь процесс синтеза белка на рибосомах подразделяется на три стадии: инициация, элонгация и терминация (рис.12).
Инициация. Инициация синтеза полипептидной цепи начинается с присоединения малой субъединицы рибосомы к соответствующему центру связывания на иРНК (рис. 12, А). Сигналом инициации трансляции служит кодон для метионина АУГ, который расположен в начале иРНК. К кодону АУГ своим антикодоном УАЦ присоединяется аа-тРНК с метионином (у бактерий с формилметионином). Затем к этому комплексу, присоединяется большая субъединица рибосомы. В результате образуется полная рибосома (80S), включающая молекулу иРНК и инициаторную аа-тРНК с метионином, которая располагается в пептидильном центре большой субъедницы (рис. 12, А).
Элонгация. В свободный аминоацильный центр рибосомы со второй аминокислотой поступает следующая аа-тРНК, которая своим антикодоном соединяется со строго с комплементарным кодоном иРНК (рис. 12, Б). В этот момент при помощи фермента пептидилтрансферазыпредшествующая аминокислота (метионин) своей карбоксильной группой (СООН) соединяется с аминогруппой (NH2) вновь пришедшей аминокислоты. Между ними образуется пептидная связь (–СО–NH–). В результате тРНК, принесшая метионин, освобождается, а в аминоацильном центре к тРНК присоединен уже дипептид. Дипептидил-тРНК благодаря перемещению рибосомы на один кодон при участии ферментатранслоказыибелковогофактора элонгации продвигается из аминоацильного центра в пептидильный. Освободившаяся тРНК и связанный 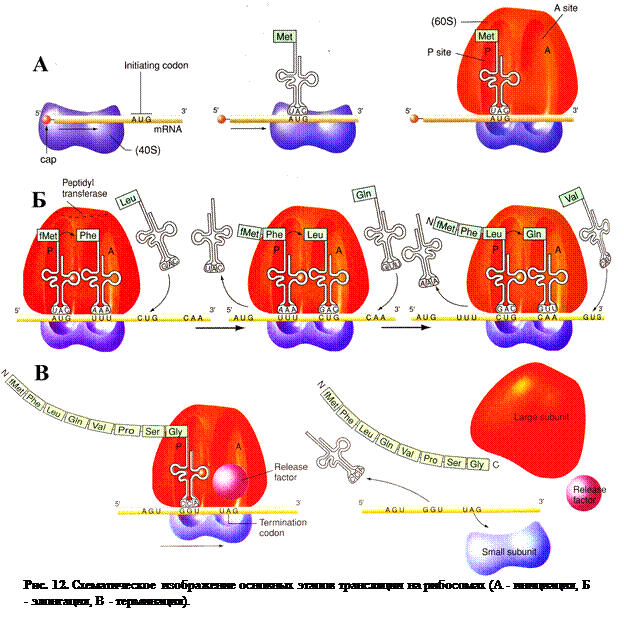 с ней кодон иРНК АУГ выходят из рибосомы (рис. 12, Б). Следующая аа-тРНК приносит новую аминокислоту в освободившийся аминоациль-ный центр в соответствии с поступившим туда кодоном. Эта аминокислота при помощи пептидной связи соединяется с предыдущей. При этом рибосома снова продвигается еще на один кодон, и процесс повторяется (рис. 12, Б).
с ней кодон иРНК АУГ выходят из рибосомы (рис. 12, Б). Следующая аа-тРНК приносит новую аминокислоту в освободившийся аминоациль-ный центр в соответствии с поступившим туда кодоном. Эта аминокислота при помощи пептидной связи соединяется с предыдущей. При этом рибосома снова продвигается еще на один кодон, и процесс повторяется (рис. 12, Б).
Терминация. Как только в аминоацильный центр рибосомы поступит один из терминирующий кодонов иРНК (УАА, УАГ или УГА), к нему присоединяется белковый фактор терминации и блокирует дальнейшую элонгацию цепи (рис. 12, В). После этого синтезированная полипептидная цепь отделяется от тРНК, рибосомные субъединицы диссоциируют и освобождают тРНК и иРНК, которые могут принять участие в синтезе следующей полипептидной цепи (рис. 12, В).
На одной молекуле иРНК работает не одна рибосома, а многие (до 100). На каждой из рибосом строится полипептидная цепь. У бактерий транскрипция и трансляция связаны между собой и трансляция начинается до завершения синтеза иРНК на ДНК. Образующиеся при синтезе полипептидные цепи претерпевают ряд посттрансляционных преобразований и только после этого начинают выполнять в организме свои специфические функции.