рефераты конспекты курсовые дипломные лекции шпоры

- Раздел Биология
- /
- Фаг λ.
Реферат Курсовая Конспект
Фаг λ.
Фаг λ. - раздел Биология, Образование РНК-затравки на о.ц. ДНК-матрице Особенность Репликации Фага В Том, Что Она Начинается По Схеме Кернса, А Зака...
Особенность репликации фага в том, что она начинается по схеме Кернса, а заканчивается по схеме разматывающегося рулона. Геном представлен линейной д.ц. ДНК длиной 48502 п.н., с липкими концами длиной 12 нт. Внутри инфицированной клетки ДНК циркуляризуется и сверхспирализуется с помощью клеточных белков. Кольцевые молекулы образуются и при выщеплении профага.
|
Структура ori.
 | | | | ||||||
 | |||||||||
|
Ori расположен внутри гена O, кодирующего ori-связывающий белок. Состоит из четырех палиндромов (их еще называют итеронами), к которым справа примыкает А/Т богатый участок.
Инициация репликации. Ori узнается вирусным белком – О-белком (продукт гена О). О-белок тетрамеризуется на ori и образует комплекс с ДНК, называемый О-сомой (комплекс виден в электронный микроскоп). В О-соме ДНК немного подплавляется и в комплекс привлекается клеточная хеликаза DnaB в комплексе с вирусным белком Р (аналог клеточного DnaC), который угнетает ее активность, а также способен взаимодействовать с О-белком.

Таким образом белок О привлекает хеликазу к ori, но она неактивна. В активации хеликазы участвуют клеточные белки – шапероны:DnaJ, GrpE, DnaK (комплекс аналогичен Hsp70 эукариот), которые вызывают диссоциацию белка Р из комплекса.
DnaB начинает расплетать ДНК. На одноцепочечные участки ДНК садится праймаза и начинает синтезировать РНК-затравку. Если в комплексе нет GrpE, то реплицируется только одна цепь (односторонняя (вправо) репликация). Для инициации репликации требуется транскрипция. Считается, что инициации репликации способствуют конформационные изменения, происходящие при транскрипции. При слабой интенсивности транскрипции происходит одностронняя репликация.
In vitro для репликации транскрипция не нужна, но если добавить белок E.coli – HU, то без транскрипции репликация не начинается.
Элонгация репликации. Осуществляется высокопроцессивной ДНК полимеразой III E.coli. Фермент состоит из нескольких субъединиц и представляет собой гетеродимер. Состоит из трех групп белков.
1)соre: α2-каталитическая (синтез ДНК 3’-5), ε2-экзонуклеазная (3’-5’, proofreading activity)
2)β2-фактор процессивности полимеразы (аналог PCNA)
3)τ2, γ2,δ2,δ’2, χ, ψ – clamp loader (загружает фактор процессивности на ДНК с затратой АТФ). τ в нужный момент способствует раскрыванию кольца β-субъединицы. δ,γ,δ’ – «гаечный ключ» раскрывает β2-фактор. Фактор процессивности снимается, когда полимераза сталкивается с затравкой.
Удаление РНК-затравок.
Затравку удаляют РНКазаН и ДНК полимераза I (обладает 5’-3’ экзонуклеазной активностью и способна удалять последний рибонуклеотид затравки, связанный с дезоксирибонуклеотидом). ДНК-полимераза III застраивает брешь.
Переход механизма репликации от схемы Кернса в схему разматывающего рулона.

 Возможны несколько путей перехода.
Возможны несколько путей перехода.
1.Односторонняя репликация.
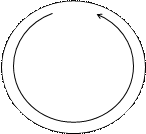 |  | ||||
 | |||||
Гипотеза: превращение θ – σ, связано с ослаблением траскрипции в участке ori и, как следствие, возникновением односторонней репликации.
Причина ослабления транскрипции: а)действие cro-репрессора;
б) в клетке присутствует ограниченное количество белка DnaA, который влияет на транскрипцию с правого промотора фага. При увеличении количества фаговой ДНК, он вытитровывается и транскрипция ослабляется.
Если бы это был единственный механизм, то конец был бы фиксированным (как у φХ174). Но концы молекул могут разными.
2.Рекомбинационный механизм.
Фаг обладает red α,β,γ – системой: redα – 5’-3’экзонуклеаза, redβ – аналог SSB, redγ – ингибитор RecBCD системы (содержит экзонуклеазу), разрушающей линейные молекулы, а также σ-молекулы.
Redα образует агрессивный одноцепочечный 3’-конец, который внедряется в дуплекс кольцевой молекулы и вытесняет небольшой участок.
|
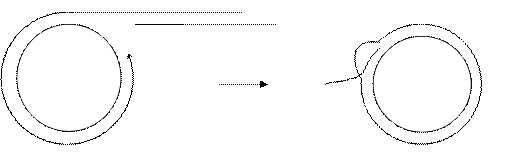
3.Внесение разрыва.
В принципе возможен случайный одноцепочечный разрыв.
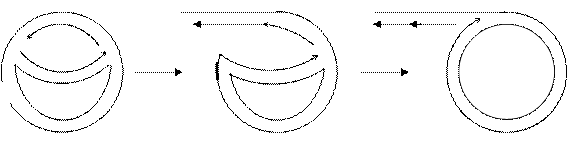
Механизмы не исключают друг друга и в принципе возможны.
Rep-хеликаза не участвует, поскольку белок О обладает хеликазной активностью.
В итоге образуются конкатемерные молекулы, которые надо разрезать на мономеры так, чтобы образовались липкие концы длиной 12 нуклеотидов. Нарезание осуществляется вирусной терминазой и сопряжено с упаковкой ДНК в вирусные частицы. Терминаза состоит из трех субъединиц А(Nu1)2 (А, потому что крайний слева в геноме). Разрезание происходит по специфическим участкам – cos.
Структура cos.
– Конец работы –
Эта тема принадлежит разделу:
Образование РНК-затравки на о.ц. ДНК-матрице
Геном представлен двуцепочечной кольцевой ДНК В геноме закодирован белок большой Т Ag кДа Он единственный вирусный белок участвующий в... способен взаимодействовать с рядом TF переход клетки в S фазу... сам является TF синтез вирусных белков...
Если Вам нужно дополнительный материал на эту тему, или Вы не нашли то, что искали, рекомендуем воспользоваться поиском по нашей базе работ: Фаг λ.
Что будем делать с полученным материалом:
Если этот материал оказался полезным ля Вас, Вы можете сохранить его на свою страничку в социальных сетях:
| Твитнуть |
Хотите получать на электронную почту самые свежие новости?

Подпишитесь на Нашу рассылку
Реклама





Информация в виде рефератов, конспектов, лекций, курсовых и дипломных работ имеют своего автора, которому принадлежат права. Поэтому, прежде чем использовать какую либо информацию с этого сайта, убедитесь, что этим Вы не нарушаете чье либо право.
© copyright 1999 - 2024 allRefs.net. Все права защищены. Страница сгенерирована за: 0.017 сек.
Новости и инфо для студентов