ПРОБЛЕМА ПРОИСХОЖДЕНИЯ И СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА
 ПРОБЛЕМА ПРОИСХОЖДЕНИЯ И СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА.
ПРОБЛЕМА ПРОИСХОЖДЕНИЯ И СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА.
Стереохимическая гипотеза. Области диалектов и области двусмысленности. Протокод и 22 аминокислоты.

mailto:azad-kerimbekov@yandex.ru
Генетический код (ГК) к настоящему времени достаточно хорошо изучен, однако его происхождение и свойства либо не объяснены вообще, либо объяснены неубедительно и остаются загадкой. Единственность, как центральная нерешенная проблема (1), триплетность, универсальность, неперекрываемость, наличие диалектов и, наконец, число нуклеотидов «4» и число аминокислот «20», неоднородная вырожденность кода –всё это требует конкретных и четких решений. В этой статье предпринята попытка ответить на ряд вопросов, вытекающих из таблицы соответствия «кодон – аминокислота (АК)» - из её современной версии. Подобных попыток в литературе много и число их в последнее время увеличивается. По последней статье Victor A..Gusev & Dirk Schulze- Makuch (1) можно ознакомиться с обширным списком публикаций о ГК и с критическим анализом наиболее значимых работ. Авторы (1) обосновывают принцип детерминизма в вопросах происхождения и свойств ГК за рамками стереотипов стерехимической, эволюционной и гипотезы случайного замораживания - frozen-accident.
Ими так же сформулирована гипотеза о первичности: «Первичным началом жизни является сам процесс органического синтеза». Это полностью созвучно с моей идеей о роли серина, как ключевой молекулы на начальном этапе химической эволюции ( http://gypoteza.narod.ru ). Нет необходимости в обсуждении вышеупомянутой (1) публикации – работа вполне самодостаточна.
Первичность синтеза обуславливает протекание ряда реакций по параллельным каналам с «перекличкой» между стадиями и промежуточными результатами. В последнее время не подвергается сомнению образование в модифицированных экспериментах по схеме Миллера простейших сахаров и азотистых циклов ( аденин и гуанин ), не говоря уже об аминокислотах. Это детерминировано (1) спецификой атомов и молекул участников. В простейшем случае биоорганических молекул – аминокислот - неминуемо образуется α-спираль или β-лист с последующей компактизацией по схеме спираль – клубок. Любое другое стерически, термодинамически или как-угодно не выгодно, или запрещено. Да и есть-ли это «другое» ?
Серьезной химической проблемой становится выявление и обоснование тех самых параллельных реакций, которые в результате привели к образованию компонентов, необходимых для самосборки нуклеиновых кислот. В качестве гипотетического можно высказать предположение о первоначальном образовании бимолекулярных ассоциатов комплементарных азотистых оснований ( A---U(T), C---G), возникновение реакционных центров на противоположных водородным связям концах и последующей сшивкой с полисахаро-фосфатными цепочками. Вероятным итогом такого типа реакций становятся нитевидные и спиральные структуры – РНК и ДНК. Часть этой проблемы затрагивается на страницах «gypoteza».
При анализе параметров и свойств генетического кода приходится исходить из детерминированной структуры ДНК ( пока нет альтернативного варианта ) и из определения генетического кода. ГК – система (правила) зашифровки (и перевода!), реализованная в виде последовательности нуклеотидов.
Особую значимость имеет идея неизбежной триплетности четырехбуквенного ГК, но это тезис дискуссионный. Триплетность можно считать следствием уникального сочетания способов хранения и чтения генетической информации, а можно – детерминированным параметром ГК. В. Гусев так резюмирует работы Aldana: «Данные, приведенные в работах (Aldana, Cazarez-Bush & al. 1998; Aldana-Gonzales, Cocho & al. 2003 ) (2) следует интепретировать так, что триплетная структура генетического кода детерминирована физикой взаимодействия мономеров уже на ранних этапах формирования гетерополимерных матриц. Другими словами, селекция структуры кодонов проходила на Земле в период химической, но не биологической эволюции».
Предварительно немного об особенностях полинуклеотидных последовательностей. Самосборка нуклеиновых кислот приводит к оптимальной геометрии с пространственной регулярностью лишь в случае непрерывного чередования азотистых оснований ( у прокариот и созревших m-РНК это смысловые последовательности ) с обязательным соблюдением принципа комплементарности: A – U(T), C – G. Появление ошибок, пропусков, несоответствий ослабит спираль или цепочку и вызовет либо случайные, либо запрограммированные автокаталитические рекомбинационные процессы - сплайсинг (для РНК они подтверждены экспериментально, например в работах Томас Р. Чек, (3)). Гибридизация полимерных цепочек нуклеотидов в двойные спирали ДНК требует отдельного серьезного рассмотрения и выходит за рамки этой работы. Дело осложняется тем, что ДНК имеет мозаичную структуру с чередующимися экзонными и интронными фрагментами, с повторами отдельных участков гена, тандемными повторами и говорить о «кодон – АК» соответствии удобнее на примере m-РНК с вырезанными интронами, т.е. на примере сформировавшейся последовательности кодонов. Но и «бестолковая» сборка ДНК по принципу «как бы чего другого не получилось» может иметь серьёзную подоплёку. Как отмечает Л.А. Грибов (6) «…жизнь возможна только тогда, когда это приемно-преобразующее устройство способно работать в условиях нечеткой информации».
Так вот у эукариот признак белка (кодирующая последовательность) случайным образом размазанный по спирали и есть как раз нечёткая информация. m-РНК выполняет роль внучки, которая целесообразно выбирает из бабушкиного сундука (ДНК) нужное! Налицо прототип одного из свойств живого эволюционирующего организма.
В типичной В-форме ДНК считывание и синтез м-РНК будет происходить по большой бороздке: РНК-полимераза, двигаясь по ДНК, (Рис. 5) обеспечит процессы синтеза молекул РНК – малой и матричной. В большую бороздку обращены характерные группы азотистых оснований: NH2, CH3, C=O, уникальное пространственное расположение которых и определяет индивидуальность кодонов. ( По малой бороздке с ДНК взаимодействует другой белок, что приводит к значительным конформационным изменениям молекулы – изгибу до 100 градусов). При шаге спирали 3.4 нм и расстоянии между линиями спаривания (водородными связями) 0.34 нм на один виток спирали приходится 9 пар нуклеотидов, что соответствует трем кодонам. (Один кодон в «просвете» - наиболее доступный кодон), (Рис. 2). Это суждение служит основанием для перевода фактора «триплетность» из категории аргументов в категорию функций и позволяет объяснить некоторые свойства ГК. Детерминированным, скорее всего, будет соответствие «АК – кодон», как следствие стереохимических и физико-химических особенностей АК и азотистых оснований, а триплетность окажется функцией.
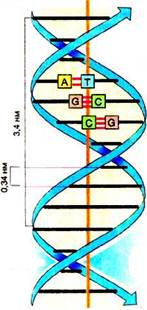
Рис. 1 Рис. 2
Фрагмент двойной спирали (ДНК). В центре рисунка 2 (помечены белыми квадратиками) 3 линейных открытых со стороны большой бороздки сцепки комплементарных оснований и 2 сцепки в просвете малой бороздки. Рисунок 1 – схема ДНК с размерами из книги Кольмана и Рёма « Наглядная биохимия» (4).
В статье Гусева со ссылкой на Aldana-Gonzales, Cocho (2) реферируется исследование авторов: « На основании решения уравнения Ньютона при движении олигомера по матрице, авторы показали, что время его пребывания в минимуме потенциала возрастает в 2-3 раза по сравнению с промежуточным состоянием. По мнению авторов, это могло послужить предпосылкой для формирования триплетной структуры генетического кода». Более того, этот результат позволяет надеяться на справедливость следующего предположения: каждый следующий акт считывания начнется лишь по завершении предыдущего как следствие последовательности считывания и времени пребывания в контакте с кодоном (времени синтеза одного нуклеотида РНК). Из этого с учетом сказанного выше следует, в частности, неперекрываемость ГК.

Рисунок 5 из работы (12) адаптирован к тексту.
Таким образом, триплетность и неперекрываемость можно считать следствиями действующих при спиралеобразовании стереохимических законов конформационных соответствий и кинетических характеристик транскрипции.
Выбор четырех букв – A, U(T), C, G пока ничем не обоснован и предыдущий вывод - молчаливое допущение априорного существования именно этих азотистых оснований. Они что? – детерминированы? Они неизбежно образуются на начальной стадии молекулярной эволюции? Или только ОНИ выбираются и вовлекаются в процесс конструирования ДНК? Роль интронов (об этом подробнее сказано выше ) проявляется в сплайсинге РНК (3) и при описании свойств ГК может не учитываться. Отмечу ещё раз, что проблему свойств ГК
можно вообще рассматривать, анализируя последовательность кодонов в m-РНК и этого вполне достаточно. Иначе мы входим в паутину причинно-следственных связей, не имеющих прямого отношения к решаемым вопросам.
В работе (1) делается смелое обобщение о ГК как фундаментальном законе природы и о детерминированности как четырех нуклеотидов, так и 20-и аминокислот. Вообще говоря, детерминированное сложное есть функция детерминированного простого и «20» аминокислот», по-видимому, следствие аргумента «4 нуклеотида». Постулируемая фундаментальность ГК фактически пропускает целую эру химической эволюции. Возможно, лучше говорить о детерминированном соответствии кодонов и АК, списывая формирование этих соответствий на законы стереохимии ( они-то детерминированы!). А вот стоит только уйти от этого условия – условия соответствия «Кодон - АК», как сразу возникает проблема множественности ГК.
В связи с проблемой единственности ГК рассмотрим возможные варианты формирования двойной спирали при различном числе нуклеотидов. Нечетное число не приведет к комплементарному спариванию и, следовательно, к образованию регулярной спиральной структуры. При двух нуклеотидах более прочная структура соответствует комплементарной паре ( 2 или 3 водородные связи ), но при этом может быть закодировано только 8 аминокислот, что противоречит АК-составу белков. 4 нуклеотида при триплетном кодировании обеспечат соответствие кодонов 64-м информационным позициям и это, в свою очередь, обеспечивает синтез наблюдаемых белков, но приводит к вырожденности кода. 6 нуклеотидов – 216 информационных позиций и вполне «хорошая» двойная спираль. Но так-ли это? У природы, повидимому, не оказалось ещё одной комплементарной пары азотистых оснований (нуклеотидов) способных участвовать в формировании двойной спирали.
Только 4 основания, а не 3 и не 5 ( и не 6 ? ) способны нормализовать геометрию двойной спирали. Скорее всего, дело не в чрезмерной избыточности – вырожденности – отброшенной природой, а в стереохимических несоответствиях и логических противоречиях возникших бы в таблице соответствия «кодон – АК», то есть, в жизни. Ниже будет подробное обсуждение числа кодируемых аминокислот, а пока, считая число «4» детерминированным, продолжим. Есть три варианта формирования последовательностей нуклеотидов в РНК и ДНК:
1. 1. «Сборка» совершенно случайной последовательности.
2. 2. «Сборка» с той или иной степенью регулярности, периодичности.
3. 3. «Сборка» по где-то записанным правилам – целевая сборка.
Вариант «2» приводит к варианту «1», т.к. в выбранном повторяющемся фрагменте последовательность случайна. Если не случайна, то результат тривиален – кодируется ограниченное число полипептидов. Вариант «3» - это уже мистика или, в лучшем случае, замкнутый круг. Кто и где записал эту программу?
Вариант «1» - формирование случайных последовательностей нуклеотидов – приводит к многообразию полипептидов и выглядит в свете последующей эволюции наиболее приемлемо.
Здесь, между прочим, уместно обратить внимание на корелляцию встречаемости и вырожденности АК. Сопоставим встречаемость (5) со средней вырожденностью из таблицы соответствия графически (Рис. 3):


Рисунок 3. Рисунок 4.
Симбатная зависимость (Рис. 3) четко устанавливается для всех АК, за исключением Аргинина (Arg) и Триптофана (Trp). Повидимому для них работает иной принцип распознавания. У Trp радикал конформационно подобен аденину (А), а у Arg фрагмент (2(NH2))+ отделен от остова АК цепочкой 3(СН2) и почти «самостоятелен». На этом основании можно предположить, что и Arg, и Trp кодируются соответствующими триплетами по своим радикалам, а не только как аминокислоты. Обозначенный синим квадратиком Met имеет вырожденность 0.5, поскольку у него общий кодон (AUG) с сигналом «start».
Для удобства при последующим исследовании ГК приведена таблица соответствия на языке m-РНК – ключ к генетическому коду. Для некодирующей спирали ДНК U заменяется на Т.
UUU Фенилаланин UCU Серин UAU Тирозин UGU Цистеин
UUC - « - UCC - « - UAC - « - UGC - « -
UUA Лейцин UCA - « - UAA stop UGA stop
UUG - « - UCG - « - UAG - « - UGG Триптофан
CUU - « - CCU Пролин CAU Гистидин CGU Аргинин
CUC - « - CCC - « - CAC - « - CGC - « -
CUA - « - CCA - « - CAA Глутамин CGA - « -
CUG - « - CCG - « - CAG - « - CGG - « -
AUU Изолейцин ACU Треонин AAU Аспарагин AGU Серин
AUC - « - ACC - « - AAC - « - AGC - « -
AUA - « - ACA - « - AAA Лизин AGA Аргинин
AUG Метионин, start ACG - « - AAG - « - AGG - « -
GUU Валин GCU Аланин GAU Аспарагиновая GGU Глицин
GUA - « - GCA - « - GAA Глутаминовая GGA - « - GUG - « - GCG - « - GAG кислота GGG - « - Предварительно рассмотрим появление диалектов в существующей версии ГК. Диалект – нарушение стандартного соответствия…Ещё раз вернёмся к проблеме выбранного природой числа АК, используя свойства гипотетического протокода. Для наглядности представим графически зависимость числа вакантных кодонов от используемого числа АК при средней вырожденности 2.5 (Рис. 4). Если, например, выбрано 10 АК, то число «занятых» кодонов будет 10*2.5 = 25, а число вакантных 64 – 25 = 39 и так далее. Из графика непосредственно следует: если число АК (при заданной средей вырожденности) меньше оптимального, то механизм кодирования приводит к результатам с многими диалектами – область диалектов; если же число АК больше оптимального, то мы попадаем в область двусмысленности. Оптимальное число кодируемых АК, а именно 22 – рубеж между областью множественности кодов и областью двусмысленности. И то и другое привело бы к невообразимой путанице в последующих шагах эволюции и было постепенно отвергнуто природой.
Обобщим проведенное рассмотрение ГК в следующих выводахж:
1. 1. Единственность, триплетность и неперекрываемость ГК следуют из детерминированного алгоритма «сборки» нуклеиновых кислот.
2. 2. Установленное число протеиногенных АК – 20, а, возможно, и 22 есть следствие детерминированного числа азотистых оснований «4». Это оптимальное число АК определяется границей между областью множественности ГК и областью его двусмысленности.
3. 3. Признаком возникновения диалекта ГК служит сочетание одинаковости двух первых букв кодона и принадлежности третьих букв некомплементарным основаниям.
4. 4. Степень вырождения по конкретным АК не установлена, но, повидимому, определяется особенностями структурных и физико-химических соответствий « АК – кодон».
5. 5. Универсальность ГК непосредственно вытекает из единственности (уникальности) стереохимических особенностей ДНК, детерминированных предшествующими шагами эволюции.
6. 6. Предположительно существует некий более совершенный ПРОТОКОД с вырожденностью не более четырёх, описывающий 22 аминокислоты, два сигнала терминации и вспомогательное сигнальное слово. Действующая версия ГК - наиболее распространенный диалект протокода.
P.S. В заключение нельзя не отметить некоторые неоправданные подходы при исследовании свойств генетического кода, особенно множество всевозможных заморочек - кубы, додекаэдры, ромбы, круги, магические числа и прчее, подобное. Что ж, они неизбежны, и делают честь авторам высшего пилотажа, но не содержат ответов на поставленные вопросы. Зачастую происходит неоправданное усложнение, хорошо воспринимаемое лишь самими авторами. Вообще-то, в любой конструкции можно отыскать совершенно неожиданные закономерности…( Возьмем пирамиду Хеопса… Там столько !!! ) Как правило закономерности, вытекающие из параметров, функциональные и не имеют обратного действия. А заморочки и от Истины увести могут – на то они и заморочки.
Литература:1. Victor A.Gusev & Dirk Schulze- Makuch,
Physics of Life Reviews, v.1, dec. 2004, 202 – 229.
2. Aldana-Gonzales, Cocho & al. J. theor. Biol., 220, 2003, 27 –
3. Томас Р. Чек Scient. American, 1987, № 1
4. Рисунок из сайта http://biochemistry.vov.ru/nagl_bio/90.htm
5. Сайт http://jabad.narod.ru/002/004/0001.htm
6. http://www.gordon.ru/konkurssite/grla.html
7. http://researchnews.osu.edu/archive/aminoacd.htm
8. Таблица из электронной публикации Г.М.Дымшица
«Введение в молекулярную биологию».
9. http://www.osu.edu/researchnews/archive/aminoacd.htm
10. http://www.computerra.ru/offline/2001/413/12786/
11. http://macroevolution.narod.ru/markov_kulikov.htm
12. http://orgchem1.city.tomsk.net/nucleic/express/express.htm
13. Анимация из сайта «textronica»