ОРИЕНТИРОВОЧНЫЙ РЕФЛЕКС - раздел Психология, Основы психофизиологии: Учебник / Отв. ред. Ю.И. Александров
Ориентировочный Рефлекс, Или Рефлекс «Что Такое?» Был Открыт ...
Ориентировочный рефлекс, или рефлекс «Что такое?» был открыт И.П. Павловым. Он описал его как комплекс двигательных реакций, который возникал на неожиданное появление нового стимула. Собака поворачивала голову, глаза, настораживала уши в направлении нового раздражителя. В работах Ю. Конорского [1970] этот комплекс соматических реакций получил название рефлекса прицеливания.
Рассматривая ориентировочный рефлекс как реакцию, направленную на восприятие нового стимула, И.П. Павлов обратил основное внимание на его тормозную, прерывающую функцию в отношении текущей условно-рефлекторной деятельности. Во время ориентировочной реакции условный рефлекс либо полностью подавлялся, либо его выполнение ухудшалось. Это свойство ориентировочной реакции нарушать текущую деятельность, в том числе и процесс обучения (выработку условного рефлекса), привело И.П. Павлова к мысли о необходимости построить исследовательский корпус в виде «Башни молчания». Это позволило бы исключить тормозные эффекты ориентировочного рефлекса (ОР) на формирование и исполнение условных рефлексов.
Хотя вначале ОР и представлялся И. П. Павлову и его коллегам преимущественно как внешний тормоз условного рефлекса, они описали многие его особенности, которые выходили за пределы тормозной функции. Так был открыт растормаживающий эффект ориентировочной реакции. Он состоял в том, что ранее выработанные, а затем угашенные условные связи под влиянием ОР вновь восстанавливались. Сегодня мы интерпретировали бы эти данные как способность восстанавливать подавленные связи в процедурной долговременной памяти. В лаборатории И.П. Павлова также были получены первые факты о влиянии ОР на формирование условного рефлекса. Во время процедуры выработки условной связи, а также при формировании ассоциации между двумя индифферентными раздражителями было отмечено усиление или появление ориентировочных реакций. В опытах И.О. Нарбутович и Н.А. Подкопаева [1936] с выработкой у собаки ассоциации в виде двух последовательно следующих индифферентных стимулов (звук – свет или вертушка – прерывистый шум) было показано, что при отсутствии на них ориентировочных реакций ее формирование идет плохо. В этих же опытах по ОР судили о сформированности ассоциации. Когда собака вырабатывала связь между звуком и светом, можно было видеть, что она поворачивает голову в сторону света еще до его предъявления.
Однако на том этапе изучения ориентировочного рефлекса еще не сложилось полного представления как о его взаимоотношениях с условным рефлексом, так и о физиологических механизмах и структуре самого ориентировочного рефлекса.
П. К. Анохин выделил три формы взаимоотношений ориентировочного рефлекса с текущей деятельностью. К первой он относит «конфликтные отношения», когда ориентировочная активность оказывает тормозный эффект. Эти отношения детально были изучены в лаборатории И.П. Павлова. Вторая форма отражает ассимиляцию ориентировочной реакции оминирующей деятельностью. В этом случае ОР не тормозит, а усиливает текущую деятельность. Например, в ситуации оборонительной доминанты появление нового стимула вызывает не ориентировочный, а оборонительный рефлекс. К третьей форме отношений относится трансформация ОР в поведенческую реакцию, связанную со скрытой, внешне непроявляющейся доминантой. Анализируя эти данные, П.К. Анохин [1968] пришел к выводу том, что эффект ОР существенно зависит от предыдущего опыта животного и от ранее сформировавшихся и зафиксированных в памяти доминант.
Открытие в 1929 г. Г. Бергером альфа-ритма человека и последующее совершенствование техники записи волновой активности мозга позволили связать ОР с депрессией альфа-ритма. Важное значение для изучения ОР имело открытие Г. Моруцци и X. Мэгуном (1949) ретикулярной активирующей системы мозга, позволившее связать блокаду альфа-ритма с реакцией активации, регулируемой этой системой. Наиболее всесторонне ориентировочный рефлекс был исследован в работах Е.Н. Соколова и его коллег.
Полиграфическая регистрация электроэнцефалографических, вегетативных и двигательных реакций животных и человека в условиях многократного повторения индифферентных стимулов привела к представлению об ориентировочном рефлексе как о многокомпонентной реакции активации. Е.Н. Соколов [I960], а затем Р. Линн [Lynn, 1966] описали систему компонентов ОР. На уровне неокор-текса ОР представлен ЭЭГ-реакцией активации (блокадой альфа-ритма, усилением бета- и гамма- колебаний), коррелирующей с увеличением возбудимости, реактивности и лабильности у нейронов коры. К вегетативным компонентам ОР относят расширение зрачков, увеличение кожной проводимости, снижение ЧСС, изменение дыхания, увеличение дыхательной аритмии в частотном спектре ритмограммы сердца, расширение сосудов головы и сужение сосудов рук. В моторной сфере ориентировочный рефлекс представлен поворотом головы, движением глаз, настораживанием ушей в направлении нового стимула и увеличением мышечного тонуса, обеспечивающего повышенную готовность к моторным реакциям. Под влиянием ОР сенсорная чувствительность анализаторов, измеряемая по субъективным реакциям, увеличивается, что получило название сенсорного компонента ориентировочного рефлекса.
Характер электроэнцефалографических, вегетативных и моторных реакций во время ОР позволяет понять суть рефлекса «Что такое?». Ориентировочный рефлекс обеспечивает лучшее восприятие вызвавшего его стимула и готовность быстро реагировать, если того потребует ситуация. Ради этого прерывается текущая деятельность и ресурсы внимания обращаются на новый стимул.
Отличительной особенностью ОР является его привыкание или угашение, развивающееся при повторении индифферентного стимула. Е.Н. Соколов [1958] выделил две формы ориентировочной реакции: генерализованную и локальную, которые выражают разные этапы процесса угашения ОР. В зависимости от длительности ОР различают тонический и фазический ориентировочный рефлекс.
Первоначально новый стимул вызывает генерализованный ОР, связанный с возбуждением ретикулярной формации ствола мозга. Он характеризуется ЭЭГ-активацией, которая охватывает всю кору в течение достаточно длительного времени (тоническая форма ОР, часто возникающая параллельно с генерализованным ОР; см. гл. 8). Ее легко увидеть у субъекта, находящегося в дремотном состоянии. Тогда говорят о реакции пробуждения (arousal reaction). Подобную реакцию также наблюдали Шарплесс и Джаспер [Sharpless, Jasper, 1956] у кошек и назвали ее тонической реакцией пробуждения. Генерализованный ориентировочный рефлекс быстро угасает, обычно после 10–15 применений индифферентного стимула.
Это позволяет выявить локальный ориентировочный рефлекс, который более устойчив к угашению и требует 30 и более применений стимула. Его генерацию связывают с активацией неспецифического таламуса. При локальном ОР реакция ЭЭГ-активации сохраняется в сенсорной коре, соответствующей модальности повторяемого раздражителя. Так, свет продолжает вызывать блокаду альфа-ритма, но только в зрительной коре, звук – блокаду тау-ритма, регистрируемого от височной области, тактильное и проприоцептивное раздражение при пассивном подъеме пальца руки вызывает депрессию мю-ритма (роландического ритма) локально в сенсомоторной коре. Длительность реакции сокращается, она становится фазической, появляясь только на включение и выключение стимула (фазическая реакция пробуждения по Шарплису и Джасперу). Продолжая повторять стимул, можно добиться полного угашения ОР.
Ориентировочный рефлекс с полным набором компонентов возникает не у всех испытуемых, что выражает их индивидуальные различия. Изучение компонентного состава ОР на звуковой стимул (1000 Гц, 50 дБ) у 100 испытуемых показало, что блокада альфа-ритма в зрительной коре наблюдается у 85 человек, блокада роландического – у 53, появление КГР – у 95, движение глаз возникало у 42 испытуемых, изменение частоты дыхания – у 63, а изменение мышечного тонуса пальца наблюдалось только у одного испытуемого. Все компоненты ОР (за исключением реакции мышц) были представлены только у 11 испытуемых из 100 (Воронин, Соколов, 1960).
Хотя ориентировочный рефлекс, и относится к безусловным рефлексам, он обнаруживает способность к угашению (так как для него специфическим стимулом является «новизна», а специфической реакцией – набор компонентов ориентировочного рефлекса). Это сближает его с условным рефлексом, который сам по себе является продуктом обучения. Именно поэтому угашение ОР рассматривается как особая форма обучения, получившего название негативного обучения, которое ведет к потере способности у данного стимула вызывать ориентировочные реакции.
Угашение ОР характеризуется высокой избирательностью к физическим характеристикам стимула. Это проявляется в том, что даже небольшое изменение любого из параметров повторяющегося стимула ведет к восстановлению ОР. Стимулы, сильно отличающиеся от угашаемого, вызывают более сильные ориентировочные реакции, а стимулы, близкие ему – более слабые. Величина ОР увеличивается пропорционально количеству одновременно измененных параметров стимула.
Для объяснения этого явления Е.Н. Соколов предположил, что в процессе повторения индифферентного стимула без специального подкрепления в мозге формируется его нейрональная модель (см. также гл. 9). Мозг замечает малейшее отклонение от параметров повторяющегося стимула благодаря сравнению с его моделью, хранящейся в памяти, будь то изменение интенсивности, длительности, цвета, формы, частоты и т.д. В нервной модели запечатлеваются не только элементарные, но и сложные признаки, например комплексы раздражителей – совместное или последовательное применение нескольких раздражителей. Исключение одного из элементов комплекса или изменение их порядка вызывают ориентировочные реакции. Нервная модель стимула многомерна, она фиксирует одновременно все его параметры, что доказывается восстановлением ОР при изменении любого физического свойства повторяющегося стимула даже в случае уменьшения интенсивности или укорочения его длительности.
Новизна как специфический тригер безусловного ОР определяется сигналом рассогласования, генерируемым при несовпадении нервной модели стимула с возбуждением от предъявляемого раздражителя. Схема взаимодействия основных функциональных блоков с нервной моделью, по Е.Н. Соколову, объясняющая угашение и восстановление ОР, включает воспринимающее устройство, исполнительное устройство ОР, блок с моделью стимула, компаратор (устройство, в котором осуществляется операция сравнения стимула с сигналом, поступающим от блока с моделью), блок активирующей системы, определяющий уровень активности исполнительного устройства ориентировочного рефлекса. По мере формирования модели она усиливает свое тормозное влияние на неспецифическую систему мозга. Торможение носит избирательный характер, оно ухудшает восприятие только повторяющегося стимула. В случае несовпадения стимула с моделью в компараторе возникает сигнал рассогласования, который активирует исполнительное устройство ОР.
Следует отметить, что некоторые исследователи увеличивают количество факторов, вызывающих ОР. Класс явлений, названных Д. Берлайном «сличительными признаками», которые стимулируют ориентировочно-исследовательское поведение, включает, помимо новизны, удивление, неопределенность, двусмысленность и сложность. Однако, по мнению Р. Линна [Lynn, 1966], они могут быть интерпретированы как разные формы новизны. Так, удивление связано с появлением чего-то нового в последовательности стимулов, а сложность, необычность и двусмысленность представляют новизну в паттерне стимулов.
Изучение нейронных коррелятов ориентировочного рефлекса выявило существование особого класса нейронов – нейронов новизны, которые были обнаружены в разных структурах мозга. Впервые нейроны новизны были найдены в зрительной покрышке лягушки Дж. Летвиным и др. [1964]. Эти клетки переставали отвечать на повторяющееся движение зрительного объекта, совершаемое в одном и том же направлении. Изменение траектории движения восстанавливало ответ нейрона в виде спайковых разрядов. Нейроны новизны были эфферентными, а их длинные аксоны оканчивались в сетчатке. Эти нейроны сходны с «нейронами внимания», выделенными Д. Хьюбелом и др. (Hubel et al., 1959) в слуховой коре кошки. Нейроны со сходными характеристиками были также найдены в ретикулярной формации ствола мозга кошки (Huttenlocher, 1961; Lindsley et al., 1973). Изучение неспецифического таламуса кролика показало существование особой формы нейронной реакции активации, отличной от простого учащения спайковых разрядов. Новые стимулы вызывали десинхронизацию спайковой активности нейрона, которая состояла из разрушения его пачечной активности, коррелирующей с волнами ЭЭГ, и замены ее на рандоминизированные одиночные спайки, появление которых совпадало с блокадой альфа-подобной активности [Данилова, 1970, 1985]. Реакция десинхронизации нейронов таламуса демонстрирует все свойства ориентировочной реакции.
Вначале формирование нервной модели стимула Е.Н. Соколов связывал с функцией корковых нейронов. Однако после открытия О.С. Виноградовой в поле САЗ гиппокампа двух групп нейронов, реакции которых обнаружили характеристики ОР (В-нейроны, реагирующие возбуждением, и Т-нейроны, отвечающие на новый стимул торможением), он связал нервную модель стимула с нейронами гиппокампа.
Е.Н.Соколов [1995] рассматривает эти две группы нейронов гиппокампа как «нейроны новизны» (В-нейроны) и «нейроны тождества» (Т-нейроны). Их совместное действие позволяет подчеркивать новые раздражители и ослаблять действие привычных стимулов за счет противоположного влияния на активирующую и инактивирующую части модулирующей системы мозга. Формирование нервной модели определяется тем, что множество нейронов-детекторов конвергируют на нейроны поля САЗ гиппокампа (нейроны новизны и тождества). Они создают на каждом из них универсальное рецептивное поле, охватывающее всю рецепторную поверхность животного [Соколов, 1981]. При действии стимула происходит селективное включение детекторов. С повторением раздражителя возбуждение детекторов сохраняется прежним. Однако меняются их синаптические контакты на нейронах гиппокампа. Веса задействованных пластических синапсов на гиппокампальных нейронах уменьшаются пропорционально силе приходящего возбуждения. В результате возбудительный ответ нейрона новизны на повторяющийся стимул подавляется и от него регистрируется лишь фоновая активность. С повторением стимула параллельно меняется и реакция нейронов тождества, с которыми детекторы связаны не возбуждающими, а тормозными пластическими синапсами. Их тормозная реакция на новый стимул постепенно уменьшается, и нейрон сохраняет свою фоновую активность. Е.Н. Соколов предположил, что реципрокные сигналы от нейронов новизны и тождества противоположно воздействуют на активирующую и синхронизирующую системы мозга. Новый раздражитель вызывает реакцию активации, так как возбуждает активирующую и тормозит синхронизирующую (инактивирующую) системы мозга. Привычный стимул перестает возбуждать активирующую неспецифическую систему и при этом через нейроны тождества стимулирует неспецифические тормозные влияния.
Таким образом, нервная модель стимула представлена на нейронах новизны и тождества матрицами потенциированных синапсов, связанных со свойствами раздражителя и отражающими его конфигурацию. Новый стимул активирует новые синапсы нейронов новизны и тождества, еще не подвергнувшихся пластическим перестройкам. Это определяет появление ориентировочного рефлекса за счет возбуждения активирующей и подавления инактивирующей систем мозга. Величина ОР увеличивается пропорционально величине различия (рассогласования) между новым стимулом и сформированной нервной моделью [Соколов, 1995].
Дальнейший прогресс в понимании механизма ОР связан с применением компьютерной техники усреднения при изучении потенциалов мозга, связанных с событиями (ССП; см. гл. 16). Значительный вклад в идентификацию и классификацию различных компонентов ССП и в поиски мозговых аналогов сигналов рассогласования внесли работы Р. Наатанена и его коллег. Им выявлена специфическая мозговая волна, названная негативностью рассогласования (HP). Она получена в опытах, в которых в случайном порядке и с короткими межстимульными интервалами, (не более 10–14 с) следовали стандартный (часто повторяющийся, 80–90%) и девиантные (несколько отклоняющиеся от стандарта и редко предъявляемые, 20–10%) стимулы. При этом внимание испытуемого было направлено на чтение интересной книги. Р. Наатанен для извлечения HP предложил процедуру вычитания ССП на стандартный стимул из ССП на девиантный стимул. Им представлено большое количество экспериментальных доказательств связи HP с операцией сличения действующего стимула с короткоживущим (не более 20 с) следом стандартного стимула в памяти. Чем сильнее они отличаются, тем больше амплитуда и короче латенция HP [Наатанен, 1997].
Исследователями выявлено два компонента HP: модально-специфический и фронтальный. Определяя локусы генерации модально-специфической HP для звуковых стимулов с помощью МЭГ, они нашли их в специфической слуховой коре, где, как предполагает Р. Наатанен, хранятся короткоживущие сенсорные следы. Для разных параметров звуковых стимулов (частоты, интенсивности и пространственного расположения) обнаружены разные HP с несовпадающими локусами их генерации. Предполагают, что сенсорно-специфическая HP трансформируется во фронтальную HP.
Процесс сличения стимула с короткоживущим следом памяти, который представлен в HP, протекает автоматически и не требует сознания. В частности, это подтверждается тем, что HP может возникать и на подпороговые девиации. При этом чем больше амплитуда подпороговой HP, тем ниже субъективный порог различения испытуемого. HP рассматривается как начальная фаза обработки информации о сигнале, за которой может следовать фаза непроизвольного внимания (безусловный ориентировочный рефлекс) к девиантному стимулу. Р. Наатанен связывает его с волной Н2б.
HP как выражение сигнала рассогласования отличается от классического ОР, который, как уже упоминалось, также возникает в результате рассогласования нервной модели стимула и наличного возбуждения от раздражителя. Однако в последнем случае время жизни нервной модели значительно больше, что позволяет при угашении ОР и формировании нервной модели использовать длительные межстимульные интервалы порядка нескольких десятков секунд.
Сравнивая HP с классическим ОР, Р.Наатанен [1997] отмечает, что процесс генерации сенсорно-специфической HP имеет очевидное сходство с процессом рассогласования, постулируемым Е.Н. Соколовым для возникновения ОР, и в то же время между ними имеется существенное различие. HP – первое событие, возникающее в активности мозга, которое направлено на инициацию ОР в ответ на физическое изменение стимула, когда стимулы предъявляются с достаточно короткими межстимульными интервалами, чтобы вызвать HP. Появление ОР в начале стимульной последовательности и в ответ на изменение стимула при длительных межстимульных интервалах имеет другой механизм, отличный от вызова ОР на основе HP.
Другой сигнал рассогласования, который более тесно связан с классическим ОР, был выявлен М. Фербатеном и др. [Verbaten et al., 1986]. Для этого был использован не метод усреднения ССП, а процедура удаления шумового компонента, что позволило исследовать динамику одиночных ССП на стимулы, предъявляемые с большими интервалами. Исследователи нашли, что при угашении ОР (его компонента в виде увеличения кожной проводимости пальцев) параллельно идет процесс уменьшения неспецифической части волны Н1. Новый стимул восстанавливает неспецифический компонент H1 и реакцию повышения кожной проводимости. Выделенная часть компонента Н1 демонстрирует неспецифические свойства ОР и зависимость от долгоживущего следа памяти (десятки секунд).
Таким образом, сигналы рассогласования, участвовавшие в генерации непроизвольного ОР, возникают по крайней мере на двух разных типах следовых процессов: короткоживущей и долгоживущей моделях стимула. Следует отметить, что теория ориентировочного рефлекса Е.Н. Соколова, несмотря на ее большую популярность, все же подвергалась критике. Так, отмечалось, что с ее позиций трудно объяснить появление ОР в случаях когнитивного рассогласования, т.е. когда не подтверждается ожидание относительно последовательности стимулов [Underwood, Moray, 1971]. Например, ориентировочная реакция возникает на цифру «9» в последовательности 1, 2, 3, 4, 5, 6, 7, 9. Другое направление критики связано с утверждением о том, что теория ОР Е.Н. Соколова не объясняет появления ОР на значимые стимулы [Bernstein, 1979; Maltzman, 1979]. Сторонники концепции значимости считают, что решающее значение для появления ОР имеют корковые процессы интерпретации и усиления. С этих позиций теория ОР, возникающего на любое физическое изменение стимула, рассматривается как не оринтированная на реальное поведение и деятельность [Bernstein, 1969].
Однако участники полемики относительно значимости ОР не учитывают, что наряду с безусловным ОР Е.Н. Соколов [1958] вводит понятие условного ориентировочного рефлекса, возникающего на значимый сигнальный стимул. Он показал, что придание раздражителю сигнального значения (обычно с помощью инструкции или речевого подкрепления, побуждающих человека отвечать определенной реакцией на определенный стимул) восстанавливает и усиливает ОР на условный сигнал. Эффект усиления ОР сохраняется до тех пор, пока условный рефлекс не достигнет стадии автоматизации. На рис. 10.1 представлено усиление КГР, измеряемого как падение сопротивления кожи, по Фере, при придании звуковому стимулу сигнального значения. По инструкции испытуемый должен был сжимать руку в кулак при предъявлении звука после предварительного угашения на него безусловного ОР. Выработку условного ориентировочного рефлекса можно увидеть при формировании ассоциации: звук 70 дБ – звук 90 дБ (рис. 10.2). После угашения КГР на индифферентный звук 70 дБ присоединение к нему второго стимула – 90 дБ восстанавливает реакцию, которая усиливается по мере сочетаний стимулов. Постепенно КГР-реакция на комплекс уменьшается и исчезает. Однако изолированное применение стимулов вызывает усиленный ОР, выражающий рассогласование стимула со сформированной в памяти моделью комплекса.
Сравнивая форму ССП на редко предъявляемые девиантные стимулы, когда они были релевантными и испытуемый должен был на них как-то реагировать и когда те же стимулы им игнорировались (испытуемый читал книгу), Р. Наатанен выделил среди компонентов ССП особый потенциал, который коррелировал с произвольным вниманием. Эта волна получила название процессной негативности (ПН). Под влиянием инструкции быть внимательным к особым стимулам, ПН усиливается на все стимулы, но особенно на релевантные. Чем короче межстимульный интервал, тем раньше возникает ПН (в среднем ее ЛП около 150 мс, но она может появляться и через 50 мс).

Рис.10.1. Усиление КГР – компонента ориентировочного рефлекса на звук, на который испытуемый должен был реагировать движением руки (10 – 24):
9 – инструкция; 1–8– индифферентные звуковые стимулы до инструкции. Видно, что усиление фазических КГР-реакций сопровождается тоническим снижением сопротивления кожи, которое сохраняется до 24-го применения стимула. По оси ординат – сопротивление в килоомах [Соколов, 1958]
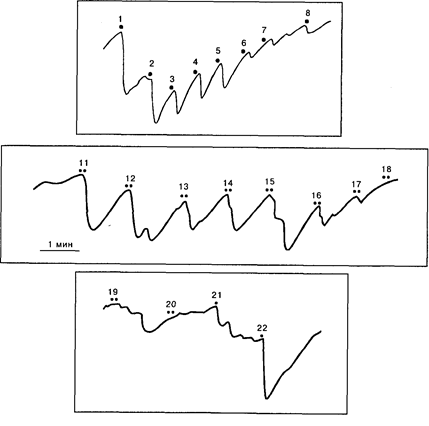
Рис.10.2. Выработка условного ориентировочного рефлекса при формировании ассоциации двух звуковых стимулов:
1-8 - звук 1000 Гц, 70 дБ, 5 с; 11–20 - комплекс: 70 дБ - 90 дБ; 21 - 70 дБ, 22– 90 дБ. По оси ординат – сопротивление в килоомах [Соколов, 1958]
ПН не тождественна Н1, хотя и совпадает с ней по латенции. Критическим признаком, отличающим Н1 от ПН, является его инверсия при смещении активного электрода по слуховой коре относительно мостадоидов (сосцевидных отростков). Объясняя ПН, Р. Наатанен [1997] вводит понятие «следа селективного внимания», который связан с памятью. Он полагает, что произвольное внимание актуализирует в сенсорной памяти (по-видимому, во вторичной ассоциативной коре) след релевантного стимула. «След внимания» понимается им как механизм опознания стимула, так как входной сигнал всякий раз сличается со следом релевантного стимула, который непрерывно поддерживает в активном состоянии произвольное внимание. Чем ближе стимул к релевантному, тем больше ПН. Р. Наатанен выделяет раннюю ПН с локусом в ассоциативной коре, выражающей завершение процесса сличения, и позднюю ПН с фронтальной локализацией. Последняя отражает произвольное усилие, направленное на дальнейшую обработку стимула, включающее произвольный ОР на значимый стимул.
Суммируя результаты собственных исследований с данными Р. Наатанена и его коллег, Е.Н. Соколов [1997] выделяет два типа энграмм, с которыми связано возникновение непроизвольного и произвольного ОР. Когда единицы кратковременной памяти подключены к нейронам новизны и тождества прямо в виде потенциированных синапсов от детекторов-признаков, сигнал рассогласования, возникающий при несовпадении стимула с энграммой, определяет генерацию непроизвольного ОР. Его сигнал рассогласования представлен в HP и неспецифическом компоненте Н1. Если же к нейронам новизны и тождества подключены единицы долговременной памяти и они обеспечивают произвольное удержание шаблона, то совпадение стимула с шаблоном порождает сигнал согласования или совпадения. В терминах нейронных популяций он представлен ПН. Сигнал совпадения достигает нейронов активирующей системы и вызывает условный ОР, направленный на значимый стимул. В случае несовпадения стимула с шаблоном сигнал совпадения и ПН не возникают и ОР на значимый стимул не появляется. При этом стимулы, отличающиеся от релевантного, вызывают безусловный ОР, непроизвольное внимание, которое представлено негативностью рассогласования и неспецифическим компонентом Н1.
Все темы данного раздела:
ОБЩИЕ СВЕДЕНИЯ
Традиционно со времен французского физиолога Биша (начало XIX в.) нервную систему разделяют на соматическую и вегетативную, в каждую из которых входят структуры головного и спинного
НЕЙРОН. ЕГО СТРОЕНИЕ И ФУНКЦИИ
Мозг человека состоит из 1012 нервных клеток. Обычная нервная клетка получает информацию от сотен и тысяч других клеток и передает сотням и тысячам, а количество соединен
Размер и форма
Размеры нейронов могут быть от 1 (размер фоторецептора) до 1000 мкм (размер гигантского нейрона у морского моллюска Aplysia) (см. [Сахаров, 1992]). Форма нейронов также исключительн
Цвет нейронов
Следующая внешняя характеристика нервных клеток – это их цвет. Он также разнообразен и может указывать на функцию клетки – например, нейроэндокринные клетки имеют белый цвет. Желтый
Синапсы
Биофизический и клеточно-биологический подход к анализу нейронных функций, возможность идентификации и клонирования генов, существенных для сигнализации, вскрыли тесную связь между
Электрическая возбудимость
Все функции, свойственные нервной системе, связаны с наличием у нервных клеток структурных и функциональных особенностей, обеспечивающих возможность генерации под влиянием внешнего
Пейсмекер
Один из удивительных видов электрической активности нейронов, регистрируемой внутриклеточным микроэлектродом, – это пейсмекерные потенциалы. А. Арванитаки и Н. Халазонитис [Arvanita
МЕТОДЫ ПСИХОФИЗИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
Физиологические процессы, как правило, скрыты от внешнего наблюдения, поэтому они длительное время оставались вне области интересов психологов, занимавшихся в основном исследованием
РЕГИСТРАЦИЯ ИМПУЛЬСНОЙ АКТИВНОСТИ НЕРВНЫХ КЛЕТОК
Изучение активности нервных клеток, или нейронов, как целостных морфологических и функциональных единиц нервной системы, безусловно, остается базовым направлением в психофизиологии.
ЭЛЕКТРОЭНЦЕФАЛОГРАФИЯ
Среди методов электрофизиологического исследования ЦНС человека наибольшее распространение получила регистрация колебаний электрических потенциалов мозга с поверхности черепа – элек
МАГНИТОЭНЦЕФАЛОГРАФИЯ
Активность мозга всегда представлена синхронной активностью большого количества нервных клеток, сопровождаемой слабыми электрическими токами, которые создают магнитные поля. Регистр
ПОЗИТРОННО-ЭМИССИОННАЯ ТОМОГРАФИЯ МОЗГА
В современных клинических и экспериментальных исследованиях все большее значение приобретают методы, дающие визуальную картину мозга субъекта в виде среза на любом уровне, построенн
ОКУЛОГРАФИЯ
Движения глаз являются важным показателем в психофизиологическом эксперименте. Регистрация движений глаз называется окулографией.
С одной стороны, окулографический показате
ЭЛЕКТРОМИОГРАФИЯ
Электромиография – это регистрация суммарных колебаний потенциалов, возникающих как компонент процесса возбуждения в области нервномышечных соединений и мышечных волокнах при поступ
ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ КОЖИ
Электрическая активность кожи (ЭАК) связана с активностью потоотделения, однако физиологическая основа ее до конца не изучена. Из центральной нервной системы к потовым железам посту
ПЕРЕДАЧА И ПЕРЕРАБОТКА СЕНСОРНЫХ СИГНАЛОВ
Сенсорные сигналы несут в мозг внешнюю информацию, необходимую для ориентации во внешней среде и для оценки состояния самого организма. Эти сигналы возникают в воспринимающих элемен
Сенсорная рецепция
Рецептором называют специализированную клетку, эволюционно приспособленную к восприятию из внешней или внутренней среды определенного раздражителя и к преобразовани
Сенсорные пороги
АБСОЛЮТНАЯ ЧУВСТВИТЕЛЬНОСТЬ СЕНСОРНОЙ СИСТЕМЫ
Абсолютная чувствительность сенсорной системы основана на ее свойстве обнаруживать слабые, короткие или маленькие по ра
ПЕРЕДАЧА И ПРЕОБРАЗОВАНИЕ СИГНАЛОВ
Процессы передачи и преобразования сигналов обеспечивают поступление в высшие сенсорные центры наиболее важной (существенной) информации о сенсорном событии в такой форме, которая у
КОДИРОВАНИЕ ИНФОРМАЦИИ
Кодированием называют совершаемое по определенным правилам преобразование информации в условную форму – код. В сенсорной системе сигналы кодируются двоичным кодом, т.е. наличием или
ДЕТЕКТИРОВАНИЕ СИГНАЛОВ
Детектированием называют избирательное выделение сенсорным нейроном того или иного признака раздражителя, имеющего поведенческое значение. Осуществляют такой анализ нейроны-детектор
ОПОЗНАНИЕ ОБРАЗОВ
Это конечная и наиболее сложная операция сенсорной системы. Она заключается в отнесении образа к тому или иному классу объектов, с которыми ранее встречался организм, т.е. в классиф
АДАПТАЦИЯ СЕНСОРНОЙ СИСТЕМЫ
Сенсорная система обладает способностью приспосабливать свои свойства к условиям среды и потребностям организма. Сенсорная адаптация – это общее свойство сенсорных
ВЗАИМОДЕЙСТВИЕ СЕНСОРНЫХ СИСТЕМ
Взаимодействие сенсорных систем осуществляется на спинальном, ретикулярном, таламическом и корковом уровне. Особенно широка интеграция сигналов в ретикулярной формации. В коре мозга
МЕХАНИЗМЫ ПЕРЕРАБОТКИ ИНФОРМАЦИИ В СЕНСОРНОЙ СИСТЕМЕ
Переработка информации в сенсорной системе осуществляется с помощью процессов возбудительного и тормозного межнейронного взаимодействия. Это взаимодействие осуществляется по горизон
ОБЩИЕ СВОЙСТВА СЕНСОРНЫХ СИСТЕМ
Сенсорной системой называют часть нервной системы, воспринимающую внешнюю для мозга информацию, передающую ее в мозг и анализирующую ее. Сенсорная система состоит из воспринимающих
Общие принципы организации сенсорных систем
Все сенсорные системы человека организованы по некоторым общим принципам. Важнейшие из них следующие: многослойность, многоканальность, наличие так называемых «сенсорных воронок», а
ЗРИТЕЛЬНАЯ СИСТЕМА
Зрение – один из важнейших органов чувств человека. Оно эволюционно приспособлено к восприятию узкой части диапазона электромагнитных излучений (видимый свет). Зрительная система да
Строение и функции оптического аппарата глаза
Глазное яблоко имеет шарообразную форму, что облегчает его повороты для наведения на рассматриваемый объект и обеспечивает хорошую фокусировку изображения на всей светочувствительно
Аккомодация
Аккомодацией называют приспособление глаза к ясному видению объектов, расположенных на разном расстоянии (подобно фокусировке в фотографии). Для ясного видения объекта необходимо, ч
Аномалии рефракции глаза
Две главные аномалии рефракции глаза – близорукость (миопия) и дальнозоркость (гиперметропия). Эти аномалии обусловлены не недостаточностью преломляющих сред глаза, а изменением дли
Зрачок и зрачковый рефлекс
Зрачок – это отверстие в центре радужной оболочки, через которое свет проходит в глаз. Он повышает четкость изображения на сетчатке, увеличивая глубину резкости глаза и устраняя сфе
Структура и функции сетчатки
Сетчатка – это внутренняя светочувствительная оболочка глаза. Она имеет сложную многослойную структуру (рис. 4.2). Здесь расположены два вида фоторецепторов (палочки и колбочки) и н
Структура и функции слоев сетчатки
Клеткипигментного эпителия образуют наружный, наиболее далекий от света, слой сетчатки. Они содержат меланосомы, придающие им черный цвет. Пигмент поглощает излишни
Нейроны сетчатки
Фоторецепторы сетчатки синаптически связаны с биполярными нервными клетками (см. рис. 4.2). При действии света уменьшается выделение медиатора из фоторецептора, что гиперполяризует
Нервные пути и связи в зрительной системе
Из сетчатки зрительная информация по волокнам зрительного нерва устремляется в мозг. Нервы от двух глаз встречаются у основания мозга, где часть волокон переходит на противоположную
Электрическая активность центров зрительной системы
При действии света в рецепторах, а затем и в нейронах сетчатки генерируются электрические потенциалы, отражающие параметры действующего раздражителя (рис. 4.4 а, а)
Световая чувствительность
Абсолютная чувствительность зрения. Чтобы возникло зрительное ощущение, свет должен обладать некоторой минимальной (пороговой) энергией. Минимальное количество кван
Зрительная адаптация
При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной системы к условиям яркой освещенно
Дифференциальная чувствительность зрения
Если на освещенную поверхность с яркостью I падает добавочное освещение dI, то, согласно закону Вебера, человек заметит разницу в освещенности только если dI/I
Яркостной контраст
Взаимное латеральное торможение зрительных нейронов (см. гл. 3) лежит в основе общего, или глобального яркостного контраста. Так, серая полоска бумаги, лежащая на светлом фоне, каже
Слепящая яркость света
Слишком яркий свет вызывает неприятное ощущение ослепления. Верхняя граница слепящей яркости зависит от адаптации глаза: чем дольше была темновая адаптация, тем меньшая яркость свет
Инерция зрения, слитие мельканий, последовательные образы
Зрительное ощущение появляется не мгновенно. Прежде чем возникнет ощущение, в зрительной системе должны произойти многократные преобразования и передача сигналов. Время «инерции зре
Цветовое зрение
Весь видимый нами спектр электромагнитных излучений заключен между коротковолновым (длина волны 400 нм) излучением, которое мы называем фиолетовым цветом, и длинноволновым излучение
Восприятие пространства
Остротой зрения называется максимальная способность различать отдельные детали объектов. Ее определяют по наименьшему расстоянию между двумя точками, которые различ
СЛУХОВАЯ СИСТЕМА
В связи с возникновением речи как средства межличностного общения, слух у человека играет особую роль. Акустические (звуковые) сигналы представляют собой колебания воздуха с разной
Структура и функции наружного и среднего уха
Наружный слуховой проход проводит звуковые колебания к барабанной перепонке, отделяющей наружное ухо от барабанной полости, или среднего уха. Это тонкая перегородка, которая колебле
Структура и функции внутреннего уха
Во внутреннем ухе находитсяулитка, содержащая слуховые рецепторы. Улитка представляет собой костный спиральный канал, который по всей длине разделен вестибулярной и
Анализ частоты звука (высоты тона)
При действии звуков разной частоты возбуждаются разные рецепторные клетки кортиева органа. В улитке сочетаются два типа кодирования высоты звука: пространственный и временной [Сомье
Слуховые ощущения
Тональность (частота) звука. Человек воспринимает звуковые колебания с частотой от 16 до 20 000 Гц. Этот диапазон соответствует 10–11 октавам. Верхняя граница часто
ВЕСТИБУЛЯРНАЯ СИСТЕМА
Вестибулярная система играет важную роль в пространственной ориентации человека. Она получает, передает и анализирует информацию об ускорениях или замедлениях, возникающих в процесс
Строение и функции рецепторного вестибулярного аппарата
Периферическим отделом вестибулярной системы является вестибулярный аппарат, расположенный в пирамиде височной кости. Он состоит из преддверия и трех полукружных каналов. Полукружны
Электрические явления в вестибулярной системе
Даже в полном покое в волокнах вестибулярного нерва регистрируется спонтанная импульсация. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при п
Рефлексы, связанные с вестибулярной стимуляцией
Нейроны вестибулярных ядер обеспечивают контроль и управление различными двигательными реакциями. Вестибуло-спинальные влияния изменяют импульсацию нейронов сегментарных уровней спи
Основные афферентные пути и проекции вестибулярных сигналов
Два основных пути поступления вестибулярных сигналов в кору мозга обезьян следующие: прямой – через вентральное постлатеральное ядро и непрямой – через вентролатеральное ядро. В кор
Функции вестибулярной системы
Вестибулярная система помогает ориентироваться в пространстве при активном и пассивном движении. При пассивном движении лабиринтный аппарат с помощью корковых отделов системы анализ
Кожная рецепция
Кожные рецепторы сосредоточены на огромной кожной поверхности (1,4–2,1 м2). В коже находится множество рецепторов, чувствительных к прикосновению, давлен
Свойства тактильного восприятия
Ощущение прикосновения и давления на кожу довольно точно локализуется человеком на определенном участке кожной поверхности. Эта локализация вырабатывается и закрепляется в онтогенез
Температурная рецепция
Температура тела человека колеблется в сравнительно узких пределах. Именно поэтому информация о температуре внешней среды, необходимой для деятельности механизмов терморегуляции, им
Болевая рецепция
Болевая, или ноцицептивная, чувствительность имеет особое значение для выживания организма, так как сигнализирует о действии чрезмерно сильных и вредных факторов. В симптомокомплекс
Мышечная и суставная рецепция (проприорецепция)
В мышцах человека содержатся три типа специализированных рецепторов: первичные окончания веретен, вторичные окончания веретен и сухожильные рецепторы Гольджи. Эти рецепторы реагирую
Передача и переработка соматосенсорной информации
Чувствительность кожи и ощущение движения связаны с проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спиноталамическому, значительно различ
ОБОНЯТЕЛЬНАЯ СИСТЕМА
Рецепторы обонятельной системы расположены в области верхних носовых ходов. Обонятельный эпителий имеет толщину 100–150 мкм и содержит около 10 млн. рецепторных клеток диаметром 5–1
Вкусовые рецепторы
Вкусовые рецепторы сконцентрированы во вкусовых почках, расположенных на языке, задней стенке глотки, мягком небе, миндалине и надгортаннике. Больше всего их на кончике языка. Кажда
Вкусовые ощущения и восприятие
У разных люден абсолютные пороги вкусовой чувствительности существенно отличаются вплоть до «вкусовой слепоты» к отдельным агентам. Абсолютные пороги вкусовой чувствительности сильн
Интерорецепторы
Механорецепторы реагируют на изменение давления в полых органах и сосудах, их растяжение и сжатие [Физиол. сенсорн. систем, 1975]. Хеморецепторы сообщают ЦНС об изменениях химизма о
Проводящие пути и центры висцеральной сенсорной системы
Проводящие пути и центры висцеральной сенсорной системы представлены, в основном, блуждающим, чревным и тазовым нервами. Блуждающий нерв передает афферентные сигналы в ЦНС по тонким
Висцеральные ощущения и восприятие
Возбуждение некоторых интероцепторов приводит к возникновению четких локализованных ощущений, т.е. к восприятию (например, при растяжении стенок мочевого пузыря или прямой кишки). В
ОСНОВНЫЕ КОЛИЧЕСТВЕННЫЕ ХАРАКТЕРИСТИКИ СЕНСОРНЫХ СИСТЕМ ЧЕЛОВЕКА
Ближняя точка ясного видения 10 см
Диаметр желтого пятна сетчатки около 0,5 мм
(1,5–2 угловых градуса)
Сила аккомодации около 10 диоптрий (D)
УПРАВЛЕНИЕ ДВИЖЕНИЯМИ
Движения (включая речь и письмо) – главное средство взаимодействия организма человека с окружением. В этом взаимодействии рефлекторные ответы, побуждаемые стимулами внешней среды, с
ОБЩИЕ СВЕДЕНИЯ О НЕРВНО-МЫШЕЧНОЙ СИСТЕМЕ
Понять принципы работы системы управления невозможно, не зная особенностей строения объекта управления. Применительно к движениям животных и человека объектом управления является оп
ПРОПРИОЦЕПЦИЯ
Для успешной реализации движений необходимо, чтобы управляющие этими движениями центры в любой момент времени располагали информацией о положении звеньев тела в пространстве и о том
ЦЕНТРАЛЬНЫЕ АППАРАТЫ УПРАВЛЕНИЯ ДВИЖЕНИЯМИ
В управлении движениями участвуют практически все отделы ЦНС – от спинного мозга до коры больших полушарий.
У животных спинной мозг может осуществлять довольно обширный кла
ДВИГАТЕЛЬНЫЕ ПРОГРАММЫ
Управление движениями немыслимо без согласования активности большого количества мышц. Характер этого согласования зависит от двигательной задачи. Так, если нужно взять стакан воды,
КООРДИНАЦИЯ ДВИЖЕНИЙ
Представление о координации движений возникло на основе наблюдений больных, которые в силу разных причин не в состоянии плавно и точно осуществлять движения, легко доступные здоровы
ТИПЫ ДВИЖЕНИЙ
Движения человека очень разнообразны, однако все это разнообразие можно свести к небольшому количеству основных типов активности: обеспечение позы и равновесия, локомоция и произвол
ВЫРАБОТКА ДВИГАТЕЛЬНЫХ НАВЫКОВ
Совершенствование двигательной функции в онтогенезе происходит как за счет продолжающегося в первые годы после рождения созревания врожденных механизмов, участвующих в координации д
ПСИХОФИЗИОЛОГИЯ ПАМЯТИ
Научение может быть рассмотрено как последовательность сложных процессов, вовлекаемых в приобретение, хранение и воспроизведение информации (см. гл. 15). В результате научения проис
ВРЕМЕННАЯ ОРГАНИЗАЦИЯ ПАМЯТИ
Временная организация следа памяти подразумевает последовательность развития во времени качественно разных процессов, приводящих к фиксации приобретенного опыта. Основные понятия, к
Градиент ретроградной амнезии
Зависимость эффективности модуляции памяти от интервала времени между обучением и применением амнестического агента характеризуетградиент ретроградной амнезии. Град
Стадии фиксации памяти
Гипотеза о двух последовательно развивающихся следах. Согласно гипотезе, формирование энграммы осуществляется в два этапа: первый характеризуется неустойчивой формо
Кратковременная и долговременная память
Развитие концепции временной организации памяти затрудняется нечеткостью и расплывчатостью основных понятий, на которые опирается теоретическая конструкция.
Понятие о кратк
Спонтанное восстановление памяти
Факты о спонтанном восстановлении памяти после действия амнестического электрошока были известны еще в 50-е гг. (см. в [Греченко, 1979]). Сообщалось о восстановлении навыка после пр
Восстановление энграммы действием второго электрошока
Многие исследователи сообщили о восстанавливающем действии вторично предъявленной комбинации «наказание (стимул, который применяется при обучении) – электрошок» (см. в [Doty, 1969])
Восстановление памяти методом напоминания
В работе Р.Дж. Коппенаала и др. [Koppenaal et al., 1967] был впервые применен метод «напоминания». Он заключается в том, что перед тестированием сохранения навыка животным предъявля
Восстановление памяти методом ознакомления
Если перед обучением животное поместить в экспериментальную камеру и дать возможность свободно передвигаться по ней, то после применения амнестического агента ретроградная амнезия н
Ретроградная амнезия для реактивированных следов памяти
Обнаружено, что после перехода памяти в долговременное хранение можно вызвать ретроградную амнезию. В опытах изучали действие электрошока на навык, сформированный несколько дней наз
Основное положение теории активной памяти
Основные положения концепции активной памяти заключаются в следующем.
Память выступает как единое свойство, т.е. не существует разделения на кратковременную и долговременну
ГИПОТЕЗА О РАСПРЕДЕЛЕННОСТИ ЭНГРАММЫ
Опыты с локальными раздражениями мозга показали, что развитие ретроградной амнезии при стимуляции определенной структуры зависит от интервала времени, прошедшего от момента завершен
Распределенность энграммы в опытах с локальными раздражениями мозга
Исследования, выполненные с использованием электрошоков, которые вызывают развитие электрической судорожной активности, показывают сложную динамику перемещений активной энграммы по
Распределенность энграммы по множеству элементов мозга
Представление о том, что след памяти не имеет определенной локализации, а считывается с нейронов разных структур мозга в зависимости от обстоятельств, подтверждено экспериментами. Ф
ПРОЦЕДУРНАЯ И ДЕКЛАРАТИВНАЯ ПАМЯТЬ
В последнее время стало приобретать все большее значение представление о множественности систем памяти. Это представление сформировалось на основе данных, полученных при исследовани
МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПАМЯТИ
В нейронауках исследование механизмов научения и памяти ведется преимущественно в контексте пластичности (см. гл. 15). Именно поэтому многие исследования имели своей целью идентифик
ДИСКРЕТНОСТЬ МНЕМИЧЕСКИХ ПРОЦЕССОВ
Буквы алфавита, атомы и молекулы – все это кодовые обозначения важных сущностей, значение открытий которых нельзя переоценить.
Первым было открытие иероглифов и азбуки. Азб
КОНСТАНТА ЛИВАНОВА
Существует множество эмпирических зависимостей, называемых законами. В качестве примеров можно назвать основной психофизический закон, устанавливающий зависимость силы ощущения от ф
ОБЪЕМ И БЫСТРОДЕЙСТВИЕ ПАМЯТИ
Если следовать логике Д. Гартли, А.А. Ухтомского, Н.Г. Самойлова, М.Н. Ливанова, Г. Уолтера, Е.Р. Джона, К. Прибрама и других сторонников идеи динамического кодирования воспринимаем
ДИАПАЗОН ОЩУЩЕНИЙ
В психофизике дробь Вебера определяет границу между ощущаемым и неощущаемым. Это едва заметный субъективно прирост ощущения, взятый по отношению к исходной величине стимула. Вытекаю
НЕЙРОННЫЕ КОДЫ ПАМЯТИ
Нейронными кодами памяти служат циклически повторяющиеся волны импульсов, порождаемые нейронными ансамблями. Длительность одного цикла составляет около 100 мс. Заметим, что по данны
ЭМОЦИЯ КАК ОТРАЖЕНИЕ АКТУАЛЬНОЙ ПОТРЕБНОСТИ И ВЕРОЯТНОСТИ ЕЕ УДОВЛЕТВОРЕНИЯ
Уильям Джеймс – автор одной из первых физиологических теорий эмоций – дал своей статье, опубликованной более 100 лет тому назад, весьма выразительное название: «Что такое эмоция?» [
СТРУКТУРЫ МОЗГА, РЕАЛИЗУЮЩИЕ ПОДКРЕПЛЯЮЩУЮ, ПЕРЕКЛЮЧАЮЩУЮ, КОМПЕНСАТОРНО-ЗАМЕЩАЮЩУЮ И КОММУНИКАТИВНУЮ ФУНКЦИИ ЭМОЦИЙ
Результаты нейрофизиологических экспериментов показывают, что потребности, мотивации и эмоции имеют различный морфологический субстрат. Так, при стимуляции зон самораздражения латер
ИНДИВИДУАЛЬНЫЕ ОСОБЕННОСТИ ВЗАИМОДЕЙСТВИЯ СТРУКТУР МОЗГА, РЕАЛИЗУЮЩИХ ФУНКЦИИ ЭМОЦИЙ КАК ОСНОВУ ТЕМПЕРАМЕНТОВ
В качестве методов диагностики устойчивых (типологических) индивидуальных особенностей поведения мы использовали две поведенческие модели: тест на предпочтение вероятности, или ценн
ВЛИЯНИЕ ЭМОЦИЙ НА ДЕЯТЕЛЬНОСТЬ И ОБЪЕКТИВНЫЕ МЕТОДЫ КОНТРОЛЯ ЭМОЦИОНАЛЬНОГО СОСТОЯНИЯ ЧЕЛОВЕКА
Сам факт генерирования эмоций в ситуации прагматической неопределенности предопределяет и объясняет их адаптивное компенсаторное значение. Дело в том, что при возникновении эмоциона
ОПРЕДЕЛЕНИЕ ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ
Наиболее часто функциональное состояние (ФС) определяют как фоновую активность нервных центров, при которой и реализуется та или иная конкретная деятельность человека. В классическо
РОЛЬ И МЕСТО ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ В ПОВЕДЕНИИ
Функциональные состояния, регулируемые модулирующей системой мозга – необходимая составляющая любого вида деятельности и поведения. Хорошо изучены отношения между уровнем активации
Стволово-таламо-кортикальная система
Исследования Дж. Моруцци и Г. Мэгуна привели их к открытию в стволе на уровне среднего мозга неспецифической системы, или мезенцефалической ретикулярной формации, активирующей кору
Базальная холинергическая система переднего мозга
Недавно было показано, что магноцеллюлярные нейроны, содержащие АХ и локализованные в базальном переднем мозге (БПМ; Basal Forebrain), моносинаптически проецируются на кору. Они уча
Каудо-таламо-кортикальная система
К регуляции уровня активности организма имеют отношение и базальные ганглии. Другое их название – стриопаллидарная система, которая представляет собой комплекс нейрональных узлов, ц
Модулирующие нейроны
В нервной системе выделена особая группа клеток – модулирующих нейронов, которые сами не вызывают реакции, но регулируют активность других нейронов. Они образуют контакты с другими
ЧТО ТАКОЕ ВНИМАНИЕ
Большое место в жизни древних римлян и греков занимала риторика. Как исскуство и как средство информации она была эффективной только в том случае, если ораторы прекрасно поставленны
ТЕОРИИ ФИЛЬТРА
Первую теоретическую модель внимания, или модель фильтра, создал Д.Е. Бродбент [Broadbent, 1958]. Он предполагал, что нервная система, несмотря на множество входов, в какой-то степе
ПРОБЛЕМА ВНИМАНИЯ В ТРАДИЦИОННОЙ ПСИХОФИЗИОЛОГИИ
Общая идея описанных ранее моделей внимания заключается в том, что на пути прохождения нервных импульсов от рецепторов, подвергшихся воздействию внешних раздражителей, до коры имеет
ПРОБЛЕМА ВНИМАНИЯ В СИСТЕМНОЙ ПСИХОФИЗИОЛОГИИ
Этот парадокс не возникает, если отказаться от рассмотрения поведения как реакции на предъявляемые стимулы. С позиций системной психофизиологии [Швырков, 1995] поведение – это реали
ОРИЕНТИРОВОЧНО-ИССЛЕДОВАТЕЛЬСКАЯ ДЕЯТЕЛЬНОСТЬ
Ориентировочную реакцию (как настройку анализаторов на лучшее восприятие нового стимула) следует отличать от исследовательских реакций и ориентировочно-исследовательского поведения.
ПСИХОФИЗИОЛОГИЯ СОЗНАНИЯ
Сознание представляет собой одно из самых сложных и в то же время загадочных проявлений деятельности мозга. Хотя слово «сознание» достаточно широко используется в повседневной речи
ОСНОВНЫЕ КОНЦЕПЦИИ СОЗНАНИЯ
Говоря о мозговых основах психики и сознания, необходимо сказать, что эти наиболее сложные проявления работы мозга не могут быть объяснены работой какой-то отдельной, изолированной
ПОВТОРНЫЙ ВХОД ВОЗБУЖДЕНИЯ И ИНФОРМАЦИОННЫЙ СИНТЕЗ
Описанная ранее концепция «светлого пятна» исходит из того, что сознание определяется некоторым уровнем возбудимости мозговых структур. Однако можно предположить, что этого недостат
Мозговая основа ощущений
Психологам с 20-х гг. известно, что ощущение возникает довольно поздно – после 100 мс от момента предъявления стимула (т.е. значительно позже прихода сенсорных импульсов в кору). В
Механизмы мышления
Ответу на этот вопрос были посвящены наши исследования последних десяти лет. В работе ставилась задача изучить структуру корковых связей при решении различных мыслительных операций.
СОЗНАНИЕ, ОБЩЕНИЕ И РЕЧЬ
Концепция коммуникативной природы сознания была выдвинута впервые П. В. Симоновым [1981]. Позднее сходные мысли были высказаны другими авторами [Hesslow, 1994; Frith, 1995]. По опре
ФУНКЦИИ СОЗНАНИЯ
Вопрос о функциональном смысле субъективных переживаний, их роли в поведении – одна из важнейших проблем науки о мозге. Представляя собой результат синтеза информации, психические ф
ПОНЯТИЕ БЕССОЗНАТЕЛЬНОГО В ПСИХОФИЗИОЛОГИИ
В процессе деятельности человека в постоянно меняющейся окружающей среде поступающая информация перерабатывается на разных уровнях центральной нервной системы. Переключение нервной
ИНДИКАТОРЫ ОСОЗНАВАЕМОГО И НЕОСОЗНАВАЕМОГО ВОСПРИЯТИЯ
Проблема экспериментального изучения неосознаваемого восприятия сводится к попыткам выявить пороговую разницу между двумя индикаторами: один из них – показатель осознания стимула; д
СЕМАНИЧЕСКОЕ ДИФФЕРЕНЦИРОВАНИЕ НЕОСОЗНАВАЕМЫХ СТИМУЛОВ
Впервые в эксперименте феномен неосознаваемого восприятия словесных стимулов был воспроизведен группой психологов «New Look» [McGinnies, 1944]. У здоровых людей (студентов) порог оп
ВРЕМЕННЫЕ СВЯЗИ (АССОЦИАЦИИ) НА НЕОСОЗНАВАЕМОМ УРОВНЕ
Из наблюдений психиатров известно, что в определенных случаях неосознаваемые внешние сигналы, если они однажды или несколько раз совпадали с сильным отрицательным эмоциональным возб
ФУНКЦИОНАЛЬНАЯ АССИМЕТРИЯ ПОЛУШАРИЙ И БЕССОЗНАТЕЛЬНОЕ
Классические работы Р. Сперри и его коллег на людях с «расщепленным мозгом» [Gazzaniga, LeDoux, 1978] открыли пути для нейропсихологического экспериментального исследования функцион
ОБРАТНЫЕ ВРЕМЕННЫЕ СВЯЗИ И БЕССОЗНАТЕЛЬНОЕ
6.1. Роль обратных временных связей в нервном механизме «психологической защиты»
Одна из форм психологической защиты выражается в повышении
Значение неосознаваемых стимулов обратной связи в когнитивной деятельности
О влиянии неосознаваемых стимулов на когнитивные функции писали многие исследователи [Костандов, 1983; Velmans, 1991], хотя не всегда описывались однозначные результаты. Этот эффект
РОЛЬ БЕССОЗНАТЕЛЬНОГО ПРИ НЕКОТОРЫХ ФОРМАХ ПАТОЛОГИИ
Формированием условного рефлекса на неосознаваемые внешние стимулы объясняется нервный механизм пространственной ориентации слепых людей [Бериташвили, 1969]. Существенная роль звуко
СОН И СНОВИДЕНИЯ
1. АКТИВНОЕ НАСТУПЛЕНИЕ СНА ИЛИ ЛИШЕНИЕ БОДРСТВОВАНИЯ?
Уже в самых ранних исследованиях механизмов сна четко намечаются две основные точки
СТАДИИ МЕДЛЕННОГО СНА И БЫСТРЫЙ СОН
Основные данные, полученные за годы многочисленных и разнообразных исследований сна, сводятся к следующему. Сон – не перерыв в деятельности мозга, это просто иное состояние. Во врем
СОН В ОТНО- И ФИЛОГЕНЕЗЕ
В онтогенезе соотношение «сон–бодрствование» изменяется. Так, у новорожденных состояние бодрствования составляет лишь незначительную часть суток, и значительную часть сна занимает б
ПОТРЕБНОСТЬ В СНЕ
Многие люди хотели бы спать меньше, так как сон, по их мнению, – это потерянное для жизни время. Другие, наоборот, хотели бы спать больше, так как недостаточно хорошо себя чувствуют
ДЕПРИВАЦИЯ СНА
Эксперименты с депривацией (искусственным лишением сна) позволяют предполагать, что организм особенно нуждается в дельта-сне и быстром сне. После длительной депривации сна основным
СНОВИДЕНИЯ
Сновидения издавна поражают и волнуют людей. В древности сновидения рассматривались как «врата в иной мир»; считалось, что через сновидения может происходить контакт с другими мирам
Две парадигмы в исследовании поведения и деятельности
При всем многообразии теорий и подходов, используемых в психологии, психофизиологии и нейронауках, их можно условно разделить на две группы. В первой из групп в качестве основного м
Реактивность
Использование принципа реактивности как объяснительного в научном исследовании базируется на идеях Рене Декарта, изложенных им в первой половине XVII в. Декарт полагал, что организм
Активность
Рассмотрение поведения и деятельности как активности, направленной в будущее, включает понимание активности как принципиального свойства живой материи; конкретная форма проявления а
Эклектика в психологии и психофизиологии
В последнее время представление об активном целенаправленном характере поведения человека и животного становится все более распространенным. Наряду с позитивными последствиями этот
ТЕОРИЯ ФУНКЦИОНАЛЬНЫХ СИСТЕМ
2.1. Что такое система?
Термин «система» обычно применяется для того, чтобы указать на собранность, организованность группы элементов и отг
Временной парадокс
Каким образом результат (событие, которое наступит в будущем) может детерминировать текущую активность, быть ее причиной? Решением этого «временного парадокса» была разработка предс
Целенаправленность поведения
Уже для Аристотеля [1937] была очевидна целенаправленность поведения. Таким образом, идея целенаправленности никак не может считаться новой, хотя в истории можно выделить период, ко
Опережающее отражение
Анализ проблем происхождения и развития жизни с позиций ТФС привел П. К. Анохина [1978] к необходимости введения новой категории:опережающее отражение. Опережающее
Теория П.К. Анохина как целостная система представлений
Итак, первое важнейшее преимущество и признак, отличающий ТФС от других вариантов системного подхода, – введение представления о результате действия в концептуальную схему. Таким об
Системные процессы
В качестве ключевых положений рефлекторной теории П.К. Анохин выделял следующие: а) исключительность пускового стимула как фактора, детерминирующего действие, которое является его п
Поведение как континуум результатов
До сих пор мы с дидактическими целями, а также следуя традиции исходного варианта ТФС, использовали понятие пускового стимула. Однако ясно, что использование этого понятия в рамках
СИСТЕМНАЯ ДЕТЕРМИНАЦИЯ АКТИВНОСТИ НЕЙРОНА
3.1. Парадигма реактивности: нейрон, как и индивид, отвечает на стимул
Как мы уже отмечали, с позиций парадигмы реактивности поведение инди
Активность как субъективное отражение
Рассмотрение соотношения индивида и среды с позиций ТФС уже давно привело к заключению о том, что поведенческий континуум целиком занят процессами организации и реализации функциона
Физические характеристики среды и целенаправленное поведение
Более 30 лет назад Дж. Леттвин и др. [1963], изучив связь активности нейронов сетчатки лягушки с ее поведением, сформулировали в очень яркой форме свое представление о том, что выде
Зависимость активности центральных и периферических нейронов от цели поведения
Убедительные примеры того, как субъективность отражения проявляется в организации активности мозга, можно получить при анализе зависимости от целей поведения активности нейронов «се
Значение эфферентных влияний
Связь активности ганглиозных клеток сетчатки с поведением при закрытых глазах обусловлена уже упоминавшимися эфферентными влияниями. Еще в начале нынешнего столетия С. Рамон-и-Кахал
ПСИХОФИЗИОЛОГИЧЕСКАЯ ПРОБЛЕМА И ЗАДАЧИ СИСТЕМНОЙ ПСИХОФИЗИОЛОГИИ
В настоящем параграфе мы ответим на перечисленные далее вопросы. Как зависят задачи психофизиологии от методологических установок? Существует ли среди них специфическая задача психо
Коррелятивная психофизиология
Традиционные психофизиологические исследования проводятся, как правило, с позиций«коррелятивной (сопоставляющей) психофизиологии». В этих исследованиях психические
Системное решение психофизиологической проблемы
Суть системного решения психофизиологической проблемы заключена в следующем положении. Психические процессы, характеризующие организм и поведенческий акт как целое, и нейрофизиологи
Задачи системной психофизиологии и ее значение для психологии
Использование приведенного решения психофизиологической проблемы в системной психофизиологии в качестве одного из важнейших компонентов методологии позволяет избежать редукционизма
Взаимосодействие коррелятивной и системной психофизиологии
В философии науки утверждается полезность сосуществования альтернативных теорий, способствующего их взаимной критике и ускоряющего развитие науки. Хорошим примером справедливости эт
СИСТЕМОГИНЕЗ
В предыдущем параграфе при формулировке задач системной психофизиологии не случайно на первое место поставлена задача изучения формирования систем. Мы увидим дальше, что история фор
Органогенез и системогенез
В отличие от концепции органогенеза, постулирующей поэтапное развитие отдельных морфологических органов, которые выполняют соответствующие локальные «частные» функции, концепция сис
Научение как реактивация процессов развития
В настоящее время становится общепризнанным, что многие закономерности модификации функциональных и морфологических свойств нейронов, а также регуляции экспрессии генов, лежащие в о
Системная специализация и системоспецифичность нейронов
Специализация нейронов относительно вновь формируемых систем –системная специализация – постоянна, т.е.нейрон системоспецифичен. В настоящее время
Историческая детерминация уровневой организации систем
Представления о закономерностях развития многими авторами разрабатываются в связи с идеями уровневой организации (см. в [Анохин, 1975, 1980; Роговин, 1977; Александров, 1989, 1995,
Структура субъективного мира и субъект поведения
Специализация нейронов относительно элементов индивидуального опыта означает, что в их активности отражается не внешний мир как таковой, а соотношение с ним индивида (см. также пара
Динамика субъективного мира как смена состояний субъекта поведения
С этих позицийдинамика субъективного мира может быть охарактеризована как смена состояний субъекта поведения в ходе развертывания поведенческого континуума (см. рис
Модифицируемость системной организации поведенческого акта в последовательных реализациях
Еще Ф. Бартлетт предлагал полностью отбросить взгляды, в соответствии с которыми «воспроизведение из памяти» рассматривается как «повторное возбуждение неизменных «следов» [Bartlett
Направления исследований в системной психофизиологии
Теоретический и методический аппарат качественного и количественного анализа системных процессов, лежащих в основе формирования и реализации индивидуального опыта в норме и его реор
Зависимость проекции индивидуального опыта от особенностей индивидуального развития
Из всего сказанного ранее с очевидностью следует, что с позиций системной психофизиологии проблема «локализации психических функций» должна быть переформулирована как проблема
Паттерны системной специализации нейронов разных структур мозга
Для того чтобы получить данные, непосредственно характеризующие проекцию тех или иных элементов опыта на структуры мозга, необходимо сопоставить паттерны системной специализации ней
Изменение проекции индивидуального опыта от животного к человеку
В качестве основного критерия эволюционных преобразований мозга рассматривается развитие его коры. Как изменяется проекция опыта на корковые структуры в процессе исторического разви
Изменение проекции индивидуального опыта в условиях патологии
Проекция опыта на структуры мозга может изменяться не только при нормальном ходе индивидуального развития. Повреждающие воздействия, такие, например, как перерезка нервов или ампута
Психофизиологическое основание закона Рибо
Особое значение для рассмотрения изменений проекции опыта в патологии имеет «закон разложения» Т. Рибо [1898]. Т. Рибо выделял «элементы» психики «с точки зрения последовательных фа
Значение материала патологии для изучения системной организации поведения
Таким образом,проекция индивидуального опыта на структуры мозга изменяется в филогенезе, определяется историей обучения в процессе индивидуального
ТРЕБОВАНИЯ К МЕТОДОЛОГИИ СИСТЕМНОГО АНАЛИЗА В ПСИХОЛОГИИ И СИСТЕМНАЯ ПСИХОФИЗИОЛОГИЯ
В заключение рассмотрим, насколько положения ТФС и развитой на ее основе системной психофизиологии отвечают требованиям, которые предъявляются к методологии системного анализа в пси
ПСИХОЛОГИЧЕСКИЕ И БИОЛОГИЧЕСКИЕ ТЕОРИИ НАУЧЕНИЯ
Совершенно очевидно, что закономерности научения изучались задолго до появления современной науки. Необходимость успешно передавать свой опыт следующим поколениям заставляла эмпирич
ПОДХОД К НАУЧЕНИЮ КАК ПРОЦЕССУ
Повышенное внимание к анализу научения как процесса происходит из обыденной задачи научить, передать опыт. Эта процессуальная сторона научения подчеркивается в его определениях как
ПРЕДСТАВЛЕНИЕ О НЕЙРОФИЗИОЛОГИЧЕСКИХ МЕХАНИЗМАХ НАУЧЕНИЯ
С появлением методов регистрации активности мозга и особенно активности нервных клеток проблема научения стала интенсивно исследоваться в нейробиологии. В силу того, что эти экспери
СПЕЦИФИКА ПСИХОФИЗИОЛОГИЧЕСКОГО РАССМОТРЕНИЯ НАУЧЕНИЯ
Психофизиология, в отличие от нейрофизиологии, принимает в расчет субъективное содержание объективных процессов и, более того, изучает как субъективное соотносится с процессами жизн
СИСТЕМНАЯ ПСИХОФИЗИОЛОГИЯ НАУЧЕНИЯ. ПРОБЛЕМА ЭЛЕМЕНТОВ ИНДИВИДУАЛЬНОГО ОПЫТА
В силу того, что, как уже отмечалось ранее, психическое связано с системными процессами организации активности целого мозга [Швырков, 1978], появление в репертуаре индивида нового п
ФИКСАЦИЯ ЭТАПОВ ОБУЧЕНИЯ В ВИДЕ ЭЛЕМЕНТОВ ОПЫТА
Обозначив принципиальный подход к выделению элементов субъективного опыта, теория функциональных систем легла в основу экспериментов по изучению субъективного дробления поведения и
ВЛИЯНИЕ ИСТОРИИ НАУЧЕНИЯ НА СТРУКТУРУ ОПЫТА И ОРГАНИЗАЦИЮ МОЗГОВОЙ АКТИВНОСТИ
Субъективный континуум, как и континуум поведения, представляет собой линейную последовательность сменяющих друг друга состояний, которые соответствуют актам поведения. Эти смены од
Связанные с событиями потенциалы мозга
ССП представляют собой широкий класс электрофизиологических феноменов, которые специальными методами выделяются из «фоновой», или «сырой», электроэнцефалограммы (ЭЭГ). Термин ССП –
Краткая история метода ССП
Связь электрической активности мозга с событиями в окружающей среде и поведении впервые была продемонстрирована и описана англичанином Р.Кейтоном (Richard Caton) в 1875–1887 гг. и н
Общая характеристика сигнала
ССП выделяют при помощи специальных методов из ЭЭГ. Частотный диапазон ССП включает полосу от 0 Гц до 3 кГц и ограничен, с одной стороны, сверхмедленной электрической активностью мо
Стандартные способы получения воспроизводимой конфигурации ССП
Методические требования к регистрации ЭЭГ (установка электродов, выбор системы отведения, полосы пропускания усилителей, способы устранения артефактов) описаны в главе 2. Заметим, ч
Усреднение
В основе выделения ССП из сигнала ЭЭГ лежат следующие допущения:
а) в ситуации многократного повторения события регистрируемый сигнал ЭЭГ (SUMi (t)) является сум
Фильтрация
Случайная, «шумовая» составляющая единичной реализации ССП («сырой» ЭЭГ) может быть устранена посредством сглаживания.
АЛГЕБРАИЧЕСКИЕ ФИЛЬТРЫ
Описание ССП
В результате накопления отрезков ЭЭГ, связанных с определенными событиями, их усреднения, цифровой фильтрации или других процедур, получают ССП – кривую, которую описывают как после
Особенности метода ССП
В последние десятилетия разработаны методы регистрации активности мозга, обладающие значительными исследовательскими возможностями (см. также гл. 2). Однако и при разработке новых м
Зрительные вызванные потенциалы
Зрительные вызванные потенциалы (ЗВП, visual evoked potentials - VEP) [Шагас, 1975; Рутман, 1979; Максимова, 1982; Rockstroh et al., 1982] регистрируются в ситуации предъявления зри
Слуховые вызванные потенциалы.
Слуховые вызванные потенциалы (СВП, auditory evoked potentials – АЕР) [Шагас, 1975; Рутман, 1979; Rockstroh et al., 1982; Hughes, 1985] регистрируются в ситуации предъявления слухов
Соматосенсорные вызванные потенциалы
Соматосенсорные вызванные потенциалы (ССВП; somatosensory evoked potentials, SEP) [Шагас, 1975; Рутман, 1979; Rockstroh et al., 1982] регистрируются в ситуации меха
Потенциалы, связанные с выполнением движений
Потенциалы, связанные с выполнением движений (ПСВД, movement-related potentials – MRP, movement-related brain potentials – MRBP; в русскоязычной литературе использу
Условная негативная волна
Условная негативная волна (УНВ, contingent negative variation – CNV, или волна ожидания, expectancy wave – E-wave). В ситуации предъявления двух стимулов, первый из
Принципы упорядочения феноменологии ССП
Перечень известных типов ССП постоянно пополняется, и нет оснований считать его близким к завершению [Donchin, Isreal, 1980]. Приведем в качестве примеров наиболее известные феномен
ПРОБЛЕМА ФУНКЦИОНАЛЬНОГО ЗНАЧЕНИЯ ССП
В рамках коррелятивной психофизиологии (см. [Швырков, 1995] и гл.14) предполагается, что колебания (компоненты) ССП отражают специфические функции структур мозга, которые реализуют
Психологические корреляты
Поиски психологических коррелятов ССП показали, что: 1) один и тот же ССП связан со многими психологическими процессами (функциями) и 2) одни и те же психические фу
Мозговые источники
Поиски мозговых источников ССП показали следующее: 1) любое колебание ССП, регистрируемое с поверхности головы, представляет собой отражение активности множества корковых и подкорко
ССП КАК ОТРАЖЕНИЕ ДИНАМИКИ ИНДИВИДУАЛЬНОГО ОПЫТА
ССП представляют собой суммарный электрический потенциал различных компонентов ткани мозга, вклад в который вносят нейроны (сома, дендриты и аксоны), глиальные клетки, мембраны клет
Потенциал универсальной конфигурации
Сопоставление ССП, сопровождающих поведение испытуемых в различных экспериментальных ситуациях, показывает, что реализации и смене поведенческого акта соответствует потенциал универ
ПЕРСПЕКТИВЫ ИСПОЛЬЗОВАНИЯ ССП
Эффективность использования ССП как метода психофизиологического исследования определяется решением основной проблемы: каково соотношение параметров ССП, активности мозга, ф
КОНЦЕПЦИЯ СВОЙСТВ НЕРВНОЙ СИСТЕМЫ
Проблемаиндивидуально-психологических различий между людьми всегда рассматривалась в отечественной психологии как одна из фундаментальных. Наибольший вклад в разраб
ОБЩИЕ СВОЙСТВА НЕРВНОЙ СИСТЕМЫ И ЦЕЛОСТНЫЕ ФОРМАЛЬНО-ДИНАМИЧЕСКИЕ ХАРАКТЕРИСТИКИ ИНДИВИДУАЛЬНОСТИ
Для экспериментальной проверки развиваемых представлений о свойствах нервной системы и их психологических проявлениях В.Д. Небылицын провел исследование физиологических основ интелл
ИНТЕГРАЛЬНАЯ ИНДИВИДУАЛЬНОСТЬ И ЕЕ СТРУКТУРА
Подход, который разрабатывался B. C. Мерлиным, составил основу развития оригинальной школы по изучению природы темперамента. Исходные положения, на которых базировался данный подход
ИНДИВИДУАЛЬНЫЕ ОСОБЕННОСТИ ПОВЕДЕНИЯ E ЖИВОТНЫХ
Важная тенденция, которая сформировалась в разработке проблемы природы индивидуально-психологических различий после Б.М. Теплова и В.Д. Небылицына и была основана на модели поведени
ИНТЕГРАЦИЯ ЗНАНИЙ ОБ ИНДИВИДУАЛЬНОСТИ
В конце 80-х гг. с целью формирования новой стратегии исследования природы индивидуально-психологических различий между людьмиВ. М. Русалов разработал методику опросного типа для оц
КРОСС-КУЛЬТУРНЫЕ ИССЛЕДОВАНИЯ ИНДИВИДУАЛЬНОСТИ
В последние годы в дифференциальной психофизиологии начала применяться методология кросс-культурного исследования.
Кросс-культурные дифференциально-психофизиологические исс
ПСИХОФИЗИОЛОГИЯ ПРОФЕССИОНАЛЬНОЙ ДЕЯТЕЛЬНОСТИ
На границах естественных наук и психологии сформировался ряд специальных научных дисциплин и направлений, в том числе психология труда, инженерная психология и эргономика, объектом
ТЕОРЕТИЧЕСКИЕ ОСНОВАНИЯ ПРИМЕНЕНИЯ ПСИХОФИЗИОЛОГИИ ДЛЯ РЕШЕНИЯ ПРАКТИЧЕСКИХ ЗАДАЧ В ПСИХОЛОГИИ ТРУДА
Пониманию необходимости и перспективности изучения психофизиологических процессов в профессиональной деятельности способствуют представления, рассматривающие психическое и физиологи
МЕТОДИЧЕСКОЕ ОБЕСПЕЧЕНИЕ ПСИХОФИЗИОЛОГИЧЕСКОГО АСПЕКТА ПРИКЛАДНЫХ ИССЛЕДОВАНИЙ
В прикладных исследованиях комплексный характер психофизиологических исследований обеспечивается применениемполиэффекторного метода, включающего регистрацию частоты
ПСИХОФИЗИОЛОГИЯ ПРОФЕССИОНАЛЬНОГО ОТБОРА И ПРОФПРИГОДНОСТИ
Применение психофизиологических методов в психологии труда было вызвано необходимостью разработки объективных и количественных критериев психофизиологического отбора, которая как пр
ПСИХОФИЗИОЛОГИЧЕСКИЕ КОМПОНЕНТЫ РАБОТОСПОСОБНОСТИ
Способность человека к выполнению конкретной деятельности в рамках заданных временных лимитов и параметров эффективности определяет содержание работоспособности как основной составл
ПСИХОФИЗИОЛОГИЧЕСКИЕ ДЕТЕРМИНАТЫ АДАПТАЦИИ ЧЕЛОВЕКА К ЭКСТРЕМАЛЬНЫМ УСЛОВИЯМ ДЕЯТЕЛЬНОСТИ
В настоящее время основными направлениями в изучении адаптации стали определение этапов становления психофизиологической системы адаптации, критериев ее сформированности, выделение
ПСИХОФИЗИОЛОГИЧЕСКИЕ ФУНКЦИОНАЛЬНЫЕ СОСТОЯНИЯ (ПФС)
Актуальность изучения ПФС определяется их вкладом в обеспечение эффективности деятельности и надежности человека, а также увеличением количества профессий и изменением условий труда
БИОЛОГИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ (БОС)
Интерес к исследованиямбиологической обратной связи (БОС) в целях произвольного управления ФС на основе объективной информации о динамике психофизиологических показ
ПСИХОФИЗИОЛОГИЧЕСКИЙ АНАЛИЗ СОДЕРЖАНИЯ ПРОФЕССИОНАЛЬНОЙ ДЕЯТЕЛЬНОСТИ
Психофизиологический анализ профессиональной деятельности предполагает «рассмотрение ее как сложного, многомерного и многоуровневого, динамического и развивающегося явления» [Ломов,
СРАВНИТЕЛЬНАЯ ПСИХОФИЗИОЛОГИЯ
Сравнительная психофизиология – наука, нацеленная на установление закономерностей и выявление различий в структурной и функциональной организации мозга, поведении и психике у животн
ПОЯВЛЕНИЕ ПСИХИЧЕСКОГО
Наиболее принятой в настоящее время точкой зрения является та, что психическое – атрибут живых систем, а неживая природа, включая созданные людьми сложные устройства, психикой не об
ЭВОЛЮЦИЯ ВИДОВ
Современные данные об эволюции животных указывают на расхождение линий эволюции и развитие параллельных линий (рис. 19.2 А), в том числе и среди млекопитающих (рис. 19.2 Б), и среди
ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА
Строение мозга у животных разных видов разнообразно. И хотя, как следует из рис. 19.3, родственные виды, например среди ракообразных или млекопитающих, имеют общие черты в строении
СРАВНИТЕЛЬНЫЙ МЕТОД В СИСТЕМНОЙ ПСИХОФИЗИОЛОГИИ
Системная психофизиология, основы которой были заложены трудами В.Б. Швыркова и его коллег, основана на признании: 1) единой психофизиологической реальности, в которой психологическ







Новости и инфо для студентов