Анатомия и гистология сельскохозяйственных животных
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ ФЕДЕРАЦИИ
ФГОУ ВПО «БЕЛГОРОДСКАЯ ГОСУДАРСТВЕННАЯ
СЕЛЬСКОХОЗЯЙСТВЕННАЯ АКАДЕМИЯ»
Ю.Н.Литвинов
Анатомия
и гистология
сельскохозяйственных
животных
Краткий курс лекций
Тема 1. Введение
1. Анатомия, цель изучения и место дисциплины в системе подготовки инженеров-технологов мясной промышленности.
2. Краткие исторические сведения о развитии анатомии.
3. Понятие о клетках, тканях, системах органов и организме.
Вопрос 1.Анатомия, цель изучения и место дисциплины в системе подготовки инженеров-технологов мясной промышленности
Анатомия и гистология сельскохозяйственных животных является первой фундаментальной дисциплиной, от усвоения которой зависит успех изучения всех последующих специальных дисциплин. Изучение макро- и микроморфологии животных способствует развитию широкого биологического кругозора. Дисциплина включает два самостоятельных курса - "Анатомия" и «Гистология». Цель курсов: "Анатомия" - формирование у студентов знаний основ атомического строения животных; "Гистология" - формирование у студентов знаний микроскопического строения органов и тканей.
Цель преподавания дисциплины - дать знания строения и процессов жизнедеятельности здорового организма животного, его систем и органов, закономерностей взаимодействия с окружающей средой, формирования и повышения продуктивности сельскохозяйственных животных, основы животноводства.
Задача изучения дисциплины - приобретение знаний строения и жизненных функций организма животного, обеспечивающих нормальную деятельность всех органов и систем.
Вопрос 2.Краткие исторические сведения о развитии анатомии
Анатомия относится к одной из древнейших наук, развитие которой находилось в тесной зависимости от развития человеческого общества и его потребностей. В древнем мире первобытные люди, охотясь на животных и разделывая добычу, познавали видовые особенности тела и внутренних органов, определяли их месторасположение и значение. С одомашниванием животных (около 8 тыс. лет назад) морфо-анатомические знания в значительной степени расширялись и углублялись, но они долгое время оставались недостаточными и несистематизированными.
Первые обобщения накопленных знаний принято связывать с именами Алкмеона, Демокрита (живших около 500 лет до н.э.) и Гиппократа (460-377 гг. до н.э). Именно они первые предпринимали анатомические исследования трупов людей и вскрытие животных на скотобойнях.
Большой интерес к познанию видовых особенностей строения тела человека и животных был проявлен в трудах Аристотеля (384-322 гг. до н.э). Аристотель изучал также развитие цыпленка. Своими трудами Аристотель заложил основы сравнительной анатомии и эмбриологии.
До начала новой эры продолжалось накопление знаний в области анатомии и морфологии. Герофилом и Эразистром были описаны легочная артерия, клапаны сердца, оболочки и желудочки мозга, строение глазного яблока, и даже высказано предположение о наличии мельчайших канальцев между артериальными и венозными сосудами.
В начале новой эры (30-54 гг., расцвет римской империи) А. Цельс систематизировал накопленные знания древнегреческих ученых. Эта работа была продолжена К. Галеном, (130-210 гг.) который значительно расширил анатомические познания собственными наблюдениями (особенно в области нервной системы). Гален описал строение стенок желудка, кишечника, кровеносных сосудов, матки. Вместе с тем, Гален ошибочно считал, что в артериях находится не кровь, а воздух, центром кровообращения является не сердце, а печень.
В раннее средневековье (III-VI века) войны и переселения привели к упадку научной мысли. VII-VIII века характеризуются вторжением в Европу арабских народов, что способствовало ассимиляции накопленных культурных и научных знаний и сохранению научного наследия античного мира (книги древних народов переводились на арабский язык).
Особенно велика в этом заслуга крупнейшего ученого таджикского врача Авиценны (Абу-Али-Ибн-Сина, 980-1037 гг.), который в своем труде "Канон медицины" не только обобщил сведения трудов Гиппократа, Аристотеля, Галена, но и расширил их собственными наблюдениями. Он отметил благотворное влияние на организм правильного питания, чистого воздуха, солнечного света, доказал влияние нервной системы на все функции организма.
С середины XV века зарождается капитализм, получает дальнейшее развитие животноводство, что способствовало повышению интереса к зооветеринарным наукам. Начинается эпоха "Возрождения". Наиболее выдающиеся ученые этой эпохи в области анатомии - итальянский художник и мыслитель Леонардо да Винчи (1452-1519) и Андрей Везалий (1514-1564). Андрей Везалий обобщил накопленный материал в капитальном руководстве "О строении человеческого тела в семи книгах".
Во второй половине XVII века появляется ряд исследователей, внесших огромный вклад в изучение строения тела человека и животных.
Б. Евстахий (ок.1510-1574) описал строение почек, зубов, органа слуха и др.
Г. Фаллопий (1523-1562) описал сосуды плода, яйцеводы, желчевыводящие пути.
И. Фабриций (1537-1619) описал венозные клапаны, особенности развития плодов и плаценты у домашних животных.
К. Руини (1530-1598) в 1598 году выпустил капитальное руководство по анатомии и лечению лошади.
В 1628 г. Вильям Гарвей (1578-1657), экспериментируя на животных, впервые открыл большой круг кровообращения и тем самым заложил научные основы физиологии. Ему принадлежит и обширный труд по эмбриологии животных (1633 г.).
В 1677 году Антоний Левенгук (1632-1723) создал микроскоп с 300 кратным увеличением и тем самым расширил возможности изучения анатомических и гистологических структур.
Справка: в России первый микроскоп изобретен при Петре I И.И. Беляевым и И.П. Кулибиным.
Начинается эра бурного развития новых разделов анатомии - физиологии, эмбриологии, микроскопической анатомии. М. Мальпиги (1628-1694), используя микроскоп, показал, что артерии и вены соединяются между собой мельчайшими сосудами - капиллярами. Р. Декарт (1596-1650) впервые открыл рефлексы, но считал их механическими явлениями.
Большую роль в изучении анатомии и гистологии внесли и ученые последующих поколений – анатомы К. Вольф, М.М. Тереховский, К.М.Бэр, А.О. Ковалевский, А.Н. Северцов, Л.А. Третьяков, А.Ф.Климов, А.И. Акаевский и др., гистологи - А.М Шумлянский, К. Биша, И.И. Мечников, А.А Заварзин, П.П. Иванов, П.А. Ковальский и др.
Вопрос 3.Понятие о клетках, тканях, системах органов и организме
Все высшие организмы состоят из микроскопических элементов – клеток и неклеточныхструктур. Наука о строении, функциях и развитии клеток называется – цитология (от лат. cytos – клетка; logos – учение).
Началом эры цитологии принято считать время открытия клетки (ткани пробки) Робертом Гуком (1665 г.). Позднее клетки животных увидел Антоний Левенгук (клетки крови), а первые специальные исследования животных клеток провел итальянец М. Мальпиги.
В результате огромного количества микроскопический исследований в 17–18 вв. накопился богатый фактический материал о строении растительной и животной клетки. Это позволило Т. Шванну (в 1839) обобщить накопленные данные и в своей знаменитой книге «Микроскопические исследования о соответствии в структуре и росте животных и растений» сформулировать клеточную теорию. Она явилась величайшим открытием в биологии, так как показала, что все растения и животные организмы построены и развиваются по единому плану. Клетка – основная форма существования живой материи.
Ткани - совокупность клеток, выполняющих определенные функции. Различают эпителиальные, опорно-трофические ткани, мышечные, нервные.
Эпителиальные ткани (эпителий) выполняют разнообразные функции: защищают нижележащие ткани от неблагоприятных воздействий внешней среды, пропускают в организм питательные вещества в пищеварительном тракте, выводят продукты обмена веществ (через кожу и почки). Клетки некоторых видов эпителия выделяют определенные вещества - ферменты, гормоны. Клетки эпителия молочной железы способны синтезировать молоко.
Опорно-трофические ткани отличаются большим разнообразием. К трофическим, т. е. питающим, тканям можно отнести кровь и лимфу, к опорным - костную, хрящевую и соединительную ткани.
Мышечная ткань, сокращаясь, обеспечивает движение как отдельных органов, так и всего организма в целом. Различают гладкую, поперечнополосатую и сходную с ней сердечную ткани. Мышечный слой стенок внутренних органов (желудка, кишечника, мочевого пузыря, матки, кровеносных и лимфатических сосудов) состоит из гладкой ткани. Гладкая мышечная ткань способна растягиваться, не меняя напряжения. Благодаря этому при наполнении (до определенных пределов) желудка, мочевого пузыря, вымени давление в них не увеличивается. В отличие от поперечнополосатой гладкие ткани сокращаются гораздо медленней, но могут находиться в сокращенном состоянии длительное время, не обнаруживая признаков утомляемости.
Из поперечнополосатой ткани состоят скелетные мышцы.
Сердечная мышца и гладкие мышцы могут сокращаться под влиянием импульсов, зарождающихся внутри этих мышц.
Нервная ткань состоит главным образом из нейронов, представляющих крупную клетку с тонкими отростками разной длины. Нейроны в отличие от клеток других тканей способны возбуждаться и передавать возбуждение в виде нервного импульса другим нервным клеткам и мышечным тканям.
Органом называют часть тела животного, состоящую из различных тканей, объединённых в соответствующую форму и выполняющих специфическую функцию (печень, мозги, зуб и т.д.).
В многоклеточных организмах каждый из жизненно-важных процессов выполняется системой специализированных органов.
Например: обмен веществ осуществляется системами органов пищеварения, мочевыделения, крово- и лимфообращения. Размножение осуществляет система органов размножения, регулируемая нервной и эндокринной системами. Различные движения производятся системой органов движения.
Под системой органов понимают совокупность различных органов, связанных между собой функционально и часто морфологически, выполняющих один из основных жизненных процессов (дыхания, кровообращения и т.д.).
Всё системы в живом организме тесно взаимосвязаны, обеспечивают его жизнедеятельность и приспособление к окружающей среде.
Тема 2. Общая характеристика строения и функций
Системы органон движения
2. Мышечная система. Скелетные мышцы как активные органы движения. Строение мышцы как органа. Мышцы как главная составная часть мяса. Относительная… Вопрос 1. Костная система. Скелет как система органов движения и опоры. Типы…Плоскости тела
Сагиттальная– плоскость, проведённая вертикально вдоль тела животного. Направление той или иной детали описанного органа в сторону этой плоскости… Сегментальная - плоскость, проведённая вертикально поперёк тела животного.Рис. 2. Плоскости тела и направления расположения органов
Плоскости: а-а - срединная сагиттальная; б-б - сегментальные; в-в - фронтальные.
Направления: 1 - оральное (назальное), 2 - аборальное; 3 - краниальное; 4 - дорсальное; 5 - каудальное; 6 - вентральное; 7 - проксимальное; 8 - дистальное; 9 - медиальное; 10 - латеральное.
Отделы и области тела
Тело позвоночных животных делят на осевуючасть и конечности.
Осевая часть
У млекопитающих различают: голову, шею, туловище и хвост. Каждый из этих отделов делится на области (звенья или стати).
Голова (лат. kephale – голова). Костный остов её (череп) делится на мозговой (место, где расположен головной мозг) и лицевой отделы (ротовая и носовая полости).
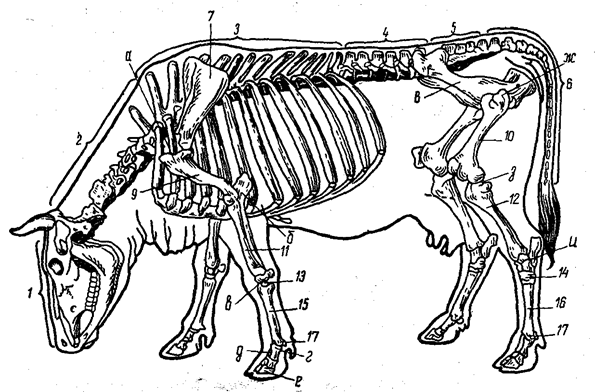
Рис. 3. Скелет коровы
1 - череп; 2 - шейный, 3 - грудной (с ребрами), 4 - поясничный, 5 - крестцовый отделы скелета туловища; б-хвостовые позвонки; 7 - плечевой пояс (лопатки); 8 - тазовый пояс; 9 - плечевая кость; 10 - бедренная кость; 11 - кости предплечья; 12 - кости голени; 13 - кости запястья; 14 - кости заплюсны; 15 - кости пясти; 16 - кости плюсны; 17 - кости пальцев. Суставы: а-плечевой сустав; б- локтевой; в - запястный; г - путовый; д- венечный; е - копытцевый; ж - тазобедренный; з- коленный; и- заплюсневый (скакательный) сустав.
В мозговом отделе различают области:
- затылочную;
- теменную;
- лобную;
- височную;
- околоушную.
На лицевом отделе выделяют:
- носовую;
- верхнечелюстную (верхней губы);
- нижнечелюстную (нижней губы);
- подглазную;
- щечную;
- область жевательной мышцы;
- подбородочную;
- подчелюстную.
В большинстве случаев их основой являются одноимённые кости и хрящевые образования.
Шея (лат. cervix; collum). Костным остовом шеи являются шейные позвонки (у млекопитающих их 7).
Туловище.Состоит из грудного, брюшного и тазового отделов.
Грудной отдел содержит спину и грудь и расположен между шеей и поясницей. Костной основой этого отдела являются:
- сверху – грудные позвонки (vertebra thoracatis);
- с боков – ребра (costa);
- снизу – грудная кость (os sternum).
Вместе они составляют грудную клетку (лат. thorax). В области грудной клетки на верхней передней части спины выделяют холку, образованную остистыми отростками грудных позвонков.
Брюшной отдел включает в себя поясницу (лат. lumbus) и живот (брюхо). Он имеет костный остов только в области поясницы, состоящий только их поясничных позвонков.
Тазовый отдел (лат. pelvis – таз) состоит из крестца и тазового пояса. Костной основой крестца служат крестцовые позвонки, сросшиеся в крестцовую кость. Тазовый пояс возник в процессе эволюции в связи с развитием тазовой конечности и служит стенками тазовой полости. Состоит из трёх парных костей: подвздошной, седалищной, лонной.
Хвост (лат. cauda). У разных животных имеет различную длину. Костной основой его являются хвостовые позвонки. Хвост делят на корень, тело и кончик.
Конечности
У наземных животных различают передние (или грудные) и задние (или тазовые) конечности. Они состоят из плечевого или тазового поясов и свободных конечностей.
Область плечевого пояса у низких (лягушка) позвоночных состоит из трех костей: лопатки, ключицы и коракоидной. У домашних млекопитающих, обладающих разнообразными движениями грудной конечности (например: кошки), встречается неполно развитая ключица.
У копытных животных, конечности которых приспособлены только к поступательному движению, развита только лопатка. Коракоидная кость у большинства млекопитающих редуцирована до небольшого отростка, сросшегося с нижним концом лопатки.
Грудная конечность
Плечо(лат. brachium) расположено ниже плечевого пояса и представлено остовом в виде плечевой кости.
Предплечье(лат. antebrachium) у всех животных расположено свободно. Его основа – две кости предплечья: лучевая (от лат. radius) и локтевая (от лат. ulna).
Кисть(лат.manus) делится на ряд звеньев: запястье, пясть и пальцы.
Запястье (лат. carpus) – это короткое утолщенное звено в середине свободной грудной конечности. В основе запястья лежат два ряда мелких костей запястья.
Пясть (лат. metacarpus) наиболее длинное звено нижней части конечности. Костный состав (количественно) у разных животных различен.
Пальцы (лат. digiti) называются порядковым числительным, при этом счёт ведется с внутренней стороны. У разных видов животных их бывает различное количество (от 1 до 5). Каждый палец состоит из трёх члеников или фаланг – промаксимальной (путовая), средней (венечная), дистальной (копытная), в основе которых лежат одноимённые косточки.
Тазовый пояс представлен безымянной костью (три сросшиеся тазовые кости: подвздонная, седалищная и лонная), крестцовой костью и первыми хвостовыми позвонками. Подвздошная и седалищная кости являются основой ягодичной области. Ягодичные области обеих сторон вместе с крестцом и началом хвоста образуют у копытных животных круп (от лат. croup).
Тазовая конечность
Бедро (лат. femor) расположено под тазом, его основу составляет бедренная кость. У копытных животных бедро, как и плечо, в своей значительной части не отделяется от туловища в действительно свободное звено и покрыто вместе с туловищем общим пластом кожи.
Голень (лат. crus) в основе имеет две кости:
- большеберцовую (лат. tibia);
- малоберцовую (лат. fibula).
Стопа(лат. pes, pedis) как и кисть делится на три звена: заплюсна, плюсна, палец.
Заплюсна (лат. tarsus) представляет собой короткое, несколько утолщенное звено, расположенное приблизительно на середине свободной конечности. Её остов – три ряда костей заплюсны, в том числе таранная (лат. talus), пяточная (лат. calcaneus) и центральная (лат. centrale).
Плюсна(лат. metatarsus) сходна с пястью, основу её составляют кости плюсны. У разных животных различное количество костей (от 1 до 5).
Пальцы (от лат. digiti) - костная основа – фаланговые косточки, как и на грудной конечности.
Развитие, форма и строение костей
Развитие. Костная ткань развивается из склеротомов сомитов мезодермы. Полное окостенение костей скелета наступает у крупного рогатого скота (КРС) к… Форма. В зависимости от функции и положения в скелете кости имеют различную форму. Различают пластичные и трубчатые…Соединение костей скелета
1 непрерывное - синартроз – сращение двух костей посредством различных тканей с образованием… - синдесмоз– кость + соединительная ткань (лучевая и локтевая кости копытных, кости черепа);Связки
Связки – это вспомогательные органы, которые укрепляют воздействие между костями.
Внутрисуставные связки выполняют механическую функцию, повышая надежность, укрепляя суставы и предотвращая их вывихи (крестовидная связка коленного сустава, круглая связка головки бедренной кости).
Внекапсулярные связки, укрепляющие сустав с боков и обеспечивающие движение сустава по строго определённой оси, ограничивая иные движения (большинство связок).
Внутрикапсулярные связки усиливают фиксирующую функцию основных связок (пальмарные связки запястья, плантарные связки заплюсны).
Вопрос 2. Мышечная система. Скелетные мышцы как активные органы движения. Строение мышцы как органа. Мышцы как главная составная часть мяса. Относительная масса мышц в теле животного и мясных тушах.
Общая характеристика мышц и их действия
Рис. 4. Поверхностные мускулы коровы: 1 - трапециевидный; 2 - дельтовидный; 3 - трехглавый мускул плеча; 4 - широчайший мускул спины; 5 - поверхностный…Мышца –это типичный паренхиматозный орган, в котором рабочую часть, или паренхиму, составляет поперечно-полосатая мышечная ткань, а остов (строму) соединительнотканные оболочки.
Пучки мышечных волокон (миофибрилл) объединены рыхлой соединительной тканью – эндомизием в пучки 1–го порядка. Пучки 1–го порядка объединяются внутренним перимизием в пучки 2–го, 3–го и т. д. порядков, составляющие основу мышечного брюшка.
Снаружи мышечное брюшко покрыто наружным перимизием (или эпимизием).
Наружный и внутренний перимизии участвуют в развитии внутримышечных соединительнотканных образованиях, по которым проходят сосуды и нервы и составляют основу сухожилий мышц.
Справка: Сухожилия(лат. tendo; tendines) – это мышцы, состоящие из плотной соединительной ткани, имеют светлую желтовато – розовую окраску. Сухожилия обладают огромной сопротивляемостью на растяжение, что делает их прочными и неутомимыми.
Сухожилия по форме бывают:
- лентовидные, часто встречаются в конечностях;
- пластинчатые (апоневрозы), располагаются преимущественно на туловище, обеспечивают крепость брюшных стенок, создавая большую площадь опоры для крепления мышц.
Внешнее строение мышцы
Скелетные мышцы в зависимости от выполняемой функции отличаются друг от друга соотношением мышечных пучков и соединительнотканных образований. Поперечники. Для суждения о различии мышц по их структуре, для сравнения силы… Анатомическим поперечником мышцы называют площадь его поперечного сечения.Вспомогательные приспособления мышц
1. фасции – покрывают мышцы, играя роль футляров, обеспечивают наилучшие условия для движения, облегчают крово- и лимфообращение. 2. бурсы– обнаруживаются в местах наибольшего трения мышц, связок, кожи и… 3. синовиальные влагалища– отличаются от бурс тем, что имеют гораздо большую длину и двухслойную стенку, почти…Тема 3. Система внутренних органов и органов
Крово- и лимфообращения
2. Общая характеристика систем органов пищеварения, дыхания, мочеотделения и размножения. 3. Строение сердца, закономерности хода и ветвления кровеносных сосудов. Общий… Вопрос 1. Закономерности строения, расположения и функции внутренностей. Понятие о полостях тела.ТИПЫ СТРОЕНИЯ ВНУТРЕННОСТЕЙ
Трубкообразные органы. Несмотря на резкие различия в строении, зависящие от функции, трубкообразные органы имеют вместе с тем черты общности. У них,… 1. Слизистая оболочка- tunica mucosa (1) - называется так потому, что она… Эпителиальный слой слизистой оболочки в местах, наиболее подверженных воздействию внешней среды (начальный участок…Рис. 5. Строение сердца
1 - нисходящая аорта (к органам тела), 2 - нижняя полая вена (от органов тела), 3 - правый желудочек, 4 - сухожильные нити, 5 - трехстворчатый клапан, 6 - полулунные клапаны при выходе легочной артерии, 7 - правое предсердие, 8 - легочные артерии (к легким), 9 - верхняя полая вена (от органов тела), 10 - аорта (к органам тела), 11 - легочная артерия (к легким), 12 - левое предсердие, 13 - легочные вены (от легких), 14 - полулунные клапаны аорты, 15 - двухстворчатый клапан, 16 - левый желудочек.
Сердце - своеобразный насос, непрерывно перекачивающий кровь по кровеносным сосудам. Продольной перегородкой оно разделено на правую и левую половины, каждая из которых состоит из двух полостей - предсердия и желудочка (рис. 5). Кровь по венам входит в предсердия, а выходит по артериям из желудочков, имеющих толстые мышечные стенки. Переход крови из предсердий в желудочки, а из желудочков в артерии регулируется соединительнотканными образованиями называемыми клапанами, которые автоматически закрываются и не дают крови течь в обратном направлении.
На работу сердца влияет целый ряд факторов. При повышенной физической нагрузке и психических воздействиях (например, испуге) чаще сокращаются стенки предсердий и желудочков. Частота сокращений сердца у отдельных видов животных различна. В состоянии покоя у лошади она составляет 32 - 42 раза в минуту, у крупного рогатого скота, овец, свиней - 60 - 80, у кур - до 300 раз. Частоту сердечных сокращений можно определить по пульсу - периодическому расширению кровеносных сосудов.
Круги кровообращения. Существуют два круга кровообращения - большой и малый. Венозная кровь от всех внутренних органов собирается в две полые вены - левую и правую, впадающие в правое предсердие. Из правого предсердия венозная кровь порциями переходит в правый желудочек, а из него по легочной артерии поступает в легкие, где отдает через легочную ткань углекислый газ и насыщается кислородом. Насыщенная кислородом кровь по легочным венам поступает в левое предсердие. Участок системы кровообращения, по которому кровь проделывает путь от правого желудочка через легкие в левое предсердие, называется малым, или дыхательным, кругом. Назначение малого круга кровообращения - удаление углекислого газа из крови и насыщение ее кислородом.
Из левого предсердия кровь попадает в левый желудочек, а оттуда в самый крупный артериальный сосуд- аорту. От аорты отходят артерии, разветвляющиеся на более мелкие. Органы и ткани снабжаются кровью через мельчайшие кровеносные сосуды - артериальные капилляры, имеющие очень тонкие стенки. Капилляры буквально пронизывают все ткани тела животного. Отдавая кислород и принимая углекислый газ и продукты обмена веществ в клетках, кровь превращается в венозную и оттекает от тканей и органов сначала по венозным капиллярам, а далее по венам.
Из левого желудочка кровь, двигаясь по артериальным, а затем по венозным сосудам и, наконец, попадая в правое предсердие, проходит большой круг кровообращения. Назначение большого круга кровообращения - снабжение кровью, обогащенной кислородом и • питательными веществами, всех органов и тканей тела.
Поскольку кровь движется по замкнутой системе, она оказывает давление на стенки кровеносных сосудов, причем давление падает с удалением крови от левого желудочка. Например, давление в аорте составляет 180-150 мм рт. ст., в артериальных капиллярах - 15-25, а в полых венах оно равно нулю. Поэтому повреждение артерий, особенно крупных, где кровь течет под большим давлением, связано с опасностью, так как животное может потерять много крови.
Кровь, кроветворение
Кровь состоит из плазмы и форменных элементов - эритроцитов, лейкоцитов и тромбоцитов. В крови около 20% сухого вещества и 80% воды. Плазма содержит… Эритроциты (красные кровяные тельца) очень важны для процесса дыхания. Они… Недостаток кислорода приводит к накоплению в организме недоокислившихся продуктов обмена - шлаков.Нервная система
Строение нервной системы. Нервную систему подразделяют на центральную и периферическую. К центральной нервной системе относят головной и спинной… Спинной мозг находится в спинномозговом канале, т е. внутри позвоночника.… Отростки нейронов - нервные волокна, выходящие из головного и спинного мозга, образуют периферическую нервную систему.…Внутренняя секреция. Терморегуляция
С током крови гормоны разносятся по всему телу. По выражению выдающегося русского физиолога А. А. Ухтомского, они сигнализируют «всем, всем, всем»,… К железам внутренней секреции относятся гипофиз 5 (рис. 6), щитовидная железа…Терморегуляция
При распаде белков, жиров и углеводов в теле животного образуется тепло, часть которого используется на нужды животного, а другая часть отдается в… Теплопроведение (кондукция) - переход тепла при соприкосновении тела животного… Конвекция - переход тепла с поверхности тела животного в окружающий воздух. Чем больше скорость движения воздуха, тем…Общая характеристика анализаторов
Научную основу работы органов чувств создали Иоганн Мюллер (нем., XIX в.) и его ученик Герман Гельмгольц. Их учение получило название «теория… Более 100 лет назад в работе «Рефлексы головного мозга» И.Н. Сеченов впервые… - воспринимающей – глаз;Механизм функционирования анализаторов
2. Генерация рецепторного потенциала; 3. Передача импульса на нервную клетку и генерация в ней потенциала действия… 4. Передача возбуждения по нервным проводниковым путям в кору больших полушарий;Общие свойства анализаторов
Рецепторные аппараты органов чувств обладают рядом общих свойств. 1. Высокая чувствительность к адекватным раздражителям (т.е. специфическим). … Например: для глаза в темноте порог чувствительности будет очень низким. Человек в полной темноте может увидеть…Строение и функции зрительного анализатора
Орган зрения в ходе эволюции животных изменялся и усложнялся от отдельных светочувствительных клеток до сложного зрительного анализатора млекопитающих.
Глаз – это орган, специально приспособленный для восприятия световых волн. У животных чувствительность фоторецепторов глаза лежит в пределах восприятия волн от ультрафиолетового до инфракрасного спектра (380 –750 нм или 0,38 –0,75 мкм).
Строение глаза
Глаз состоит из: - оптической части: роговица, передняя камера, зрачок, задняя камера,… - фоторецепторной части – сетчатки;Рис. 5. Схема строения глаза
Ход лучей в глазу
Преломление лучей при переходе из одной среды в другую называется рефракцией.Преломление лучей происходит потому, что все преломляющие среды имеют… - роговица – 8 мм - передняя сторона хрусталика – 10 ммФокус
Точка, в которой сходятся параллельные световые лучи, прошедшие через линзу, называется фокусом.
 |
Преломляющая сила линз измеряется в диоптриях (Д) и рассчитывается по формуле
Д= 1/f,где f фокусное расстояние (м)
Для человеческого глаза f = 1,7 см, следовательно, Д = 1/0,017 = 58,7 диоптрий.
Справка: 1 диоптрий – преломляющая сила линзы с фокусным расстоянием в 1 м.
В зависимости от того, где образуется фокус, выделяют три вида рефракции:
1. Нормальная рефракция (эмметропия) – характеризуется образованием фокуса точно на сетчатке. Это обеспечивает ясное изображение на ней видимых объектов.
2. Близорукая рефракция (миопия) – возникает обычно в результате врожденного удлинения глаза в передне-заднем направлении. В результате излишне сильного преломления лучей фокус будет находиться перед сетчаткой. Это влияет на четкость изображения удаленных предметов. Устраняют этот недостаток двояковогнутой рассеивающей линзой очков.
3. Дальнозоркая рефракция (гиперметропия) – наблюдается при увеличенном длиннике глаза. Недостаточная преломляющая сила способствует формированию фокуса за сетчаткой. Как и в случае с близорукостью на сетчатке получается круг светорассеивания. Устраняют очками с выпуклыми линзами.
Дальнозоркость может появляться в старческом возрасте. Но связана она уже с ухудшением упругости и эластичности хрусталика. При расслаблении цинновых связок он не может сильно изменять свою выпуклость, и старый человек отодвигает книгу при чтении. Называется это отклонение – пресбиопия.
К аномалиям рефракции следует отнести и астигматизм. Астигматизм – это несведение лучей в одной точке на сетчатке из за разности преломления лучей различными средами глаза. Это приводит к нечеткому изображению предмета.
Аккомодация и зрачковый рефлекс
Механизм аккомодации связан с сокращением ресничных мышц и изменением выпуклости хрусталика. Хрусталик находится в капсуле, которая за счет цинновых… С возрастом аккомодация ухудшается вследствие снижения эластичности… Зрачок – это отверстие в радужной оболочке. В толще радужки заложены гладкие мышечные волокна двух видов – циркулярные…Строение и функции сетчатки глаза
У человека в сетчатке глаза » 130 млн. палочек и » 7 млн. колбочек. Наружный слой сетчатки, прилегающий к сосудистой оболочке, состоит из… У ночных животных между пигментным слоем и фоторецепторами расположен слой, отражающий свет и состоящий из особых…Цветное зрение
- улучшает видимость предметов; - увеличивает полноту представления о них; - способствует лучшей и скорейшей ориентировке в окружающей среде;Бинокулярное зрение
Глаза – это парные органы, расположенные симметрично. Одновременное видение предмета двумя глазами называется бинокулярным зрением. Это обеспечивает более широкое поле зрения и видение предмета как бы с двух сторон. При бинокулярном зрении изображение предмета возникает на сетчатках обоих глаз в так называемых идентичных точках. Если изображение не будет попадать в идентичные точки сетчатки обоих глаз, то предмет будет двоиться.
Слуховой анализатор. Орган равновесия
В процессе эволюции у животных сформировался орган, воспринимающий и анализирующий звуковые колебания – слуховой анализатор. У млекопитающих слуховой аппарат делится на три части: - наружное ухо (ушная раковина, слуховой проход);Механизм восприятия звука
2. Барабанная перепонка начинает колебаться с частотой, соответствующей звуку. 3. Колебания барабанной перепонки передаются по системе косточек к внутреннему… 4. Стремечко передает колебания через мембрану овального окна преддверия на перилимфу.Диапазон
Звуки человеческой речи содержат частоты от 150 до 2500 Гц. Орган равновесия. Животные обладают способностью сохранять равновесие,… Перепончатые лабиринты каналов заполнены жидкостью и содержат отолиты (кристаллы СаСО3 и Са3 (РО4)2. В мешочках…Обонятельный анализатор
Периферическая часть обонятельного анализатора располагается в обонятельной зоне носовой полости. В этом эпителии выделяют опорные клетки,… Механизм обонятельной рецепции.При контакте молекул пахучего вещества со… Далее нервный импульс (величина которого зависит от концентрации пахучего вещества) передается по очень сложной цепи в…Вкусовой анализатор
С помощью грибовидных сосочков корм исследуется поверхностно, так как с этими сосочками он соприкасается в самом начале. Листовидные и валиковидные… В толще слизистой оболочки сосочков расположены рецепторные аппараты –… На языке выделяются зоны, которые реагируют преимущественно на определенные вещества:Температурная чувствительность
- холодочувствительные – расположены поверхностно; - теплочувствительные – лежат глубже, между дермой и подкожной клетчаткой. … Холодочувствительные – колбы Краузе, у человека их всего до 250 000. Могут возбуждаться и при действии тепла…Тактильная чувствительность
Тактильная чувствительность воспринимается следующими структурами: - тельце Фатер-Пачини – соприкосновение, давление (лежат глубоко в коже); - тельце Майснера – осязание (расположены в сосочках кожи);Болевая (ноцицептивная) чувствительность
Рецепторный аппарат болевого анализатора представлен свободными нервными окончаниями кожи и внутренних органов. Боль возникает при действии сильных… - 200 г на 1 мм2 кожи, - сильного света на глаз,Взаимодействие анализаторов
Функционально близко между собой органы обоняния и вкуса. Поэтому животные охотно поедают корм с ароматным запахом и наоборот. Ультрафиолетовые лучи, действующие на рецепторы кожи, снижают восприятие… Интенсивный шум или какая-либо мелодия приводит к торможению боли и изменению восприятия других раздражителей.Система органов движения.
Скелет
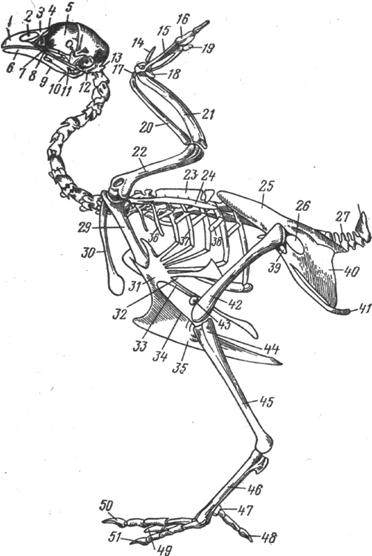
Рис. 15. Скелет курицы:
1 - резцовая кость; носовое отверстие; 3 - носовая и 4 - слезная кости; 5 - перпендикулярная пластинка решетчатой кости; 6 - зубная; 7 - небная; 8 - квадратно-скуловая; 9 - крыловидная; 10 - квадратная и 11 - суставная кости; 12 - барабанная полость; 13 - первый шейный позвонок; 14 - второй палец; 15 - третья пястная кость; 16 - третий палец; 17 - запястная лучевая кость; 18 - запястная локтевая кость; 19 - четвертый палец; 20 - лучевая; 21 - локтевая; 22 - плечевая кость; 23 - грудные позвонки; 24 - лопатки; 25 - подвздошная кость; 26 - седалищное отверстие; 27 - хвостовые позвонки; 28 - пигостиль; 29 - коракоидная кость; 30 - ключица; 31 - грудина; 32 - реберный; 33 - боковой и 34 - средний отростки; 35 - гребень грудины; 36 - астернальные и 37 - стернальные ребра; 38 - крючковидный отросток; 39 - запертое отверстие; 40 - седалищная, 41 - лонная и 42 - бедренная кости; 43 - коленная чашечка; 44 - малоберцовая; 45 - большеберцовая; 46 - плюсневая и 47 - первая плюсневая кости; 48 - первый; 49 - второй; 50 - третий и 51 - четвертый пальцы.
Мышцы
Мускулатура птиц отличается крайне неравномерным распределением на теле (рис. 16). Основная масса ее (45%) расположена в области грудной кости при переходе на крыло. Этой мускулатурой осуществляется работа крыла при полете. Вторым пунктом расположения мощной мускулатуры является верхняя половина тазовой конечности. Эти мышцы обеспечивают передвижение птицы по земле.

Рис. 16. Мышцы птицы
1 - полусухожильная; 2 - двуглавая бедра; 3 - напрягатель широкой фасции; 4 - портняжная; 5 - зубчатая вентральная; 6 - крыловой складки; 7- предлопаточная; 8 - широчайшая спины; 9 - мышцы крыла; 10 - мышцы, разгибающие шею и голову и отклоняющие их в сторону; 11 - мышцы, сгибающие шею и голову; 12 - косая брюшная наружная; 13 - поверхностная грудная; 14 - икроножная; /5 - пяточная и малоберцовая длинная; 16 - абдуктор четвертого пальца; 17 - сгибатель пальцев; 18- разгибатель пальцев.
Система органов кожного покрова и его производные
Перья.В пере различают стержень и опахало (рис. 17). Стержень состоит из очина и стебля. Очин - начальная, более толстая часть пера; стебель -… .Система органов пищеварения
Наибольшие особенности в строении органов пищеварения птицы наблюдаются в ротоглотке, передней и задней кишках.

Рис. 18. Ротоглотка кур
А - органы дна; Б - крыша ротоглотки; 1- верхушка языка; 2 - сосочки тела языка; 3 - основание 4 - кольцевидно-черпаловидные железы; 5 - пищевод; 6 - подчелюстные железы; 7 - вход гортань; 8 - гортанные сосочки; 9 - разрез подъязычной кости; 10- узкая часть небной щели; 11 - небные сосочки; 12 - хоаны; 13 - вход в слуховые грубы; 14 - отверстия челюстных желез; 16 - латеральные небные железы; 16 - медиальные небные железы; 17 - разрез мышц; 18 - глоточные железы; 19 - глоточные сосочки.

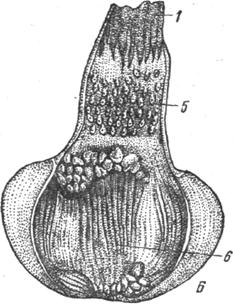
Рис. 19. Желудок курицы (А - снаружи, Б - вскрытый):
1 - пищевод; 2 - железистая и 3 - мышечная части желудка; 4 - двенадцатиперстная кишка; 5 - железы вскрытой части; 6-слизистая оболочка мышечной части желудка.
Средняя кишка, или тонкий отдел кишечника, состоит из двенадцатиперстной, тощей и подвздошной кишок. В стенке ее залегают пристенные - общекишечные - железы. Застенными железами являются печень и поджелудочная железа (рис. 20). Желез двенадцатиперстной кишки у птиц нет. Длина кишечника зависит от вида корма. Так, кишечник уток и гусей длиннее тела в 4 -5 раз, а кишечник кур - в 5-6 раз. В кишечнике птиц длинные ворсинки, а слизистая оболочка его собрана в складки, увеличивающие путь пищи по кишечнику.
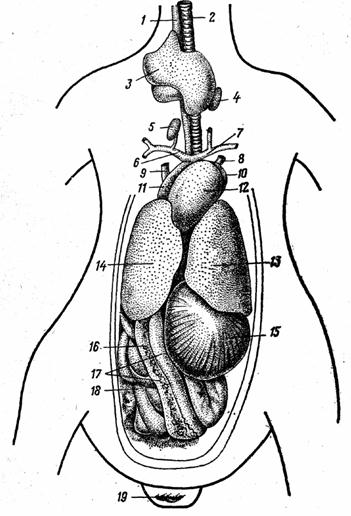
Рис. 20. Вскрытая полость тела курицы с брюшной стороны:
1-пищевод, 2 - трахея; 3 -зоб, 4 и 5 - щитовидные железы; 6 и 7- правая и левая плечеголовные артерии; 8 в 9 - левая и правая краниальные полые вены; 10 и 11 - левое и правое предсердия; 11 - желудочек сердца; 13 и 14 - левая и правая доля печени; 15 - мышечная часть желудка; 16 - поджелудочная железа; 17 - двенадцатиперстная кишка; 18 - тощая кишка; - отверстие клоаки.
Двенадцатиперстная кишка выходит из переднего участка мышечной части желудка и идет до таза, а затем возвращается, образуя петлю из двух колен. В этой петле расположена поджелудочная железа.
Тощая и подвздошная кишки подвешены на тонкой брыжейке, соприкасаясь друг с другом и образуя спиральные завитки. Подвздошная кишка - открывается в заднюю кишку на границе слепых кишок с прямой кишкой.
Печень -hépar - птиц довольно большая, делится на две доли и занимает значительную часть вентральной половины грудобрюшной полости. На правой доле печени расположен желчный пузырь, который отсутствует у цесарок, голубя, страуса.
Поджелудочная железа - páncreas - расположена в петле двенадцатиперстной кишки, у уток и гусей имеет две доли и два протока, у кур - три доли и три протока.
Задняя кишка, или толстый отдел кишечника, птиц ободочной кишки не имеет. У уток, гусей и кур две слепые кишки. Вершинами они обращены краниально, а от подвздошной кишки отграничиваются круговой складкой. Прямая кишка переходит в клоаку, которая двумя поперечными складками делится на три отдела: передний, средний и задний. В передний отдел открывается прямая кишка, в средний - мочеточники, семяпроводы (у самцов) и яйцеводы (у самок). Задний отдел клоаки заканчивается анальным отверстием, через которое непереваренные остатки корма, смешанные с мочой, а у самок также яйца выбрасываются наружу. Через этот же отдел проходит и сперма самцов во время спаривания. На дорсальной стенке его у молодых птиц имеется выпячивание - фиброзная (фабрициева) сумка, которая у взрослых птиц редуцируется.
Система органов дыхания и мочевыделения
Легкие сравнительно малы и дополняются девятью воздухоносными мешками. Лежат легкие в углублениях между ребрами. Последние оставляют на лёгких… Воздухоносные мешки представляют собой выпячивание слизистой оболочки бронхов.… Воздухоносные мешки, важнейшие приспособления для полета птиц, обеспечивают им интенсивный газообмен, вентиляцию и…Рис. 21. Схема легких и воздухоносных мешков птицы
1 - межключичный мешок; 2 - шейные мешки; 3 - подмышечный дивертикул; 4 - ход в плечевую кость; 5 - краниальные и 6 - каудальные грудные мешки; 7 - брюшные мешки; 8 - главный бронх; 9 - преддверие главного бронха с отверстиями во вторичные бронха; 10 - легкие; 11 - эктобронх брюшного мешка; 12 - мешковый бронх каудальных грудных мешков; 13 - мешковый бронх брюшных мешков
Система органов мочевыделения птиц состоит из почек и мочеточников (рис. 22.). Мочевого пузыря у птиц нет. Почки довольно длинные, лежат в углублениях пояснично-крестцового отдела позвоночного столба и подвздошной кости и делятся на три доли: переднюю, среднюю и заднюю. Мочеточники располагаются на медиальном крае почек и открываются в клоаку.
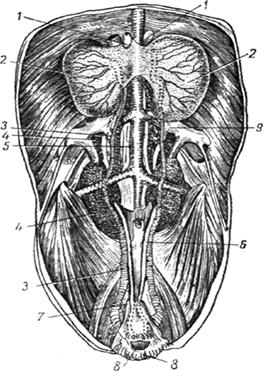
Рис. 22. Органы мочевыделения и размножения петуха:
1 - надпочечник; 2 - семенник; 3 - семяпровод; 4 - почки; 5 - аорта; 6 - мочеточник; 7 - отрезок прямой кишки; 8 - клоака; 9 - общая подвоздушная вена.
Система органов размножения
Рис. 23. Органы размножения курицы: А - 1 - легкие; 2-почки; 3 - повздошная кишка; 4 - слепые кишки (обрезаны); 5 - пряма кишка; 6 - воронка яйцевода; 7 -…Система органов кровообращения.
Рис. 24. Сердце птицы: 1 - легочная артерия; 2 - левое предсердие; 3 - легочные вены; 4 - левая краниальная полая вена; 5 - левый желудочек;…Нервная система и органы чувств
Рис. 25. Головной мозг птицы (А - с латеральной стороны, Б - с вентральной): … I - обонятельный нерв и луковица; II - зрительный нерв; III - глазодвигательный; IV - блоковый; V -тройничный; VI -…МОДУЛЬ II
Тема 6. Клетки и ткани как основные структурные элементы
Организма животного
1. Общие понятия о гистологическом строении технологического сырья мясной промышленности.
2. Основные разделы гистологии.
3. Структурная организация клетки и межклеточного вещества.
4. Краткая гистологическая характеристика тканей животного.
Вопрос 1. Общие понятия о гистологическом строении технологического сырья мясной промышленности.
Любой орган состоит из клеток, которые, в свою очередь, образуют соклетия, симпласты, ткани. Микроскопирование этих структур позволяет понять их… Особенно важное значение имеет знание гистологии мышечных тканей, так как…Вопрос 2. Основные разделы гистологии.
Гистология (histos - ткань, logos - учение) - учение о тканях. В действительности же курс гистологии отнюдь не ограничивается изучением тканей, он состоит из следующих разделов: 1) цитологии (учение о клетках); 2) общей эмбриологии (учение о ранних стадиях развития зародыша); 3) гистологии в собственном смысле этого слова и 4) микроскопической анатомии с основами частной эмбриологии. Первые три раздела часто объединяют под названием «общая гистология», а курс микроскопической анатомии и частной эмбриологии называют «частная гистология».
Гистология - отрасль биологии, которая познает сложные и очень тонкие процессы возникновения нового организма из оплодотворенного яйца. Она прослеживает все этапы развития клеток, исследует строение их, а равно и тканей, органов, изучает условия, необходимые для нормального их существования. Её можно определить как науку, трактующую о возникновении и развитии тончайшей структурной организации клеток, тканей и органов человека и животных, о ее функционировании и о возможностях направленного воздействия на нее в интересах практики.
Следует, однако, иметь в виду, что гистология занимается не только тончайшим строением организма; в настоящее время такое изучение является не самоцелью, а лишь средством познания общих биологических закономерностей, определяющих жизнь организма на всех стадиях онтогенеза. Знание этих закономерностей абсолютно необходимо для разработки эффективных способов вмешательства в жизнь организма, что важно и для лечения животных и для повышения их продуктивности.
Вопрос 3. Структурная организация клетки и межклеточного вещества.
Каждая клетка организма находится во взаимодействии с окружающей средой и в результате постоянного обмена веществ с ней изменяется в зависимости от… - химическим составом; - характером обмена веществ;Органоиды общего назначения
Система мембран сети непрерывна: она сливается с ядерной оболочкой клетки. Это очень гибкая, постоянно перестраивающаяся структура. Различают два вида эндоплазматической сети: гладкую (агранулярную) и… На мембранах шероховатой (гранулярной) ЭПС расположены рибосомы (состоящие из белка и рибонуклеиновой кислоты).Пластинчатый аппарат (или комплекс Гольджи).
Функции комплекса Гольджи: - накопление и формирование секреторных гранул; - синтез полисахаридов и липидов;Органоиды специального назначения
- миофибриллы – в мышечных клетках, обеспечивают мышечное сокращение; - нейрофибриллы – в нервных клетках, обеспечивают проведение нервного… - тонофибриллы – в эпителиальных клетках, обеспечивает прочность клеток и их объединение в пласт и др.Включения
Включения имеют вид глыбок, капель, вакуолей, гранул, кристаллов и хорошо видимы даже в световой микроскоп. По составу включения бывают: - белковые – видимые в микроскоп в норме встречаются только в яйцеклетках в… - жировые – в виде жиров и липидов присущи каждой животной клетке. Иногда заполняют почти всю клетку.Ядро
Ядро (nucleus) – жизненно необходимая составная часть клетки. Часто оно располагается в центре. Как правило, в клетке бывает одно ядро, однако некоторые клетки (остеокласты, эпителиоциты, эндотелий) имеют несколько ядер (многоядерные). Встречаются и безъядерные клетки – эритроциты (живут не долго - до 120 суток).
Форма и размеры ядер разнообразны, но типичны для каждого вида клеток. Форма ядер в значительной мере обусловлена формой клетки: у шаровидных – ядро круглое, у цилиндрических – овальное, у вытянутых - веретенообразных мышечных – вытянутое и т. д.
У клеток, меняющих свою форму (лейкоциты), ядра могут быть даже дольчатые.
Размеры ядра зависят от размеров клетки и от её функционального состояния (и возраста). Для каждого типа клетки отношение размеров ядра к размеру цитоплазмы постоянно.
В ядре различают ядерную оболочку (кариолемму), хроматин, ядрышко и ядерный сок.
Ядерная оболочка – состоит из двух элементарных липопротеиновых мембран, разделённых пренуклеарным пространством в 20-100 нм. Наружная мембрана может быть связана с мембранами цитоплазматической сети. Наличие рибосом на поверхности этой мембраны обуславливает синтез белка. Внутренняя мембрана не имеет рибосом. К ней прикреплены участки хроматина, чем обеспечивается упорядоченное расположение хроматина в определённой части ядра. Через правильные интервалы наружный и внутренний листки мембраны смыкаются друг с другом, образуя ядерные поры, через которые ядро и цитоплазма обмениваются своим содержимым. Размер пор от 20 до 80-90 нм. Количество 10-30 до 100 пор на 1 мкм3.
Ядерный сок (кариоплазма, кариолимфа) – микроскопически бесструктурное вещество ядра. Он содержит воду, простые белки ДНК, ферменты и соединения, участвующие в процессах синтеза нуклеиновых кислот, белков и др. веществ, йоны К+, Mg+, и др.
Ядрышко (nucleolus) - самое плотное образование клетки, тельце сферической формы диаметром 1-5 мкм. Ядрышко - это участок хромосомы (ядрышковый организатор), на котором образуется РНК, поступающая в цитоплазму. Ядрышко не имеет оболочки, и вещество ядрышка свободно соприкасается с кариоплазмой.
Содержимое ядрышка – РНК, ДНК и белок, йоны металлов (в частности цинк).
Функция ядрышка – синтез и накопление рибонуклеопротеидов и формирование рибосом.
Хроматин (от лат. chroma – краска)- плотное вещество, хорошо воспринимающее красители, особенно основные. Хроматин – это по существу хромосомы в определенном состоянии спирализации. По химическому составу хроматин состоит из: ДНК (до 40%), РНК (до 1%), белков (60%).
ДНК способна к самоудвоению. При разъединении цепей нуклеотидов (путем разрыва водородной связи ферментом ДНК – полимеразой) каждая половина ДНК из элементов окружающей кариоплазмы создает недостающую половину. При делении клетки её ДНК таким образом удваивается, и дочерние клетки получают полную структуру ДНК, соответствующую материнской.
Синтез белка: ДНК каждой клетки имеет свою специфическую структуру, которая определяется порядком чередования трех нуклеотидов (триплетов). Этот процесс называется – транскрипция.
Порядок чередования нуклеотидов в молекуле и РНК в свою очередь определяет порядок чередования аминокислот в молекуле белка. На каждые три нуклеотида «устанавливается» одна аминокислота. Это процесс называется – трансляция.
Таким образом, ДНК ядра выполняет функцию передачи наследственных признаков. Кроме того, ядро отвечает за специфичность белков организма и является регулятором обменных процессов.
Межклеточные связи. В состав связующего комплекса входят троякого рода структуры: замыкающий поясок, поясок слипания и многочисленные десмосомы (некоторые авторы относят к комплексу только первые две структуры). Они увеличивают механическую прочность соединения клеток и участвуют в регуляции перехода веществ по межклеточным щелям.
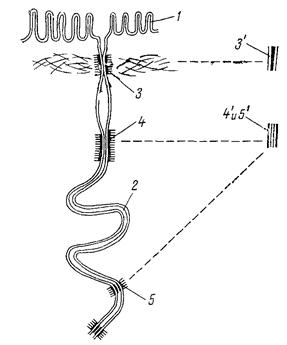
Рис 36. Разные типы соединения смежных клеток
1 - микроворсинки, 2 - межклеточное пространство, 3 - замыкающий поясок, 3' - пятислойность пояска, возникшая благодаря слиянию плазмалемм, 4 - поясок слипания, 5 - десмосома, 42 и 5' - между смежными плазмалеммами видно межклеточное пространство, содержащее кислый мукополисахарид.
Межклеточные щели заполнены своеобразным склеивающим веществом, или «межклеточным цементом». Это вещество пластично, не препятствует изменениям объема и формы клеток и даже их перемещению. Например, если вывернуть «наизнанку» тело гидры, так что внутренние клетки, выстилающие пищеварительную полость, окажутся снаружи, то через некоторое время все клетки снова займут их естественное положение. В простейшем случае клетки удерживаются вместе благодаря слипанию их поверхностей, для чего необходимо наличие солей кальция. Зародыш морского ежа, перенесенный в морскую воду, лишенную кальция, рассыпается на составляющие его клетки. У высших животных склеивающее вещество имеет сложный полисахаридный состав и обладает высокой видовой специфичностью, но и у них инъекция кальция в сосуды увеличивает прочность межклеточных соединений в кровеносных капиллярах.
Межклеточное вещество. В некоторых тканях клетки образуют большие количества межклеточного вещества. В нем нет органелл, обмен веществ протекает медленно, тем не менее, оно живое, выполняет опорную функцию и участвует в минеральном обмене. Эта своеобразная форма неклеточного живого вещества теснейшим образом связана с клетками и является их производным. Такого рода структуры называют метаплазматичвскими. В некоторых тканях смежные клетки плотно соединяются своими отростками. При рассматривании в световой микроскоп границ между ними не видно. Такие сетевидные клеточные скопления называют синцитием (то есть соклетием). В действительности слияния клеток в синцитии не происходит, они отграничены одна от другой клеточными мембранами (плазмалеммами). Встречаются, наконец, случаи (мышечные ткани), когда более или менее объемистые массы протоплазмы не расчленяются на клетки, содержат много ядер и обильную цитоплазму с присущими ей органеллами. Эти структуры называют симпластом (syn - совместно, plastos - образованный).
Таким образом, наряду с клетками, являющимися основной формой живого вещества, в теле животных и человека существует и межклеточное (метаплазматическое) вещество. Клетки обычно отделены друг от друга межклеточными щелями. При отсутствии щелей, видимых в световой микроскоп, возникают синцитии сетевидного строения. Встречаются и не расчлененные на клетки многоядерные массы цитоплазмы - симпласты. При повреждении симпласт может распасться на отдельные клетки, и, наоборот, отдельные клетки могут слиться в симпласт.
Деление клеток (амитоз, митоз, мейоз)
Приведенное выше описание ядра относится только к неделящейся клетке. Чтобы полнее оценить роль ядра, необходимо внимательно изучить процесс деления клеток.
Самовоспроизведение, или репродукция - общее свойство всех живых систем. С репродукцией связаны рост организма, замещение стареющих и отмирающих частей, регенерация поврежденных тканей, обновление постепенно стареющей структурной организации клетки, теряющей способность к биосинтезу.
Преобладание ассимиляции сопровождается ростом клетки, а рост приводит к затруднению, и даже к прекращению ассимиляции. Объем цитоплазмы клетки требует больше питательных веществ, а площадь поверхности мембраны не способна их пропустить в нужном количестве. Это противоречие устраняется тем, что разросшаяся до критических размеров клетка делится на две дочерние.
Различают три типа деления соматических клеток:
- митоз (или кариокинез) (от лат. mitos – нить; caryon – ядро; kinesis – движение, изменение) – непрямое деление;.
- амитоз (от лат. а – без; mitos – нить) – прямое деление;
- мейоз (от лат. meiosis – уменьшение) – редукционное деление.
Вопрос 4. Краткая гистологическая характеристика тканей животного.
Оплодотворенная яйцеклетка в процессе своего деления (дробления) и развития превращается в сложный многоклеточный организм. В ходе развития… Процесс образования специализированных тканей из клеточного материала…Ткань - это исторически сложившаяся система клеток и неклеточных структур, характеризующаяся общим строением, функцией и происхождением.
Тканям присуща физиологическая и репаративная регенерация. Физиологическая регенерация – это естественный процесс восстановления и… Репаративная регенерация – процесс восстановления целостности тканей и органов после повреждения.Эпителиальная ткань
Эпителиальная ткань (или эпителий) развивается из всех трех зародышевых листов. Эпителий располагается у позвоночных животных и человека на… В эпителиальном пласте кровеносных сосудов, как правило, нет. Питание… Основными отличительными признаками эпителиальных тканей являются:Железистый эпителий
Экзокринные железы имеют выводные протоки и выделяют секрет в полость внутренних органов или на поверхность. Примеры: слюнные, потовые, молочные, железы желудка, печень, сальные и т.д. … Эндокринные не имеют выводных протоков, их секрет поступает непосредственно в кровь или лимфу.Опорно-трофические ткани
Опорно-трофические ткани образуют каркас (строму) органов, осуществляют трофику органа, несут защитную и опорную функции. К опорно-трофическим тканям относят: кровь, лимфу и соединительные ткани.… Всем опорно-трофическим тканям присущи следующие признаки:Кровь
Кровь состоит из клеток (форменных элементов) и межклеточного вещества – плазмы. Плазма – это жидкость соломенного цвета, содержащая различные белки, жиры, углеводы, конечные продукты обмена, минеральные соли. Во взрослом организме клеточные элементы крови образуются в красном костном мозге, селезёнке и лимфатических узлах. Форменные элементы крови представлены эритроцитами, лейкоцитами, кровяными пластинками (у птиц - тромбоцитами).
Лимфа
В организме плазма постоянно просачивается сквозь стенку кровеносных капилляров и включается в состав тканевой жидкости и межклеточного вещества. Тканевая жидкость проникает в лимфатические капилляр, дальше идет по лимфатическим путям, проходит лимфатические узлы. В лимфоузлах она обогащается лимфоцитами (95%) и моноцитами, другие форменные элементы быстро разрушаются. Затем лимфатические сосуды впадают в грудной проток, и оттуда лимфа поступает в венозную кровь. Если нарушается отток жидкости через лимфатическую систему, возникают отеки тканей.
По химическому составу лимфа близка к плазме крови, но содержит меньше белковых тел.
Собственно соединительные ткани.
1. Рыхлая волокнистая – распространена в организме повсеместно, составляя основу всех слизистых оболочек трубкообразных органов. 2. Плотная волокнистая – она подразделяется на: а) плотную неоформленную соединительную ткань, которая входит в состав основы кожи (сетчатый слой дермы);Хрящевая ткань
Хрящ сверху покрыт надхрящницей – плотной соединительной тканью, состоящей из коллагеновых волокон и удлинённых клеток. Надхрящница постепенно… Гиалиновый хрящ наиболее распространен в организме. Из него состоят: большая… Эластический хрящ образует ушные раковины, надгортанные и частично черпаловидные хрящи гортани.Костная ткань
Клеточные формы костной ткани представлены остеобластами, остеоцитами и остеокластами. Остеоциты располагаются по окружности на периферии костной… Межклеточное вещество может быть аморфным (осеомукоидом) и волокнистым… По характеру расположения структурных элементов различают грубоволокнистую и пластинчатую костные ткани.…Мышечные ткани
Все виды мышечных тканей объединены в одну группу на основании общности их функции – сокращения. Особенности строения разных мышечных тканей… Гладкая мышечная ткань развивается из мезенхимы, скелетная – из миотомов…Нервная ткань
униполярные – с одним неразветвленным отростком; ложноуниполярные – с одним разветвлённым отростком; биполярные – с двумя отростками;Тема 7. Мышечная ткань.
1. Общая характеристика гладкой и поперечно-полосатой мышечной ткани.
2. Микроскопическое и электронно-микроскопическое строение мышечной ткани как основы мяса.
3. Изменения структуры мышечной ткани в процессе автолиза и на разных стадиях технологической обработки.
Вопрос 1. Общая характеристика гладкой и поперечно-полосатой мышечной ткани.
По происхождению, физиологическим свойствам и строению мышечные ткани делят на гладкую, поперечнополосатую и близкую к ней по строению сердечную.…ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ
Сформировавшаяся гладкая мышечная ткань состоит из удлиненных клеток, плотно прилегающих друг к другу (рис. 61). Благодаря тонким прослойкам… При изучении в световом микроскопе обнаруживаются оформленные миофибриллы,…Рис. 61. Рыхлая сеть гладкомышечных клеток эндокарда.
Иннервируется гладкая мышечная ткань вегетативной (автономной) нервной системой, и действие ее непосредственно не зависит от коры головного мозга, хотя и контролируется ею.
ПОПЕРЕЧНОПОЛОСАТАЯ МЫШЕЧНАЯ ТКАНЬ
Развивается поперечнополосатая мышечная ткань из миотомов, являющихся частью сомитов мезодермы. Миотомы содержат клетки удлиненной формы -… Рис. 62. Поперечнополосатая мышечная ткань:Рис 65. Схема строения мышцы
1 - наружный перимизий 2 - внутренний перимизий, 3 - кровеносные сосуды, 4 - нерв 5 - жировые клетки, 6 - эндомизий 7 - мышечные волокна в поперечном разрезе (точками обозначены миофибриллы в мышечных волокнах)
Группы поперечнополосатых мышечных волокон при помощи соединительной ткани (эндомизий) связываются в пучки I порядка (первичный мышечный пучок). Несколько пучков I порядка более значительным количеством соединительной ткани (внутренний перимизий) объединяются в пучки II порядка (вторичный мышечный пучок). Пучки II порядка, соединяясь между собой при помощи новых слоев соединительной ткани, образуют пучки III порядка (третичный мышечный пучок) и т. д. Наконец, самые наружные слои соединительной ткани окутывают всю мышцу (наружный перимизий). Все прослойки соединительной ткани, проходящие между пучками различных порядков, а также между отдельными волокнами в пучке, связаны и составляют единый соединительнотканный каркас - строну мышцы (рис. 65) По прослойкам соединительной ткани проходит большое количество кровеносных и лимфатических сосудов, а также нервов. Иннервируется поперечнополосатая мышечная ткань черепномозговыми и спинномозговыми нервами. В составе последних находятся как двигательные волокна, передающие возбуждение от головного мозга в мышцу, так и чувствительные, передающие возбуждение от мышцы в головной мозг. Контролируется работа мышц непосредственно корой головного мозга.
СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ
Эта ткань образует один из слоев стенки сердца - миокард. Она делится на собственно сердечную мышечную ткань и проводящую систему. Рис. 66. Схема строения сердечной мышечной ткани:Вопрос 2. Микроскопическое и электронно-микроскопическое строение мышечной ткани как основы мяса.
Основной структурной единицей гладкой мышечной ткани является клетка – гладкий миоцит, поперечнополосатой скелетной – волокно, а сердечной – клетки (сердечные миоциты), объединенные в волокно.
Гладкая мышечная ткань находится в стенке кишок, сосудов, молочных альвеол. Поперечнополосатая ткань – это мышцы тела (соматическая мускулатура). Сердечная ткань (рабочая и проводящая) образует сердце.
Строение саркомера
Каждый саркомер отделен друг от друга телофрагмой или Z-мембраной.
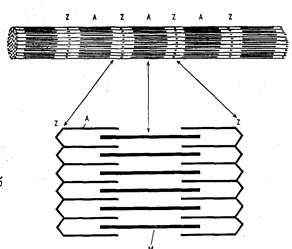
Рис. 2. Строение миофибрилл поперечнополосатой мышечной ткани.
а — строение одиночной миофибриллы;
б — схема строения саркомера:
А — актиновые (тонкие) протофибриллы;
Μ — миозиновые (толстые) протофибриллы;
Z — диск, содержащий т-актинин
При сокращении мышечного волокна концы актиновых и миозиновых нитей заходят один за другой, образуя зону перекрытия, кажущуюся наиболее темной.
В зоне перекрытия образуется актомиозиновый комплекс. Здесь распадаются молекулы аденозинтрифосфорной кислоты (АТФ), и освобождается энергия, необходимая для сокращения.
Вопрос 3. Изменения структуры мышечной ткани в процессе автолиза и на разных стадиях технологической обработки.
Мясо в основном состоит из мышечной ткани. Гладкомышечная ткань не имеет большого практического значения, а поперечнополосатая мышечная ткань - это основа любого вида мяса. Мышечная ткань в основном состоит из белковых элементов, поэтому является ценнейшим питательным продуктом для человека.
При технологической обработке мышечная ткань подвергается множественным изменениям, которые определяют ее качественные и вкусовые особенности.
ПЕРЕЖИВАНИЕ ОТДЕЛЬНЫХ ЭЛЕМЕНТОВ МЯСА
Сроки такого переживания для разных органов различны. Они в значительной степени зависят от условий, в которых находится орган. Отдельные мышечные волокна могут сохранять признаки жизнеспособности до трех… Автолиз мяса - это чрезвычайно сложный и многогранный процесс до настоящего времени еще недостаточно ясен. Однако…МИКРОСТРУКТУРА ГОВЯЖЬЕГО ПАРНОГО МЯСА
Микроскопический анализ четко выявляет общие структурные особенности исследуемых мускулов парного мяса, а также различия в их внутренней структуре.… Обращает на себя внимание гофрированная, или складчатая, форма многих мышечных… На поперечных срезах видна также и разница в строении. В длиннейшем мускуле спины более крупные мышечные волокна. Они…Рис. 98. Расположение эритроцитов в большом поясничном мускуле между волокнами.

Рис. 99. Рваные поперечные трещины в мышечных волокнах.
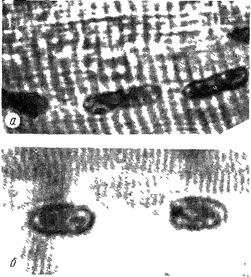
Рис. 100. Структура ядер в расслабленном волокне и в узлах сокращения:
а - в расслабленном; б - в сокращенном.
К характеристике структуры узлов сокращения относятся фигуры «веера» (рис. 91) чаще и четче выявляющиеся в мышечных волокнах большого поясничного мускула.
В свою очередь эти фигуры свидетельствуют о наличии каких-то механизмов, расположенных по длине мышечных волокон, которые способствуют образованию узлов сокращения не на всю глубину волокна.
Специфические особенности узлов - это резкое сокращение миофибрилл в пределах узла и поперечные разрывы мышечных волокон по узлам или в непосредственной близости от узлов (рис. 87, 88, 90). При этом следует отметить, что в начальные сроки после убоя (от 30 мин до 1-1,5 ч) разрывов по узлам в мышечной ткани встречается мало, а в более поздние сроки (через 4- 6 ч) их становится больше, особенно много в период rigor mortis.
Эти данные свидетельствуют о том, что разрывы волокон по узлам сокращения - это закономерное явление, обусловленное не механическими, а скорее внутримышечными молекулярными процессами в узле сокращения. При этом число разрывов в пределах одного узла может достигать 2-3 и более (рис 84, 88, 90).
Из структурных элементов в парном мясе особое внимание привлекают ядра мышечных волокон. В расслабленных волокнах они имеют овальную, вытянутую, иногда палочковидную форму (рис. 99). Такие ядра обычно хорошо структурированы, в них видна хроматиновая гранулированность. Совершенно иной вид ядра принимают в пределах узлов сокращения (рис. 100). По общему виду они более компактны. Следовательно, размер ядер в узлах сокращения несколько уменьшается (табл. 3).
Значительно труднее говорить о количественном сравнении соединительной ткани в мускулах, так как идентичных мест сравнения для соединительной ткани в разных мускулах трудно или даже невозможно найти. Прослоек больше в длиннейшем мускуле спины, в связи с этим и островки жировой ткани в нем встречаются чаще. В прослойках соединительной ткани большого поясничного мускула чаще обнаруживаются скопления лимфатических клеток, которые иногда выявляются в виде обширных островков, занимающих всю ширину прослойки на большом расстоянии.
Итак, в парном мясе в обоих мускулах обнаруживаются разная степень сокращенности в мышечных волокнах и разное количество сокращенных и гофрированных волокон.
В сравниваемых мускулах постоянно формируются узлы сокращения. Их меньше в первые часы после убоя, а в более поздние часы проходящего автолиза мяса число их заметно увеличивается. Структурно узлы сокращения различаются в разных мускулах.
Таким образом, по чисто морфологическим особенностям волокон, по их саркомерам, узлам сокращения и другим признакам можно судить о функциональных и качественных различиях отдельных мышц из разных топографических участков тела животного.
МИКРОСТРУКТУРА ГОВЯЖЬЕГО ОХЛАЖДЕННОГО МЯСА
В 1970 г. А. А. Соколов, Д. В. Павлов, А. С. Большаков, Н. К. Журавская и др. в своих работах установили, что охлажденным мясом считается такое,… Автолитические изменения к концу охлаждения делают мясо пригодным для… В условиях бескислородного снабжения еще существуют некоторые живые элементы тела и осуществляются процессы…МИКРОСТРУКТУРА ОХЛАЖДЕННОГО ГОВЯЖЬЕГО МЯСА ПРИ ХРАНЕНИИ
В 1970 г Н. П. Янушкин и И. А. Лагоша установили, что при хранении охлажденного мяса большое значение имеет образование корочки подсыхания в… Гистологический анализ показал, что уже через 1 сут хранения мышечные волокна… Через сутки охлаждения диаметр одного- двух поверхностно расположенных волокон уменьшается.ГИСТОЛОГИЧЕСКАЯ СТРУКТУРА
И ИЗМЕНЕНИЯ ЗАМОРОЖЕННОГО МЯСА ПРИ ХРАНЕНИИ
Замораживание мяса является сложным процессом. Ход его в значительной степени зависит от продолжительности периода, прошедшего после убоя животных,… Замораживание мяса необходимо для предотвращения размножения в нем… Гистологический анализ замороженного парного и охлажденного мяса показал, что уже после хранения в течение I сут при…Рис. 102. Зоны, возникающие в процессе замораживания парного мяса
(по Тинякову, Писменской, Костенко);
а - фация и первая зона (подсыхания); б - вторая зона (быстрой кристаллизации);
в - третья зона (перехода к нормальным волокнам)
В мясе, замороженном в парном состоянии и хранившемся 3-9 мес., поверхностная зона, лежащая непосредственно под фасцией, самая узкая (рис. 102). В зависимости от топографии ее толщина 250-350 мкм. Поверхностная зона состоит из стройных, компактно уложенных мышечных волокон толщиной 15,6 мкм (в 3-4 раза тоньше обычных волокон парного мяса). Все волокна этой зоны сохраняют плотное расположение и имеют поперечную исчерченность, хотя она четко не везде обнаруживается.
Во второй (средней) по глубине зоне, резко отличающейся от первой, многие мышечные волокна сильно фрагментированы и в значительной степени деформированы (см. рис. 102). Располагаются они рыхло, и в промежутках между ними часто обнаруживаются просветы с неровными краями. Эти просветы, по-видимому, точно повторяют формы водяных кристаллов, которые находились здесь до размораживания мяса. Ширина второй зоны при замораживании парного мяса 1-2 мм. Вторую зону следует считать зоной сравнительно интенсивной кристаллизации водной фазы. Энергичные процессы кристаллизации в этой зоне являются причиной рыхлого и хаотического расположения мышечных волокон, их фрагментации и деформации. Толщина волокон во второй зоне 27,4 мкм, т. е. почти в два раза больше, чем в первой.
Толщина мышечных волокон в третьей зоне (см. рис. 102) 33,6 мкм. В ней мышечные волокна в большей степени приближаются к нормальному виду, хотя в 1,5-2 раза тоньше обычных. В их расположении и структуре не отмечается резких отклонений. Волокна располагаются довольно тесно. Между ними хорошо выявляются тонкие прослойки эндомизиума. Поперечная исчерченность волокон всюду хорошо проявляется.
Вторая зона изобилует кристаллами водной фазы самой разнообразной формы и величины. Они располагаются как между волокнами, так и внутри них. Кристаллы между волокнами обнаруживаются и в третьей зоне, но здесь их значительно меньше.
В охлажденном мясе, замороженном при -20°С, под микроскопом выявляются описанные выше три зоны. Однако охлажденное мясо промерзает, по-видимому, несколько иначе. Это проявляется в некоторых структурных особенностях зон замороженного мяса.
Первая поверхностная зона в охлажденном мясе, взятом в замороженном состоянии, несколько больше. Толщина этой зоны 300-400 мкм. Она состоит из компактно расположенных волокон. Толщина второй зоны 350-1300 мкм. В ней волокна в значительной степени деформированы. Третья зона более сходна с третьей зоной парного мяса, взятого в замороженном состоянии. Однако в охлажденном замороженном мясе волокна этой зоны тоньше (в среднем 29,7 мкм), тогда как в парном мясе, взятом в замороженном состоянии, их толщина 33,6 мкм
Таким образом, гистологический анализ устанавливает закономерный ход процесса замораживания. Это выражается в образовании трех структурно разных зон, начиная с поверхности замораживаемого отруба мяса.
Структурные зоны, возникающие в процессе замораживания, при хранении мяса влияют на его вкусовые качества.
В зависимости от режима замораживания можно изменять структуру зон и тем самым улучшать органолептические показатели мяса.
Закономерность правильного расположения зон в замороженном мясе длиннейшего мускула спины определяется совокупным действием при замораживании кинетики теплового обмена и упорядоченными движениями тканевой и вымерзающей воды, связанной с тканями.
В большом поясничном мускуле тоже много кристаллов между волокнами, но они лежат не между отдельными волокнами, а между их пучками. Кроме того, форма кристаллов значительно более гладкая и ровная.
Эти особенности структуры мышечных волокон становятся более четко выраженными через 3-9 мес. хранения. В это время кристаллы между волокнами становятся особенно крупными.
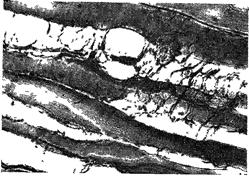
Рис. 103 Структура поперечных анастомозов между волокнами в мясе после 6 мес. хранения (по Писменской).
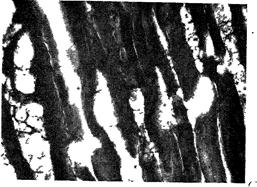
Рис. 104 Разная форма волокон в мясе после 6 мес. хранения.
Наряду с этим через 6 мес. хранения резко проявляется новый морфологический тест, представленный четкими поперечными анастомозами между отдельными волокнами. Такие анастомозы представляют собой совокупность кровеносных капилляров и нервных волоконец.
Проходя над обширными светлыми полостями, в которых были кристаллы, анастомозы отчетливо проявляются, создавая характерную поперечную сеть (рис 103). Четкость поперечных анастомозов позволяет довольно точно определить срок хранения мяса.
Такой тест анастомозов годен для мяса длиннейшего мускула спины, так как в большом поясничном мускуле он проявляется значительно слабее. Это объясняется тем, что в большом поясничном мускуле кристаллы тоньше и не образуют широких просветов между отдельными волокнами. Кроме того, сами волокна в этом мускуле тоньше, и анастомозы между ними менее мощные.
В мясе через 6 и 9 мес хранения белково-сывороточный материал иногда плотной массой заполняет все просветы, где располагались кристаллы. Эта зернистая масса слабее выражена и более рыхлая в большом поясничном мускуле
В связи с сильной кристаллизацией между волокнами и внутри волокон значительно меняется общий вид структуры замороженных мышц на продольных и поперечных срезах. В длиннейшем мускуле спины на поперечных срезах чаще выявляются картины разобщенных групп и сильно деформированных одиночных волокон (рис. 104). Деформации их вызывается острыми гранями кристаллов. Крупными овальными кристаллами внутри создаются кольцевые фигуры.
Иная картина наблюдается на поперечных срезах мышечной ткани большого поясничного мускула: волокна расположены плотными, слабо разобщенными пучками, кольцевых структур почти не образуется в связи с отсутствием крупных кристаллов в волокнах. Структура мышечных волокон на поперечных срезах интересна еще и тем, что дает дополнительный тест по определению срока хранения мяса.
На поперечных срезах замороженного мяса длиннейшего мускула спины при кратковременном хранении видны рыхло расположенные волокна полигональной формы.
При длительном хранении (после 6 мес.) часто обнаруживаются волокна серповидной и кольцевидной формы (см. рис. 104). Они возникают под действием крупных кристаллов, расположенных между и внутри волокон. Подобных кристаллов при кратковременном хранении (до 3 мес.) не образуется.
В мороженом мясе установлено существование трех структурных зон, которые возникают на основе разного воздействия холода на поверхностные и более глубинные слои мускула или отруба.

Рис. 105. Разные форма и величина льдинок из мороженого мяса.
Итак, установлено, что при замораживании мяса его поверхностные и более глубокие слои выявляют разную реакцию на воздействие холода, а в связи с этим в них образуется и совершенно различная структура. Это четко выражается в образовании трех структурно разных зон (рис. 102).
Закономерность правильного расположения зон в мороженом мясе в значительной степени зависит от правильного расположения волокон, параллельно к фасции. В тех случаях, когда волокна располагаются под углом, правильность расположения зон нарушается.
Если в препарат попадает одна из зон, то по ее структуре о морфологии всего мускула или данного отруба мяса судить нельзя. Для анализа следует брать пробы, по крайней мере, из 3-4 участков отруба, если было неизвестно направление действия холода.
Для точной оценки надо знать, какая часть мускула или отруба взята для анализа -периферийная, средняя или глубинная. Следует также учитывать направление волокон в замороженном мускуле, структуру расположения волокон в нем.
Таким образом, процесс воздействия замораживания па структурные элементы мяса - сложный. Общая направленность его едина для замороженного мяса, взятого как в парном, так и в охлажденном состоянии, за исключением некоторых различий в деталях.
Общий вид волокон иногда заметно изменяется в связи с тем, что поперечная исчерченность в них проявляется хуже. В процессе хранения она резко ослабляется в длиннейшем мускуле спины, особенно при замораживании после охлаждения, значительно меньше выявлена в большом поясничном мускуле.
Все волокна после замораживания становятся тоньше.
Особенностью при замораживании мяса является интенсивная кристаллизация водной фазы мяса в промежутках между отдельными волокнами. Общий вид кристаллов в разных мускулах различен. Так, кристаллы между волокнами в длиннейшем мускуле спины даже через 1 сут после замораживания получаются крупные, длинные и с неровными краями. Подобные кристаллы сильно деформируют расположенные с ними по соседству мышечные волокна, поэтому в замороженном мясе длиннейшего мускула спины часто обнаруживаются фрагментированные волокна с сильно изрезанными и изуродованными краями.
Закономерности кристаллизации в мясе при замораживании. Скорость замораживания влияет на ход кристаллообразования, на количество, размеры кристаллов льда и на характер их распределения в тканях. Форма и размеры кристаллов в мороженом мясе разнообразны (рис. 105). В большинстве случаев форма кристаллов неправильная, полигональная и нередко с острыми боковыми гранями.
Выше отмечалось, что даже через 1 сут замораживания как парного, так и охлажденного мяса намечаются три зоны, в каждой из которых кристаллизация протекает по-своему.
В поверхностной зоне кристаллы обычно встречаются редко. Во второй средней зоне они появляются как между волокнами, так и внутри, причем между волокнами они крупнее. В волокнах кристаллы относительно мельче, короче и чаще имеют овальную или круглую форму. Крупные кристаллы, располагающиеся между волокнами, имеют угловатую, причудливо изрезанную форму, часто с острыми краями. Поэтому они сильно деформируют и фрагментируют окружающие мышечные волокна, создавая разрыхлённую структуру мяса.
В третьей зоне процессы кристаллизации осуществляются главным образом между волокнами, но не бурно, чем во второй зоне. Кристаллы более гладкие, ровные и мельче, чем во второй зоне.
В парном мясе, замороженном в виде кусочков при -50ºС, в поверхностной зоне всюду обнаруживается энергичная кристаллизация в основном в волокнах. На продольных срезах кристаллы в волокнах нередко выявляются в виде четок (рис. 106).

Рис. 106. Процесс кристаллизации внутри волокон, входные или выходные канальцы, связанные с кристаллами (по Тинякову, Писменской, Куликовой).
Кристаллизация в волокнах характеризуется образованы выходных или входных канальцев, идущих в кристалл (рис. 105). Не исключена возможность, что по этим канальцам происходит миграция водной фазы.
Кристаллы, возникают при замораживании, нанос структуре мышечных волок повреждающий чисто механический эффект.
Кристаллы, расположенные в волокнах, прежде всего разрыхляют, а затем по мере роста спрессовывают отдельные пучки миофибрилл, оттесняя их к сарколемме. Кроме того, формируя внутри волокна каналы, они повреждают всю структуру волокна. В связи с этим в волокнах, содержащих кристаллы, раньше начинает исчезать поперечная исчерченность.
Вместе с тем рост кристаллов тесно связан со сложнейшими физико-химическими и биохимическими процессами, происходящими в мясе в период раннего автолиза, замораживания и хранения. В процессе замораживания и хранения связанная внутриклеточная вода отнимается от клеточных структур и перемещается. Отсюда и формирование канальцев в области расположения кристаллов. Следует подчеркнуть, что характер распределения кристаллов, их количество, форма, размер, структура и связанная с ними степень разрушения морфологических элементов мышечных волокон и других частей мяса в основном зависят от режимов и способов замораживания.
СТАДИИ АВТОЛИЗА МЯСА, ОПРЕДЕЛЯЕМЫЕ ПО ГИСТОЛОГИЧЕСКИМ ПРИЗНАКАМ
Сердечные мышечные волокна свиньи отличаются от волокон крупного и мелкого рогатого скота: в них ядра расположены тесной цепочкой. Мышечные волокна свиней в полтора раза толще мышечных волокон крупного…Рис. 107. Крупные узлы сокращения в волокнах кита-кашалота.
Поперечнополосатую мышечную ткань кита-сейвала трудно отличить от такой же ткани крупного рогатого скота, свиней и кита-кашалота. Однако в волокнах кита-кашалота имеются крупные и весьма характерные узлы сокращения (рис. 107), которых у кита-сейвала, коров и свиней нет.
Толщина мышечных волокон 15-200 мкм. В одной и той же мышце могут встречаться волокна разной толщины. Так, диаметр поперечно полосатых мышечных волокон длиннейшего мускула спины свиньи (порода крупная белая) 20-112 мкм (в среднем 62 мкм). На продольных срезах мускула часто обнаруживаются картины параллельного хода мышечных волокон, а иногда-резко выраженной гофрированности или волнистости волокон. Долгое время причины возникновения подобной гофрированности оставались не совсем ясными. Оказалось, что мышечные и соединительнотканные волокна крупного рогатого скота, зафиксированные через полчаса после убоя животного, расположены прямолинейно. По данным В. М. Макаева, волнистых волокон встречается мало - до 3%. Все мышечные волокна в срезах выявляют четкую, но разную поперечную исчерченность (рис. 73, а, б, в). Большинство волокон обладает сравнительно, мелкой исчерченностью, длина саркомера 2,3 мкм (рис. 73, б). Около 10% волокон имеют широкие темные и светлые диски, в них легко выявляются полоски Z и Н, длина саркомера 2,7-3,3 мкм (см. рис. 73, а). Обнаруживаются волокна и с более тонкой поперечной исчерченностью, длина саркомера 1,5-1,25 мкм (рис. 73, в).
В некоторых волокнах встречаются узлы сокращения. В них наблюдается характерная структура, состоящая из закономерно расположенных участков, разнокачественной поперечной и продольной исчерченности.
Мышечная ткань через 3 ч после убоя имеет такое же строение, что и через. 0,5 ч. Поперечная исчерченность в большинстве волокон обнаруживает структуру типа 4 (рис, 73, б). Встречаются волокна с мелкой поперечной и с продольной (длина саркомера 1,25 мкм) и только с продольной исчерченностью. Волокна типа 1 и 2 (рис. 73, а и б) бывают редко и в единичных экземплярах. Волнистые и гофрированные мышечные волокна обычно четко выявляют исчерченность типа 3 (рис. 73, б).
Через 12 ч после убоя увеличивается количество волокон с продольной исчерченностью, соответствующей типу 4 (см. рис. 73, е). Кроме того, встречаются гофрированные и волнистые волокна, в которых наблюдаются переходные состояния, т. е. мелкая поперечная исчерченность заменяется выраженной продольной. Такие же переходы наблюдаются и в прямолинейно расположенных волокнах. Узлы сокращений через 12 ч после убоя встречаются чаще, а в некоторых местах - группами по 2-3 узла.
Соединительнотканные волокна в прослойках обычно имеют причудливо извилистые очертания.
Через 24 ч после убоя большинство мышечных волокон выявляет продольную исчерченность, диаметр их увеличивается примерно в 2 раза по сравнению с волокнами типа 1 (рис. 73, а), имеющими длину саркомера 2,7-3,3 мкм. Узлов сокращений обнаруживается меньше, чем в срезах через 12 ч после убоя, а волокон с поперечными разрывами становится больше. Соединительнотканные волокна в прослойках парного мяса более извилисты по сравнению с образцами, взятыми через 3 и 12 ч после убоя.
Мышечные волокна через 48 ч после убоя в большинстве случаев обладают продольной исчерченностью, но некоторые из них имеют и поперечную - структура волокна принимает вид сетки. Узлы сокращений встречаются редко, но разрывов в волокнах еще больше, чем в предыдущей пробе. Наряду с волнистыми волокнами, обладающими поперечной исчерченностью, часто встречаются гофрированные волокна с продольной исчерченностью.
Через 72 ч после убоя наряду с мелкой поперечной исчерченностью (длина саркомера 1,25 мкм) встречаются участки со средней (длина саркомера 1,5 мкм) и с крупной (длина саркомера 2 мкм) поперечной исчерченностью. Это свидетельствует о начавшемся разрешении периода посмертного окоченения. Соответственно и волокнистый материал в прослойках эндомизиума приобретает менее извилистый характер. Узлов сокращений уже почти не обнаруживается.
Таким образом, длительный период автолиза, приводящий к созреванию мяса, складывается из ряда последовательных стадий, имеющих разное морфологическое выражение.
Первая стадия расслабленного состояния мышц начинается сразу после убоя; она характеризуется наличием большинства прямых, параллельно расположенных мышечных волокон Волнистых или гофрированных волокон очень мало.
Вторая стадия-начало окоченения - обнаруживается через 3 ч после убоя. Большинство мышечных волокон в этой стадии прямые, однако многие приобретают более мелкую поперечную исчерченность [тип 3 (рис. 73, в)], свидетельствующую о развитии сокращений. В этой стадии чаще начинают встречаться волокна с гофрированной структурой.
Третья стадия - развитие окоченения - начинается примерно через 10-12 ч после убоя и характеризуется четкими морфологическими признаками: большинство мышечных волокон обнаруживает только поперечную исчерченность типа 3 (рис. 73, в); многие волокна теряют поперечную исчерченность и приобретают продольную типа 4 (рис. 73, г); появляется значительное число гофрированных, или складчатых, волокон, приобретающих такую структуру под воздействием соседних волокон с наибольшим сокращением; соединительнотканные волокна в прослойках приобретают причудливо извилистый характер; обнаруживается много узлов сокращений, иногда по нескольку в одном волокне. Кульминационного развития третья стадия посмертного окоченения достигает к 20-24 ч после убоя.
Четвертая стадия - разрешение - обычно начинается через 2-3 сут после убоя. В это время многие мышечные волокна приобретают мозаичную структуру и становятся дискретными образованиями, так как по своей длине выявляют разную степень сокращения. Наряду с сильно сокращенными участками обнаруживаются районы расслабления с саркомерами длиной до 2 мкм. Узлы сокращений в это время почти не возникают, но наблюдается большое количество поперечных разрывов в волокнах.
Обычно узлы сокращений представляют сложную мозаику миофибрилл в разной степени сокращения. Чаще с одной стороны узла наблюдается почти полное расслабление (рис. 73), затем следует переход к более сильной степени сокращения [тип 3 (рис. 73, в)]. В центральной части узла иногда заметна полоса аморфного вещества, ближе к другому концу узла видна зона наибольшего сокращения, характеризующаяся продольной исчерченностью [тип 4 (рис. 73, г)]. Она переходит в зону меньшего сокращения с мелкой и более крупной поперечной исчерченностью.
Узлы сокращения - это предшественники расчленения мышечных волокон, которые имеются непосредственно в зоне максимального сокращения узла (рис. 88). Расчленение по узлам сокращения иногда происходит сразу в 2 - 3 местах. Тогда возникают короткие фрагменты волокон (рис. 87). Их положение среди других цельных волокон указывает на то, что разрывы произошли давно, быть может еще при жизни животного. Следовательно, узлы сокращения - это естественная структура, которая может создаваться не только в переживающих мышечных волокнах, но и при жизни животного. Таким образом, в результате развития ферментативных процессов физиологический аппарат субмикроскопического сокращения, т. е. тонкая структура актомиозинового комплекса, после максимума сокращения разрушается.
Следовательно, с морфологической точки зрения созревание мяса в первых стадиях обусловливается нарушением субмикроскопического аппарата сокращения. Оно сопровождается длительным процессом возникновения разных степеней сокращения и расслабления мышечных волокон, образования узлов сокращения, поперечных разрывов по узлам и продольных разъединении волокон. Мясо становится нежным, качество улучшается. В стадии полного окоченения мясо жесткое, и качество его низкое.
ВЛИЯНИЕ АВТОЛИЗА НА ВЕЛИЧИНУ ПОПЕРЕЧНОПОЛОСАТЫХ
МЫШЕЧНЫХ ВОЛОКОН
Latissimus dorsi 32-48; Longissimus dorsi 16-48; Infraspinatus 32-64;СОЕДИНИТЕЛЬНОТКАННЫЕ ЭЛЕМЕНТЫ В МЫШЕЧНОЙ ТКАНИ
Таблица 9 Часть туши Содержание, % коллагена эластина Шейная 3,11 0,1-1,1 … Определенное количественное соотношение коллагеновых и эластических волокон в… Чем старше животное, тем толще прослойки рыхлой соединительной ткани в его мышцах. По данным Зламаловой, у молодых…Рис 109. Жировые клетки среди мышечных волокон
При значительных скоплениях их мя получается лучшего вкуса и обладает большей питательностью (мраморное мясо).
В иных случаях встречаются мышцы, в которых жировой ткани становится очень много, и она начинает вытеснять волокна мышечной ткани. Такие мышечные волокна представляют собой перерождающиеся дегенерирующие волокна, уступающие место жировой ткани (рис 108).
Жировые клетки в соединительнотканных прослойках мышц на гистологических препаратах легко окрашиваются суданом III в ярко-желтый или оранжевый цвет. Они выявляются в виде округлых, овальных либо веретеновидных образований, более длинная ось всегда расположена по длине мышечных волокон (рис 109). Отложения ткани наблюдаются в жировой ткани наблюдается в мышцах грудной области, а также и в других у животных мясных пород.
ИЗМЕНЕНИЯ СТРУКТУРЫ МЫШЕЧНЫХ ВОЛОКОН В ПРОЦЕССЕ
ТЕХНОЛОГИЧЕСКОЙ ОБРАБОТКИ
Рис 110. Разрывы мышечных волокон в длиннейшем мускуле спины свиньи при посоле…Рис. 111. Складчатость сарколеммы в рассоле с циркуляцией
(по Тинякову и Большакову)
а - складчатость, видимая при увеличении микроскопа в 80 раз,
б - складчатость при увеличении в 400 раз
При посоле длиннейшего мускула свиньи в рассоле с циркуляцией (частота вращения 0,24 мин) наблюдается максимальная сжимаемость мышечных волокон (средний диаметр 43 мкм), примерно у половины волокон возникает грубая поперечная складчатость (рис 111). Сарколемма в таких волокнах имеет складки и собирается поперечным утолщением тяжами (рис 111).
В некоторых случаях обнаруживаются мышечные волокна, ярко окрашивающиеся суданом III, в которых поперечная исчерченность исчезает, а выявляются отдельные миофибриллы. Среди пучков миофибрилл хорошо заметны мельчайшие капельки жироподобного вещества, окрашенные суданом III в ярко-желтый или бурый цвет. При большом увеличении окрашенные капельки придают всему волокну мозаичную структуру, а при малом увеличении эти волокна выглядят однотонно окрашенными темно-бурыми нитями. Особенно хорошо они видны на поперечных срезах, легко установить их относительное количество На рис 112 видно, что почти в каждом из пучков мышечных волокон, разделенных прослойками внутреннего перемизия, имеется 2-3 волокна, окрашивающихся Суданом III Иногда подобных волокон встречается до 15%
Итак, установлено, что при посоле длиннейшего мускула свиньи поперечная исчерченность мышечных волокон может хорошо сохраняться в течение 40 дней. Но она не может служить точным показателем качественных изменений мяса в процессе посола.
Замораживание и размораживание. В процессе замораживания строение мышечных волокон нарушается. Эти нарушения вызываются кристаллами льда, которые образуются при замерзании воды, содержащейся в мышечных волокнах и между ними.
При 0,5-5°С создается максимальное количество кристаллов льда. При -18°С замерзает больше 75% воды, находящейся в мясе. Полное замерзание всех растворов мяса (температура эвтектики) наступает только при -56° С.
Форма кристаллов и место их образования зависят от скорости замораживания. Размер кристаллов льда обратно пропорционален скорости замораживания. При медленном замораживании формируются крупные кристаллы, но они немногочисленны. Такие кристаллы сильнее повреждают окружающую их ткань. При быстром более резком замораживании возникает значительное число мелких кристаллов внутри клеток, внутри поперечнополосатых мышечных волокон и между ними. Такие кристаллы не разрушают тканей

Рис 112 Мышечные волокна с жировым перерождением
По данным Постольски, длина крупных кристаллов 5-10-4 см, а ширина 1-10-4 см. Длина мелких кристаллов 5-10-6, ширина 1-10-4 см, Хайнер, Мэдсен и Хенкинс установили, что кристаллы льда при медленном замораживании говядины возникают в промежутках между волокнами, а при быстром - непосредственно внутрь волокна.
Процесс замораживания также вызывает определенные структурные изменения мышечной ткани. При повторном замораживании и размораживании изменяются концы мышечных волокон, при повторном образовании кристаллов разрушаются волокна и наступает дегидрация. Соединительная ткань эндомизия не изменяется.
В жидкости, вытекающей из размораживаемого мяса, Моал обнаружил частички разрушенных миофибрилл. Он установил также разрывы в мышечных волокнах.
Измельчение и сушка. При измельчении мышечная ткань сильно изменяется и разрушается. В первые же минуты разрушаются миофибриллы, пре вращаясь в аморфное вещество фарша. В готовом фарше под микроскопом видны масса аморфного более светлого вещества, отдельные сохранившиеся пучки мышечных волокон, жировые клетки, вкрапленные в аморфное вещество, и капли жирового вещества из разрушенных жировых клеток. В аморфном веществе фарша часто встречаются довольно крупные микропустоты овальной и округлой формы - это места, где были воздушные пузырьки, возникающие в процессе обработки фарша на куттере. Фарш, изготовленный из передних частей туши, грубее, чем из задних отрубов. В нем под микроскопом выявляют значительно больше участков с соединительной тканью. Мясо в процессе сушки значительно изменяется. Мышечные волокна имеют волнистость и значительное количество продольных трещин. Продольная и поперечная исчерченность исчезает, размер волокон уменьшается.

Рис 113 Мясная мука (по Бем и Плева)
1-мышечные волокна; 2-кусочки кости, 3-обрывки соединительной ткани.
При лиофилизации мышечные волокна расположены свободно, отделяясь одно от другого свободными пространствами. Поперечная исчерченность обычно сохраняется, окрашиваемость ядер в волокнах хорошая. Высушенная таким способом мышечная ткань при регидратировании почти полностью восстанавливается после пятиминутной выдержки в воде мясо имеет исходный свежий вид.
В мясной муке (рис 113) обнаруживают до 70-90% из мельченных мышечных волокон, а также аморфное вещество, костную и жировую ткани и инородные примеси. Соединительные элементы представлены в виде клейких частичек желтовато-коричневого цвета. Поперечнополосатые мышечные волокна в мясной муке сохраняют исчерченность, ослабленном виде.
Мясокостная мука содержит значительно меньше мышечной ткани, а больше аморфного вещества и высохшей крови. В ней встречается много волокон, песка и элементов растительного порядка.
Тема 8. Кожа и её производные.
Особенности строения некоторых органов пищеварения.
1. Особенности гистологического строения кожи у млекопитающих.
2. Микроскопические изменения, возникающие в процессе предварительной обработки кож на мясокомбинатах.
3. Понятие о строении слизистой оболочки пищеварительного тракта. Строение стенки кишок, являющихся сырьем для колбасного производства.
4. Печень, поджелудочная железа.
Вопрос 1. Особенности гистологического строения кожи у млекопитающих.
Клетки поверхностного слоя постепенно отмирают и слущиваются, а на смену им появляются новые в результате деления клеток более глубоких слоев… Дерма - наиболее толстый и плотный слой соединительной ткани, пронизанный… Подкожный слой, связывающий кожу с глубже лежащими частями тела, состоит из рыхлой соединительной ткани, в состав…Гистологическая структура кожи животных
Кожа развивается из эктодермы и мезенхимы. Эктодерма дает начало наружному слою кожи, или эпидермису (рис. 49, а, б, в, з), а мезенхима,… Затем его толщина и количество слоев постепенно увеличиваются, но ко времени…Эпидермис
Рис.49. Кожа мякиша лапы кошки(микрофото) а - роговой, б - блестящий, в - зернистый и г - шиповатый слои эпидермиса; д - сосочковый и е - сетчатый слои дермы, ж…Рис. 50. Схема строения кожи грубошерстных овец (по Диомидовой).
а - эпидермис, б - дерма, в - потовая железа, г - сальная железа, д - первичный и е - вторичный фолликулы
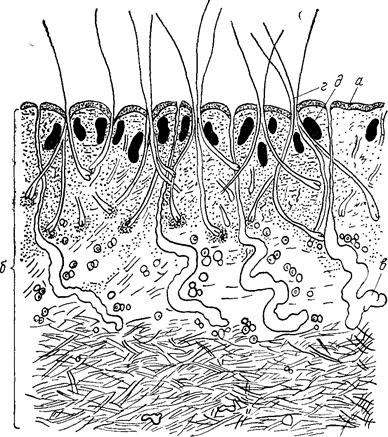
Рис. 51. Схема строения кожи тонкорунных овец (по Диомидовой):
а - эпидермис; б - дерма; в - потовая железа; г - волос; д - сальные железы.
Осязательные, илисинуозные, волосы толстые, длинные и упругие. Концентрируются обычно возле губ, век, ноздрей. Как показывает название, они обладают наполненными кровью синусами, или полостями между слоями волосяной сумки, которая в таких волосах развита очень хорошо, как и сальные железы. В синуозных волосах у копытных между двумя слоями волосяной сумки проходит множество соединительно тканых перекладин, вследствие чего синус напоминает пещеристое тело. У хищных перекладин нет, и синус принимает форму кольца. У этих волос отсутствуют мышцы-подниматели волос.
Смена волос. Волосы растут только до определенной длины и, оставаясь в таком положении, сменяются. Если это происходит с волосяным покровом на обширной площади, то такую смену называют линькой. Различают смену волос возрастную, происходящую до половозрелого возраста, периодическую, связанную со временем года, и перманентную, которая происходит у животных беспрерывно. Линька начинается с прекращения кровообращения в сосочке и новообразования клеток в корне волоса, который отделяется от производящих его клеток (рис. 52, г}. Новый волос (б) отрастает в результате регенерации луковицы и волосяного сосочка, которые развиваются так же, как и в эмбриональный период. Когда новый волос достигнет места прикрепления мышцы, старый волос вследствие ее сокращения выталкивается и выпадает.
Кровеносные сосуды и нервы волос. Кровеносные сосуды проходят только в соединительнотканых частях корня волоса. В волосяной сосочек, как правило, проникает специальная веточка артерии. Волос получает нервы от ножных нервных сплетений (рис. 53). В волосяной сумке имеется две системы терминалей: частокольная из продольных волокон и циркулярная из кольцевых. В синуозных волосах этих волокон больше и они толще. Располагаются они главным образом на периферии волосяной сумки. По мере приближения к стекловидной оболочке они многократно ветвятся, истончаются и формируют довольно плотное и чрезвычайно запутанное сплетение. Мышцы, поднимающие волос, снабжены собственными симпатическими нервами.

Рис. 52. Смена волос в коже свиньи (по Ржаницыной)
а - волосяной сосочек, б - волосяная луковица, в - новый волос, г - старый волос, 8 - нервные волокна

Рис. 53. Иннервация волоса из кожи соска кобылы (по Кацнельсону)
а - волос, б - сальная железа, в - нервные волокна
Использование кожного покрова и его производных
Основную массу шкуры используют в легкой промышленности для получения меха и кожи. Кератиновые производные кожи в основном применяют для получения… Кератины являются полноценными белками (в отличие от белков соединительной ткани), что обусловливает их использование…ТОНКАЯ КИШКА
Слизистая оболочка тонких кишок (рис. 47) собрана в многочисленные кишечные складки, которые располагаются не совсем правильно и соответствуют… Эпителиальный слой (б) слизистой оболочки образован однослойным призматическим… Мышечная оболочка.Внутренний кольцевой слой (к) очень мощный, наружный продольный (л) – тонкий. Между ними залегает…ТОЛСТАЯ КИШКА
В толстых кишках пищеварительные процессы играют значительно меньшую роль, чем в тонких; здесь происходит интенсивное всасывание, главным образом… Наиболее просто развивается толстый отдел кишечника у хищных. У домашних…МОДУЛЬ III
Тема 9. Выведение и разведение животных
1.Народно-хозяйственное значение животноводства. Значение курса для подготовки инженеров-технологов мясной промышленности.
2. Понятие о конституции, интерьере и их связь с направлением продуктивности животных.
3. Понятие об индивидуальном развитии, породе животных и методах разведения.
4. Мясная продуктивность животных и факторы, влияющие на качество мяса.
Вопрос 1. Народно-хозяйственное значение животноводства. Значение курса для подготовки инженеров-технологов мясной промышленности.
Животноводство является важной отраслью сельского хозяйства, обеспечивающей население разнообразными продуктами питания, а легкую промышленность -… Молоко, мясо, яйца и продукты их переработки отличаются высокой калорийностью… Из боенских отходов, туш, непригодных в пищу, получают мясо-костную, костную и кровяную муку, а из некоторых органов и…Вопрос 2. Понятие о конституции, интерьере и их связь с направлением продуктивности животных.
Конституция
В истории животноводства было немало попыток разработать определенную классификацию типов конституции, но наибольшее признание получила…Рис. 10. Бык нежной рыхлой конституции
Животные нежного типа имеют мягкий шелковистый шерстный покров, тонкую эластичную кожу, тонкий костяк, тонкие ноги и небольшую легкую голову, часто повышенную возбудимость.
У животного грубого типа шерстный покров грубый, кожа толстая, скелет массивный (толстые конечности и шея, большая и тяжелая голова).
Животные плотного типа имеют плотную кожу, хорошо развитую плотную (твердую на ощупь) мускулатуру, прочный скелет.
У животных рыхлого типа кожа рыхлая с сильно развитой подкожной соединительной тканью, мускулатура пышно развита с большим количеством жировых прослоек. Животные этого типа спокойные, флегматичные.
Нежный плотный тип характерен для коров молочных пород, скаковых лошадей, кур пород яичного направления, некоторых пород тонкорунных овец. У животных этого типа повышенный обмен веществ, слабая способность к жироотложению.
Нежную рыхлую конституцию имеют животные специализированных мясных пород (рис. 10). У них хорошо развита мускулатура, но мышцы рыхлые. Эти животные скороспелы и при хорошем кормлении и содержании дают много нежного и вкусного мяса.

Рис. 11. Бык плотной грубой конституции
Грубый плотный тип (рис. 11) имеют некоторые животные молочных и мясо-молочных пород крупного рогатого скота (холмогорской и симментальской); породы лошадей (якутской) и овец (гиссарской), которых разводят в условиях, близких к естественным.
Грубый рыхлый тип конституции является, как правило, нежелательным, так как у животных этого типа понижен обмен веществ, они малоподвижны, невыносливы и неспособны давать много продукции. Такой тип чаще встречается у некоторых быков и хряков, а также у лошадей тяжеловозных пород.
У животных крепкой конституции здоровое и гармоническое телосложение, нормально развиты все системы и органы. Этот тип конституции был предложен академиком М. Ф. Ивановым в дополнение к описанным тинам.
Указанные типы конституции не всегда можно четко выделить у данного конкретного животного. Иногда трудно бывает найти и явное различие между разными типами.
Интерьер
Особо большое значение в обмене веществ и энергии, а значит и многих интерьерных параметров, имеют эндокринные железы. Железы внутренней секреции… Гормон щитовидной железы (тироксин) регулирует интенсивность обменных… Гормоны гипофиза воздействуют на разнообразные функции организма. При удалении гипофиза или недостаточной его функции…Вопрос 3. Понятие об индивидуальном развитии, породе животных и методах разведения.
Онтогенез
Индивидуальное развитие организма (онтогенез) начинается со времени оплодотворения яйцеклетки и образования зиготы и протекает на протяжении всей… Онтогенезом называется совокупность количественных и качественных изменений,… В онтогенезе организм претерпевает изменения роста и развития. Еще Ч. Дарвин в своей работе «Изменение домашних…ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ РОСТА И РАЗВИТИЯ ЖИВОТНЫХ
Для роста и развития сельскохозяйственных животных характерны неравномерность и периодичность. Сельскохозяйственные животные в большинстве своем… Внутриутробное развитие млекопитающих проходит следующие подпериоды - стадии… Зародышевый (эмбриональный) подпериод. В первые семь суток у крупного рогатого скота проходит стадия дробления зиготы.…ИЗМЕНЕНИЕ ПРОПОРЦИЙ ТЕЛА ЖИВОТНЫХ В ПРОЦЕССЕ РОСТА
Неравномерность развития сельскохозяйственных животных в различные периоды проявляется и в изменении с возрастом пропорций их телосложения. Первыми в этом направлении были исследования проф. Н. П. Чирвинского, который более тридцати лет (с 1886 г.) изучал вопросы роста животных. Дальнейшие работы проф А. А Малигонова обогатили зоотехнию новыми данными о неравномерности развития организма животного.
Рис. 7. Пропорции телосложения взрослой коровы и новорожденного теленка.
По данным Н. П Чирвинского, ребра в постэмбриональный период растут быстрее в 5 раз, чем пясть Остальные кости занимают промежуточное положение. Кости периферического скелета (кости конечностей) - плечевая, бедренная, большая берцовая, лучевая, локтевая, плюсневая и пястная - в постэмбриональный период растут значительно медленнее, чем кости туловища (осевой скелет)-позвоночник, ребра, грудная кость. В результате различий в росте массы отдельных частей скелета пропорции телосложения животных с возрастом существенно изменяются. Так как в утробный период интенсивнее растут кости периферического скелета, то молодняк рождается относительно высоконогим, с высокой крестцовой костью, узкогрудым, с широкой и короткой головой, неглубоким и узким туловищем (рис. 7).
ПОРОДА
Породойназывается достаточно большая группа животных общего происхождения, обладающая сходными анатомо-физиологическими и продуктивными качествами и определенными требованиями к условиям окружающей среды, которые передаются по наследству.
По существующей классификации породы делят на примитивные, заводские и переходные.
К примитивным относят породы, выведенные в условиях, близких к природным. На формирование таких пород большое влияние оказал естественный отбор. К примитивным породам относят некоторые породы лошадей и овец, разводимых в азиатской части СССР, якутский и монгольский скот и некоторые другие. Характерной особенностью этих пород является грубый плотный тип конституции, большая выносливость и неприхотливость к условиям содержания. Несмотря на сравнительно низкую продуктивность эти породы имеют большую ценность из-за прекрасной приспособленности к специфическим, очень суровым условиям жизни.
Заводские породы выведены в условиях хорошего кормления и содержания. Эти породы отличаются высокой продуктивностью, большим разнообразием и повышенной изменчивостью. Так, в черно-пестрой породе крупного рогатого скота имеются коровы, дающие 2000 и 23000-25000 кг молока за лактацию. Некоторые из них получили мировое распространение, например, черно-пестрая и герефордская породы крупного рогатого скота, крупная белая порода свиней, порода кур леггорн. Заводская порода может состоять из нескольких отродий - весьма больших групп животных, которых разводят в различных районах страны. Так, существуют уральское, сибирское, центральное (среднерусское) и прибалтийское отродья черно-пестрой породы.
Переходные породы занимают промежуточное положение между примитивными и заводскими. Путем улучшения условий содержания и кормления, тщательного отбора и подбора переходная порода может быть преобразована в заводскую, как, например, получилось с холмогорской и ярославской породами крупного рогатого скота, романовской породой овец.
Методы разведения
Чистопородное разведение - спаривание животных одной породы применяют в племенных хозяйствах, на молочных фермах, во многих овцеводческих… Скрещивание - это спаривание животных разных пород. Для создания новых пород… Поглотительное скрещивание применяют для постепенной замены местной малопродуктивной породы или большого массива…Рис 17. Схема поглотительного скрещивания
Вводное скрещивание применяется, когда в целом удовлетворительную породу надо улучшить по какому-то отдельному признаку, например по жирности молока. С этой целью в хозяйство, разводящее скот с высокими удоями, но низкой жирностью завозят быка джерсейской породы (жирность молока коров 5,5-6%). Помесей I поколения быка из хозяйства убирают и продолжают чистопородное разведение, а также спаривание между собой помесей желательного типа. Таким образом, у черно-пестрого скота жирность молока можно повысить с 3,3-3,4 до 4,0-4,5% без уменьшения продуктивности. Такое скрещивание также называют корректирующим, так как оно не меняет основных свойств улучшаемой породы.
При промышленном скрещивании полученных помесей первого поколения используют для откорма (в мясном скотоводстве, птицеводстве и свиноводстве). У помесей первого поколения проявляется гетерозис - явление повышенной жизнестойкости, скорости роста, лучшего усвоения корма. Биологическая сущность гетерозиса пока остается неясной. Одна из загадок гетерозиса заключается в том, что он проявляется только у помесей первого поколения.
Так, в свиноводстве для получения молодняка для откорма используют хряков и маток разных пород. Помесные поросята превосходят своих чистопородных сверстников по приросту живой массы на 10-15%, а на 1 кг прироста затрачивают кормов на 8-10% меньше. Цыплят, откармливаемых на мясо - бройлеров, также получают от скрещивания разных пород.
Гибридизация - спаривание животных разных видов. Ее применяют для использования гетерозиса (от спаривания осла с лошадью получают мула, который превосходит лошадь по работоспособности, неприхотливости, устойчивости к инфекционным заболеваниям) и выведения новых пород. Казахские овцеводы путем спаривания диких горных архаров с матками тонкорунных пород прекос и рамбулье вывели новую тонкорунную породу казахский архаромеринос, в которой соединились крепость конституции, приспособленность к содержанию в высокогорных районах с хорошим качеством шерсти и высокой шерстной продуктивностью. Путем спаривания дикого среднеазиатского кабана с матками крупной белой и кемеровской пород получена казахская гибридная порода свиней.
В США путем скрещивания шортгорнской мясной породы и индийского скота зебу выведена мясная порода крупного рогатого скота санта-гертруда, соединяющая в себе хорошую мясную продуктивность с приспособленностью к жаркому климату южных штатов.
Особенности племенной работы в современном животноводстве
Технология промышленного животноводства рассчитана на групповое безвыгульное содержание на щелевых полах в помещениях с регулируемым микроклиматом.… Ведется селекция на устойчивость животных к инфекционным заболеваниям, так как… Американский птицевод Хатт путем длительного отбора кур, устойчивых к белому бациллярному поносу (пуллорозу), создал…Вопрос 4. Мясная продуктивность животных и факторы, влияющие на качество мяса.
Таблица 2 - Химический состав и калорийность мяса животных различных видов средней упитанности (%) Мясо Сухие вещества … В состав туши, кроме мышечной, входят жировая, костная, соединительная ткани, хрящи и связки. Наилучшей считается…Тема 10. Кормление животных
1. Значение кормовой базы в повышении продуктивности животных.
2. Химический состав кормов.
3. Энергетическая, протеиновая, аминокислотная, витаминная и минеральная питательность кормов.
4. Растительные корма.
5. Корма животного происхождения и дрожжи.
6. Комбикорма.
7. Минеральные подкормки.
Вопрос 1. Значение кормовой базы в повышении продуктивности животных.
Для животноводства важно не только количество, но и качество кормов, которое определяется содержанием в них питательных веществ.Вопрос 2. Химический состав кормов.
Вода. Больше всего воды (60-80%) содержат сочные корма. Определить содержание воды в кормах можно путем высушивания их. Разность в массе корма до и…Рис. 1. Химический состав кормов
Сухое вещество. Сухое вещество состоит из органической и неорганической (минеральной) частей. Минеральные вещества или зола - это то, что остается от корма после его сжигания,
Органическая часть сухого вещества корма состоит из азотистых (протеина) и безазотистых веществ и витаминов.
Протеин включает в себя белки и амиды (небелковые азотистые соединения органического происхождения). Подобно жирам и углеводам белок содержит углерод, водород и кислород, но в отличие от них еще и азот.
Белки - один из основных элементов питания человека и животных, они служат источником «строительных материалов», из которых создаются клетки и ткани организма. Белки играют роль регуляторов обмена веществ. Из белков состоят некоторые гормоны. К белкам относят антитела, выполняющие в организме защитную функцию, и ферменты.
В белке нуждаются все животные, так как они не могут синтезировать его подобно растениям и микроорганизмам из небелкового азота. При недостатке или отсутствии белка в корме нарушается азотистый баланс в организме. Потребность организма в белке зависит от вида животного, его возраста и направления продуктивности. Молодняку необходимо давать повышенное количество белка, так как животные интенсивно растут, увеличивается масса всех их органов и тканей. Много белка необходимо животным, выделяющим его со своей продукцией - дойным коровам, курам, овцам. Без большого количества белка невозможен мясной откорм, так как сухое вещество мяса в основном состоит из белка.
Белки состоят из аминокислот. Разнообразие свойств растительных и животных белков определяется тем, как аминокислоты расположены в молекуле белка.
Синтез белков тела животного можно представить в виде такой упрощенной схемы: растительный белок - аминокислоты - животный белок. Некоторые аминокислоты белка корма могут быть перестроены в организме в другие аминокислоты, необходимые для животного.
Десять аминокислот (лизин, метионин и др.) животные должны получать в готовом виде, так как организм не может их синтезировать. Поэтому эти аминокислоты называются незаменимыми. Их недостаток тормозит синтез животных белков. К белкам, содержащим все незаменимые аминокислоты, относят белки молока, мясокостной и рыбной муки, гороха и сои. Они имеют высокую биологическую ценность.
В настоящее время промышленность выпускает синтетические незаменимые аминокислоты. При добавлении их к корму значительно улучшается использование белков, что позволяет сократить скармливание дорогостоящих кормов животного происхождения. Поросята и цыплята, получающие набор кормов, содержащий все незаменимые аминокислоты в нужном количестве, дают на 15-20% больше привес, чем те животные, которые получают рацион, несбалансированный по аминокислотам.
Различные корма сильно отличаются друг от друга по содержанию белка. Белком богаты мясокостная и рыбная мука, жмыхи и шроты, зернобобовые. В свекле и картофеле мало белков.
К безазотистым веществам относятся углеводы и жиры.
Углеводы составляют 50-70% сухого вещества корнеплодов, клубнеплодов и зерновых кормов. Следовательно, поедая растительные корма, животные получают огромное количество углеводов в виде Сахаров и полисахарида - крахмала. Но в теле животных углеводов очень мало (1-1,5%) и находятся они в основном в виде животного крахмала - гликогена. Это объясняется тем, что большая часть углеводов корма распадается в клетках с выделением энергии, необходимой для жизни животного. По своей энергетической ценности углеводы занимают третье место, но являются лучшим источником энергии, так как быстро всасываются и легко распадаются. При недостатке углеводов в корме снижается продуктивность животных и ухудшается их состояние, особенно дойных коров. Если животное получает углеводы в избытке, то их излишек превращается в жир. Эту способность животных превращать углеводы в жир широко используют при откорме свиней до жирных кондиций.
К углеводам корма относится также клетчатка - полисахарид, входящий в состав клеточных стенок растений, который не расщепляется ферментами пищеварительной системы. Чем выше ее содержание в корме, тем ниже его питательность. Однако она необходима в определенных количествах для нормального процесса пищеварения, особенно у жвачных животных.
Жиры в растительных кормах находятся в небольших количествах (2-3%); исключение составляют лишь семена масличных культур и продукты их переработки. Жир, находящийся в теле животного, играет роль своеобразного резерва энергии. При ухудшении кормления животное использует его в качестве источника энергии (за исключением цитоплазматического жира). Не случайно у животных, которых разводят в суровых естественных условиях (у овец, табунных лошадей, верблюдов), в теле откладывается большой запас жира. Имея очень низкий коэффициент теплопроводности, подкожный жир выполняет роль теплоизолятора, уменьшая отдачу тепла и предохраняя животное от охлаждения. Упитанное животное гораздо лучше переносит холод, чем худое и истощенное.
Детальные исследования показали, что при скармливании обезжиренных кормов замедляется рост, нарушаются половые функции и т. п. Поэтому животные должны получать с кормом определенное количество жира.
Минеральные вещества выполняют важные физиологические функции в организме. Различают две группы минеральных веществ: макроэлементы (кальций, фосфор, натрий и др.) и микроэлементы - железо, медь, кобальт и др. Общее содержание минеральных веществ в кормах не превышает 4-7%.
Кальций и фосфор необходимы животным в больших количествах, так как они входят в состав костной ткани. Кальций понижает возбудимость нервной системы, влияет на свертываемость крови. Много его в листьях и стеблях растений. Фосфор входит также в состав нуклеиновых кислот, играет большую роль в углеводном обмене. В растительных кормах (за исключением зерновых, отрубей, жмыхов и шротов) фосфора в 3-4 раза меньше, чем кальция.
В растительных кормах калия больше, чем натрия, но при правильном соотношении они обеспечивают нормальную работу сердца и пищеварительных органов. Калий влияет на работу сердца. Натрий повышает возбудимость нервной системы, регулирует водный обмен. Ионы натрия обладают способностью притягивать к себе воду. Крупному рогатому скоту и овцам поваренную соль можно давать вволю, а свиньи и птицы чувствительны к избытку натрия.
Магний входит в состав костной ткани, регулирует действие некоторых ферментов, участвует в процессах сокращения мышц. При его недостатке наступает расстройство сердечной деятельности, появляются судороги
Животные нуждаются в очень небольших количествах микроэлементов. Например, корове в сутки требуется всего лишь 6-15 мг кобальта, но его отсутствие или недостаток может вызвать серьезные расстройства в обмене веществ. Медь, железо и кобальт влияют на процесс кроветворения. Железо входит в состав гемоглобина. Марганец влияет на процессы размножения. Цинк входит в состав гормона поджелудочной железы - инсулина, а йод - в состав тиреоглобулина - гормона щитовидной железы. В кормах микроэлементы содержатся в очень небольших количествах. При недостатке микроэлементов их добавляют в корма. При избытке (молибдена, никеля, фтора и др.) в почвах некоторых районов России, а, следовательно, в кормах и воде у животных появляется целый ряд заболеваний.
Витамины - это органические вещества, содержащиеся в корме в очень небольших количествах. В XIX в. господствовало мнение, что для жизни человеку и животному необходимы белки, жиры, углеводы и минеральные вещества. В 1881 г. молодой русский ученый Н. И. Лунин провел на мышах следующий опыт: одна группа мышей получала натуральное коровье молоко, а другая - смесь из молочного сахара, молочного белка, растительного жира и минеральных веществ. Первая группа чувствовала себя нормально, другая довольно быстро погибла. Из этого опыта Н. И. Лунин сделал вывод о том, что помимо белков, жиров, углеводов и минеральных веществ в очень малых количествах нужны какие-то еще вещества, в данном случае содержащиеся в натуральном молоке. Эти вещества впоследствии были названы витаминами.
Отсутствие витаминов в кормах вызывает серьезные заболевания - авитаминозы. Но чаще мы встречаемся не с полным отсутствием витамина, а с его недостатком.
Основное значение витаминов заключается в том, что они входят в состав ферментов, без которых не может осуществляться обмен веществ. Свежая зеленая трава и некоторые другие корма содержат необходимые для животных витамины. Большинство витаминов разрушается при нагревании и длительном хранении, под действием света, кислорода, тяжелых металлов.
Витамин А - способствует росту клеток эпителиальной ткани. При отсутствии витамина А в кормах замедляется рост и развитие молодняка. У взрослых животных недостаток витамина А приводит к бесплодию. Явным признаком авитаминоза или гиповитаминоза А является воспаление роговой оболочки глаз. Авитаминоз А вызывает куриную слепоту - снижается острота зрения ночью.
В растительных кормах содержится только провитамин А - каротин, который в печени превращается в витамин А. Каротином богаты зеленая трава и морковь. Красно-оранжевый цвет моркови свидетельствует о высоком содержании в ней каротина.
Витамин D регулирует фосфорно-кальциевый обмен. Иногда у молодняка встречается заболевание, называемое рахитом. При рахите нарушается образование костной ткани. Из-за пониженного содержания кальция и фосфора кости теряют свою прочность, деформируются под тяжестью тела. Состояние молодняка резко ухудшается. При введении кальция и фосфора с кормом развитие болезни не прекращается, так как они не усваиваются организмом без витамина D. Поэтому его называют антирахитическим. Наиболее богат витамином рыбий жир.
Летом рахита, как правило, не бывает. Это связано с антирахитическим действием ультрафиолетовых лучей солнца. Под их влиянием находящееся в подкожном слое вещество - 7-дегидрохолестерин превращается в витамин D. Поэтому зимой животных необходимо облучать специальными лампами. Ультрафиолетовые лучи являются прекрасным средством предупреждения и лечения рахита. При ультрафиолетовом облучении кормов, особенно дрожжей, в них повышается содержание витамина В.
Витамин Е влияет на плодовитость животных. При авитаминозе Е может произойти оплодотворение, но во время беременности зародыши гибнут. У самцов нарушается образование сперматозоидов.
Витамин К - антигеморрагический. При недостатке его кровь плохо свертывается. У птиц происходят множественные кровоизлияния во внутренние органы.
Витамины группы В состоят из витаминов, совершенно различных по своему действию. Витамин В1 входит в состав фермента, регулирующего распад углеводов.
При авитаминозе В в организме накапливаются вещества, вызывающие воспаление нервных стволов. Это заболевание, называемое полиневритом, может привести к смерти. В витамине В1 нуждаются лошади, свиньи, кролики и особенно куры. Витамин В1 содержится в зерне, но особенно много его в рисовых отрубях и дрожжах.
Витамин В3 необходим для нормального роста и развития, регулирует процессы клеточного дыхания, оказывает специфическое влияние на яйценоскость кур.
Витамин В12 называют антианемическим. При его отсутствии у человека и животных развивается анемия, т. е. в крови резко уменьшается количество эритроцитов. В витамине В12 содержится кобальт.
В витамине С нуждаются лошади, свиньи и птицы; жвачные животные в нем не нуждаются, так как синтезируют его сами.
Добавка к основным кормам рациона незаменимых аминокислот, витаминов, микроэлементов и других биологически активных веществ значительно улучшает использование корма. Свиньи на 1 кг прироста потребляю! около 8 кг зерна. Если рацион сбалансирован по всем необходимым веществам для организма, то на 1 кг прироста затрачивается 3,5-4 кг зерна.
Вопрос 3. Энергетическая, протеиновая, аминокислотная, витаминная и минеральная питательность кормов.
Под питательностью кормов понимают свойство последних удовлетворять природные требования животных в пище. Оценивают питательность кормов по их… В состав кормов входят органические и неорганические вещества. К первым… О количестве переваримых веществ судят по разности между принятыми с кормом и выделенными с калом. Отношение величины…Питательность кормов в обменной энергии
За энергетическую кормовую единицу принята питательность 1 кг, а в птицеводстве 1 г зерна овса, которые содержат соответственно 2500 и 2,5 ккал… В табл. 8 приводится пример определения питательности сена люцернового в…Протеиновая питательность кормов
Протеины растительных кормов бедны лизином, метионином, триптофаном. Эти аминокислоты принято называть «критическими». Некоторые аминокислоты (аланин, глицин, пролин, аспаргиновая кислота и др.)… Таблица 9- Биологическая ценность протеина отдельных кормов Виды кормов Коэффициент использования…Азот усвоенный
Коэффициент использования азота = ------------------------------------ X 100
Азот переваренный
Недостаток некоторых аминокислот в одних кормах можно пополнить за счет других. Так, например, протеин кукурузы беден лизином, триптофаном и…Витаминная питательность кормов
В хозяйственных условиях обеспеченность животных витаминами зависит от сезона года в летний период она в основном нормальная, в зимний -… Биологическую активность витаминов определяют в интернациональных единицах… Потребности сельскохозяйственных животных в каротине удовлетворяются за счет скармливания зеленых кормов, сена,…Минеральная питательность кормов
Хотя минеральные вещества содержатся в организме в незначительном количестве, жизнь без них невозможна: они входят в состав многих сложных… Основная роль микроэлементов - в повышении активности различных ферментных… Особенно важны макро- и микроэлементы для молодняка, поскольку образование новых клеток без них немыслимо. Но и…Вопрос 4. Растительные корма
Зеленый корм - что трава естественных лугов и специально возделываемая для нужд животноводства. Важное биологическое значение травы объясняется… Для бесперебойного обеспечения животных травой на протяжении всего… У крупного рогатого скота и овец трава клевера и люцерны, особенно молодой и после дождя, вызывает тимпанит. Приучать…Вопрос 5. Корма животного происхождения и дрожжи.
Отходы молочной, мясной и рыбной промышленности содержат в своем составе много белков высокой биологической ценности, минеральных веществ и… Снятое молоко - ценный белковый продукт, богатый молочным сахаром и… Пахта. По содержанию белка, молочного сахара и общей питательности приравнивается к снятому молоку. Хороший корм для…Вопрос 6. Комбикорма.
Комбикорма-концентраты состоят в основном из зерна, отрубей, жмыхов, шротов, высушенной барды, пивной дробины, жома, рыбной и кровяной муки,… Качество комбикормов должно отвечать определенным рецептам, утвержденным… Полнорационные комбикорма должны содержать в своем составе все питательные вещества, по объему отвечать строению…Вопрос 7. Минеральные подкормки.
Поваренную соль используют для всех животных как источник натрия и хлора, которых не хватает в растительных кормах. В 1 кг соли содержится 367 г… Наиболее распространенными из фосфорно-кальциевых подкормок являются… Обесфторенный фосфат должен содержать не меньше 16 г фосфора, 34 г кальция и не больше 0,2 г фтора в 100 г. К…Тема 11. Скотоводство
1. Биологические особенности крупного рогатого скота и особенности пищеварения у жвачных.
2. Основные породы молочного (голландская, голштинская, черно-пестрая, холмогорская, красная степная, ярославская), двойного (симментальская, щвицкая) и мясного (шортгорнская, герефордская, шаролезская, казахская белоголовая, калмыцкая) направлений продуктивности.
3. Откорм скота.
Вопрос 1. Биологические особенности крупного рогатого скота и особенности пищеварения у жвачных.
Крупный рогатый скот лучше, чем другие виды животных, переваривает корма с высоким содержанием клетчатки. Благодаря синтезу аминокислот в… Крупный рогатый скот хорошо акклиматизируется в различных климатических…Особенности пищеварения в желудке жвачных
Желудок состоит из четырех крупных камер – рубца, сетки, книжки и сычуга. Первые три камеры называются преджелудочными и являются безжелезистыми… У некоторых животных (верблюд, лама, альпака) желудок трехкамерный (обычно… Слизистая преджелудков покрыта многослойным ороговевающим эпителием и имеет характерное строение в разных камерах: в…Состав и свойства желудочного сока
Неорганические вещества Йоны Na, K, Mg, HCO3, HPO4, SO4, Cl. Преобладающими ионами являются Na, Cl,… Роль HCl:Молочного направлений продуктивности
По сообщению П. Н. Кулешова (1947), голландская порола относится к древним породам Европы, послужившим к улучшению скотоводства до 1700–1750 гг. Как… Голландская порода выделилась из общей массы местного скота и послужила… Голландская порода скота имеет черно - пеструю масть, и поэтому его в большинстве стран мира так и называют…Двоенного направления продуктивности
Длительное время симменталов использовали как рабочий скот. В этот период при отборе животных в первую очередь ценились их рабочие качества. С… Среди скота симментальской породы встречаются животные двух типов:…Рис 28 Корова симментальской породы
Конституция симментальского скота крепкая, у части животных несколько грубоватая Голова тяжелая, широкая во лбу. Рога желтоватые, со светло-кopичневыми кончиками. Носовое зеркало, язык, зев и веки светли-розовые. Темные пятна на этих местах - свидетельство нечистопородности скота. Шея средней длины, мускулистая, холка широкая, у быков раздвоенная) Грудь глубокая и умеренно широкая, спина и поясница длинные, широкие и прямые, крестец приподнят, зад средней длины, широкий и хорошо омускуленный. Конечности средней длины, толстые, с хорошо развитыми суставами Признаки молочности выражены удовлетворительно. У ряда коров передние доли вымени развиты хуже задних («козье вымя»). Кожа толстая, тяжеловесная и рыхловатая (составляет 10-14% от живой массы). Масть животных палево-пестрая, красно-пестрая, палевая (рис. 28).
Симментальский скот - один из крупных. Масса коров - 550- 650 кг, быков - 850-1000 кг; наибольшая масса коров - 870 кг, быков - 1300 кг. Телята при рождении весят 36-45 кг и в условиях полноценного кормления в годовалом возрасте достигают 330- 390 кг. Мясные качества скота высокие. Масса откормленного молодняка в 1,5-2,5 года составляет 500-700 кг, убойный выход откормленного молодняка 56-58%, а хорошо упитанного взрослого скота - до 65%. Мясо у симменталов высокого качества, мраморное. Однако в туше относительно много костей: даже у хорошо откормленного молодняка кости составляют 18-19% массы туши.
По данным ГПК, удои коров досгигают 3700-4100 кг, однако в передовых хозяйствах они значительно выше - до 4500-5000 кг. Среднее содержание жира в молоке (в зависимости от типов) 3,7-4,0%, белка - 3,3-3,6%.
Швицкая порода. Выведена в Швейцарии путем длительного отбора животных по молочной и мясной продуктивности в хороших условиях кормления и содержания.
Рис. 9. Корова швицкой породы
Телосложение швицкого скота типично для животных молочно-мясного направления продуктивности, конституция крепкая Голова средней величины, с широким и коротким лбом, рога светлые с темными кончиками; шея средней длины, мускулистая, подгрудок развит умеренно. Туловище длинное, грудь глубокая и широкая, линия верха ровная, зад средней длины, прямой, широкий и хорошо омускуленный. Признаки молочности хорошо развиты Конечности короткие, правильно поставленные и крепкие. Масть бурая, с оттенками от светло-бурой до темно-бурой Породный признак животных швицкой породы - темное носовое зеркало со светлым кольцом по окружности, светлый ремень вдоль спины и более светлая окраска волос на внутренней стороне ног, вымени и внутри ушных раковин (рис. 29). Помимо Швейцарии, швицкий скот распространен в ряде соседних с ней стран и в США.
В Россию швицев стали завозить в начале XIX в. Благодаря хорошей акклиматизации, высокой молочной и мясной продуктивности швицкий скот широко использовался в качестве улучшающей породы. Путем воспроизводительного скрещивания местного скота со швицким в СССР созданы зональные породы скота молочпо-мясного направления: костромская, лебединская, алатауская, бурая карпатская, кавказская бурая.
Масса коров этой породы составляет 480-550 кг, быков - 800- 950 кг и более. Удои молока в племенных стадах от 3500 до 5000 кг на фуражную корову в год, содержание жира от 3,8 до 4,1% и белка - 3,46-3,56%.
Мясные качества высокие: при интенсивном выращивании и откорме бычки в 17-18-месячном возрасте имеют массу 450-500 кг, убойный выход откормленных животных - 55-60%.
Мясного направления продуктивности
шортгорнская,
герефордская,
шаролезская,
казахская белоголовая,
калмыцкая
Мясные великорослые породы отличаются так называемой великорослой скороспелостью. Они быстро растут» достигая в раннем возрасте большой массы, но позже заканчивают развитие. Прирост массы у великорослого мясного скота происходит в основном за счет роста мышечной ткани. Процесс интенсивного жироотложения начинается в значительно более позднем возрасте, чем у животных скороспелых мясных пород. Благодаря этому скот великорослых мясных пород на единицу прироста массы расходует меньше кормов, а мясо его менее жирное, чем у скота сокороспелых мясных пород.
Шортгорнская порода. Выведена на северо-востоке Англии в XVIII в. Здесь, в районе долины р. Тис, еще в XVI в. разводили скот, отличавшийся большой живой массой, хорошим развитием мускулатуры и рабочими качествами. В XVII и XVIII вв, сюда завозился голландский скот. На базе местного и голландского скота и была создана шортгорнская порода (в переводе - «короткорогая»). Племенная работа при выведении шортгорнского скота велась в направлении улучшения мясных качеств, а затем, с увеличением спроса на молоко,- на повышение молочности. В шортгорнской породе два направления продуктивности: мясное и молочно-мясное. В настоящее время шортгорнскую породу в Англии, США и Канаде разводят в основном как мясную.
Шортгорны мясного направления имеют нежную рыхлую конституцию, голова небольшая, широкая во лбу, рога небольшие (сейчас создаются отродья безрогих мясных шортгорнов); шея короткая и толстая, туловище компактное, широкое и глубокое, с очень хорошо развитой мускулатурой, особенно в задней части; костяк тонкий, но довольно крепкий, кожа мягкая и рыхлая.
Экстерьер шортгорнов молочно-мясного направления не отличается от экстерьера скота современных молочных заводских пород. Масть красная, чалая, реже белая. Скот крупный: масса коров мясного типа составляет 570-650 кг, быков - 850-950 и до 1250 кг. Шортгорнская порода высокоскороспела. Животные заканчивают развитие к 3-4 годам. Масса телят при рождении 27-32 кг, они быстро растут. При откорме молодняк к 18-месячному возрасту достигает 450-480 кг. Мясо нежное, равномерно пронизанное жировыми прослойками, сочное и вкусное, убойный выход - 65-72%.

Рис 30 Корова герефордской породы
В Россию шортгорнский скот впервые был завезен в XIX в. Впоследствии в Курганской области улучшенный скот послужил основой для создания отечественной курганской породы мясного направления. В условиях России шортгорнский скот довольно хорошо акклиматизируется, но требователен к условиям кормления и содержания. В настоящее время разводят шортгорнов «в чистоте», а также используют их для улучшения отечественных мясных пород скота и в промышленном скрещивании. По данным ГПК, масса чистопородных шортгорнских коров молочно-мясного направления составляет 480-570 кг, удой - 2500-3250 кг молока жирностью 3,7-3,9%.
Герефордская порода. Выведена в XVIII в. в Англии в графстве Герефорд путем отбора по мясным качествам среди местного рабочего скота Животные этой породы имеют крепкое здоровье, хорошо акклиматизируются, приспособлены к пастбищном} содержанию, легко переносят перегоны на большие расстояния
Масть герефордов красная, с белым подгрудком, нижняя линия живота, нижняя часть конечностей и кисть хвоста также белые. По экстерьеру современный герефордский скот представляет животных типичного мясного склада с хорошо развитой мускулатурой и широким туловищем на коротких и прочных ногах (рис 30). Масса коров составляет 500-580 кг, быков - 800-900 кг Мясные качества скота высокие. При интенсивном выращивании и откорме уже к годовалому возрасту бычки достигают массы 400-420 кг, давая убойный выход 60-68% Мясо герефордов мраморное, нежное, высоких пищевых достоинств.
Шаролезская порода (шароле). Выведена во Франции путем длительного отбора местного скота по выносливости, скороспелости и мясным качествам. Раньше животные этой породы разводились не только для получения молока и мяса, но и как рабочий скот. Затем в связи со снижением потребности в живом тягле порода шароле совершенствовалась в мясном направлении. В середине XIX в. этой породе приливали кровь шортгорнов, а еще раньше - симменталов, но это не оказало большого влияния на формирование шаролезского скота.

Рис. 32. Бык породы шароле
Животные породы шароле имеют ряд особенностей, отличающих их от скота других мясных пород. Во-первых, они одни из самых крупных: масса коров составляет 780-1045 кг, быков-1100-1500 кг; во-вторых, молодняк обладает очень высокой энергией роста: в течение длительного времени среднесуточные приросты составляют 940-1300 г в сутки. При этом откладывается меньше жира и больше белка, благодаря чему от молодняка получают нежирную говядину, пользующуюся сейчас особенно высоким спросом. Вместе с тем в мясе откормленных животных этой породы в достаточной степени выражена мраморность.
Масть скота желтая с разными оттенками вплоть до белой. Голова короткая и широкая во лбу, шея мясистая, с несильно развитым подгрудком. Грудь широкая и глубокая, зад широкий, хорошо омускуленный. Кожа средней толщины, костяк довольно массивный. Передняя, средняя и задняя части туловища пропорционально развиты (рис.. 32). Животные выносливы и хорошо акклиматизируются в различных климатических условиях. Скороспелость высокая: телята рождаются массой 40-50 кг и к 6 месяцам достигают 210-260 кг. При интенсивном выращивании и откорме масса молодняка годовалого возраста достигает 450-500 кг, а в возрасте 18 месяцев - 610-620 кг. Убойный выход 60-70%.
В Россию животные породы шароле завозились на Украину еще до 1917 г Помеси от скрещивания этой породы с серым украинским скотом отличались очень высокими мясными качествами. Новый завоз скота этой породы начался с 1961 г. Сейчас порода шароле разводится в ряде центральных областей России и используется главным образом для промышленного скрещивания со скотом молочного и комбинированного направления продуктивности.
Казахская белоголовая порода. Выведена в период с 1930 по 1950 г в Казахстане и Нижнем Поволжье путем скрещивания калмыцкого и казахского скота с герефордской породой Скрещивание обычно проводилось до II-III поколений, а затем полней разводились «в себе» Животные этой породы сочетают ценные качества местного и герефордского скота. Наряду с крепким здоровьем, приспособленностью к условиям континентального климата и использованием сухих степных пастбищ они обладают достаточно высокой скороспетостью и имеют хорошие мясные качества. По масти и типу телосложения казахская белоголовая порода сходна с герефордской. Скот относительно некрупный, компактного телосложения, с крепким легким костяком, туловище округлое и широкое, с хорошо развитой мускулатурой. Коровы имеют массу 500- 550 кг, быки - 800-900 кг (рис 31). При нагуле без подкормки концентратами среднесуточные приросты у животных достигают 800-900 г. Убойный выход коров после нагула 52-58%, а хорошо откормленных кастратов - 63-67%, мясо сочное и достаточно мраморное. При интенсивном выращивании и откорме молодняк к 1,5 годовалому возрасту достигает живой массы 420-480 кг.
Рис 31 Бык казахской белоголовой породы
Сейчас порода совершенствуется в мясном направлении. Она разводится в Казахстане, Оренбургской и Волгоградской областях.
Калмыцкая порода. Калмыцкий скот попал на юго-восток европейской части нашей страны в начале XVII в. вместе с перекочевавшими сюда из Западной Монголии калмыцкими племенами. Создавалась эта порода в условиях кочевого скотоводства при круглогодовом содержании животных на пастбищах в исключительно суровом континентальном климате степей. Снежные бураны и гололедица нередко приводили к гибели значительной части животных. Естественный и искусственный отбор действовали в направлении сохранения животных, способных откладывать в запас много жира и вместе с тем крепких и выносливых. Благодаря этому калмыцкий скот в благоприятных кормовых условиях откладывает на внутренних органах и под кожей много жира, который затем зимой используется как резерв питательных веществ. Помимо этого, жир предохраняет животных от холода.
Масть калмыцкого скота преимущественно красная. Однако встречаются животные красные с белой головой, красно-пестрые, белохребтовые, а также с белыми отметинами на нижней части туловища. Голова у скота горбоносая, легкая с коротким лбом, рога расположены вертикально вверх и изогнуты в виде полумесяцев, затылочный гребень вдавлен Шея у животных средней длины, мускулистая, туловище компактное, широкое, с хорошо развитой и крепкой мускулатурой. Однако у части животных отмечается недостаточное развитие зада (рис. 33).
В породе выделяют животных двух типов: мясного скороспелого и мясного позднеспелого. Животные первого типа быстрее заканчивают развитие, имеют более легкий костяк и более высокий убойный выход (на 2-4%), однако они несколько мельче мясного позднеспелого типа. Телята при рождении имеют массу 22-25 кг и к отъему (к 8 месяцам) достигают 200-220 кг. Масса коров - 420-500 кг, быков - 750-850 кг. При интенсивном выращивании на мясо молодняк в 16-18-месячном возрасте весит 350-450 кг. Убойный выход составляет 55-60%, а хорошо откормленных животных - до 68% .
Рис 33 Бык калмыцкой породы
Вопрос 3. Откорм скота.
Успех откорма зависит от уровня, типа и качества кормления, породных особенностей, возраста животных и их скороспелости. Быстрее и лучше… Так, молодняк до года на 1 кг прироста живой массы затрачивает 5-6 корм. ед.,… Большое значение имеет и продолжительность откорма. Наиболее выгоден короткий интенсивный откорм, особенно молодняка,…Нагул скота
Для успешного проведения нагула необходимо сформировать гурты одновозрастного скота. В степной зоне гурты составляют примерно из 150 голов… В зависимости от зональных условий, вида и качества пастбищ в расчете на одно… За 100-120-дневный нагул живая масса полновозрастного скота увеличивается обычно на 30-40%, а живая масса молодняка за…Тема 12. Свиноводство
1.Биологические особенности свиней.
2. Основные породы свиней (крупная белая, ландрас, беркширская).
3. Типы откорма свиней. Факторы, влияющие на результаты откорма свиней и качество свинины.
Вопрос 1. Биологические особенности свиней.
Все породы по направлению продуктивности делят на три группы. К 1-й наиболее многочисленной, группе относят свиней мясосального (универсального)… Для воспроизводства желательно выбирать свинок универсального направления…Основные показатели продуктивности свиней
Вопрос 2. Основные породы свиней (крупная белая, ландрас, беркширская). В настоящее время в стране разводят 22 породы и 3 типа свиней. Основная из них - крупная белая, удельный вес которой…Рис. 1 - Крупная белая
Кожа плотная, эластичная, без складок. Щетина тонкая, гладкая, густо покрывает все туловище. Живая масса взрослых хряков 300-360 кг, свиноматок - 220-260 кг. Длина туловища у хряков 178-183 см, у свиноматок 160-165 см. Многоплодие маток 10-12 поросят при средней массе их 1,2-1,3 кг, молочность (масса гнезда в 21 день) 48-50 кг. К 2-месячному возрасту, масса 1 поросенка составляет 16-18 кг и более.
При интенсивном откорме молодняк в возрасте 6-7 месяцев достигает живой массы 100-110 кг при затрате на 1 кг прироста 4-4,5 корм, единицы. Животные хорошо используют пастбища, а также сочные корма до 30 % от общей питательности рационов.
Порода ландрас. Выведена в Дании в результате скрещивания местной датской свиньи с крупной белой породой. Животные белой масти, с большими свисающими ушами, обладают длинным, относительно узким туловищем, широкими, хорошо развитыми окороками. Среднесуточный прирост молодняка на откорме составляет 700-750 г при затратах на 1 кг прироста 4,0-4,1 корм, единицы, выход мяса в тушах -58-59 %.
Свиноматки достаточно многоплодны - в среднем 10-11 поросят на опорос. Животные этой узкоспециализированной беконной породы довольно требовательны к условиям содержания и кормления. Тем не менее, порода ландрас широко распространена по всей территории России и используется для скрещивания в качестве отцовской формы.
Беркширские, или средние черные, свиньи принадлежат к одной из самых старых пород Англии. Произошли эти свиньи от простых позднеспелых свиней в графстве Беркшир, имевших красновато-бурый цвет с черными пятнами. Сначала они улучшались путем разведения «в себе», но уже начиная с XVIII века, их стали скрещивать, с неаполитанскими, а затем китайскими и английскими мелкими черными свиньями. В результате этих скрещиваний тип прежних беркширских свиней существенно изменился, и они приобрели округлость форм, скороспелость, хорошую способность к откорму и прекрасные вкусовые качества мяса. Беркширы относятся к мясо-сальному типу свиней, но они пригодны в молодом возрасте и для получения высокого качества бекона. Густая и мягкая щетина современных беркширов имеет черную окраску, с белыми отметинами на концах ног, морды и на хвосте. Кожа тонкая без складок. Голова легкая и скорее короткая, с широким лбом, с умеренно вогнутым профилем и с средней величины ушами, несколько наклоненными вперед. Ноги средней длины, тонкие, но крепкие и широко расставленные. Благодаря своей бодрости, подвижности и крепкой конституции беркширы хорошо используют пастбище и высоко оплачивают корм. По сравнению с белыми породами средние черные лучше переносят жару на пастбище и вообще для южных районов с жарким климат являются более подходящими.
Беркширы приобрели некоторую популярность в дореволюционной России главным образом в юго-западных районах, за их скороспелость, превосходное качество «мраморного» (с прослойкой сала) мяса, хорошее здоровье и приспособленность к жаркому климату.
Беркширские свиньи устойчиво передают потомству свои полезные качества при скрещивании с местными простыми свиньями, давая скороспелый приплод, ценный для мясо-сальных и для беконных целей.
В настоящее время их в России мало. Чистопородные беркширы до последней войны разводились в отдельных хозяйствах Московской области, а также в Ростовской области, в Краснодарском крае, в УССР и единично встречались в некоторых учебно-опытных хозяйствах зоотехнических вузов и техникумов.
В СССР беркширские свиньи использовались для выведения многих новых пород и породных групп свиней (миргородской, каликинской и северокавказской пород, молдавской, кемеровской, подольской и др. породных групп). В настоящее время беркширы используются для промышленного скрещивания с другими породами. При этом для скрещивания используют хряков беркширской породы, а маток крупной белой, украинской степной белой и других пород.

Рис. 206. Хряк и матка беркширскои породы.
Разводимые в России беркширские свиньи по своим размерам некрупные. Живой вес хряков в трехлетнем возрасте бывает 200-250 кг, длина туловища 140-145 см, обхват груди обычно равняется длине туловища.
У наиболее крупных хряков живой вес достигает 260-280 кг. Матки беркширской породы имеют живой вес 160-180 кг, длину туловища 130-135 см, обхват груди 130-132 см. Плодовитость, как это отмечено выше, 8-9 поросят на опорос, вес помёта в 30-дневном возрасте 50-60 кг. Лучшие взрослые матки имеют вес 225-250 кг, при длине туловища 145-150 см и обхвате груди 150-160 см. Плодовитость лучших маток также бывает 10-11 поросят на опорос, а молочность - 60-70 кг.
Вопрос 3. Типы откорма свиней. Факторы, влияющие на результаты откорма свиней и качество свинины.
У поросенка должен быть правильный прикус: нижняя и верхняя челюсти равномерные по длине и ширине, а зубные поверхности легко и равномерно… Поросенок должен иметь хороший аппетит, а корм захватывать и пережевывать, а… У скороспелых свиней голова короткая с прогнутым профилем рыла, широким лбом. Уши тонкие, небольшие, прямостоячие или…Виды откорма
Для откорма свиней до мясных кондиций используют картофель, сахарную свеклу, комбинированный силос и пищевые отходы. Их содержание в рационе доводят… Беконный откорм. Это - особый вид мясного откорма, при котором к качеству… Беконом называют свинину, полученную от молодых животных и приготовленную в виде специально разделенных и особым…Факторы, определяющие эффективность откорма
Откорм помесных свиней, полученных в результате скрещивания двух заводских пород, при полноценном кормлении дает лучшие результаты, чем откорм… Здоровье. Только здоровые, конституционально крепкие животные имеют высокую… Чаще всего свиньи заболевают в подсосный и полеотъемный период, поэтому необходимо особенно в это время выполнять все…Влияние кормов на качество свинины
Первая группа. Это зерновые корма, способствующие получению свинины высокого качества - ячмень, пшеница, рожь, горох, люпин, просо. Из сочных кормов… из зеленых - люцерна, клевер, эспарцет, вико-овсяные и горохо-овсяные смеси;… Вторая группа. Это корма, при скармливании которых качество свинины несколько снижается (сало мягкое, рыхлая невкусная…Живая масса свиней, снимаемых с откорма
Однако, как показала практика, откорм свиней до более высокой живой массы (120-130 кг) экономически выгоднее, но для этого на откорм необходимо… Убойный выход у свиней изменяется в зависимости от их массы при убое. Так, при… Однако при расчете эффективности откорма до разной живой массы необходимо учитывать все затраты, произведенные на…Правила убоя свиней и первичная переработка свинины
Если подвесить свинью не представляется возможным, ее оглушают ударом кувалды в лоб и затем ножом перерезают кровеносные сосуды. Кровь свиней имеет… Обескровливание свиней продолжается 5-6 минут. Если имеется в виду… Опаливать щетину лучше лампой паяльной или газовой горелкой, после чего для придания салу специфического приятного…Тема 13. Овцеводство
1.Биологические особенности овец.
2. Основные породы овец тонкорунного (советский меринос, асканийская, прекос), полутонкорунного (цигайская, линконьнская, куйбышевская, горьковская) полугрубошерстного (сараджинска, гаджикская) и грубошерстного (романовская, каракульская, гиссарская) направлений продуктивности.
3. Откорм овец.
Вопрос 1. Биологические особенности овец
Значительное место в мясном балансе занимает баранина. Одна из ценных ее особенностей - наименьшее содержание холестерина по… Примечание: Употребляют в пищу и конское мясо. Конина является незаменимым компонентом высших сортов колбас. У ряда…Воспроизводство стада
За полтора месяца до начала случки маткам желательно создать оптимальные условия кормления и содержания. К этому времени у них должны быть отняты… В условиях хозяйства на 15-20 голов, целесообразно иметь одного… При приближении родов овцематки стремятся найти укромное место, блеют и проявляют сильное беспокойство. Иногда,…Вопрос 2. Основные породы овец
Тонкорунное направление продуктивности Советский меринос(шерстно-мясная, тонкорунная). Порода имеет сложное… Племенная работа велась на получение животных, сочетающих ценные качества шерсти новокавказских мериносов (длина,…Рис 55 - Баран цигайской породы
Цигайские овцы отличаются высокой скороспелостью и хорошими мясными качествами: убойный выход составляет 55%, масса ягнят ко времени отбивки достигает до 28-30 кг.
Из шкур овец этой породы, убитых в молодом возрасте, готовят меха, из которых шьют цигейковые шубы.
В Молдавии и ряде Балканских стран цигайских овец доят. От матки, помимо того, что она кормит ягнят, получают по 40-60 кг молока. Плодовитость составляет 120-130 ягнят на 100 маток. Овец этой породы разводят в основном в Крыму, на юге Украины, Молдавии, в Ростовской и Саратовской областях, Краснодарском крае и Казахстане.
Полутонкорунные мясо-шерстные породы овец. Эти породы в основном были выведены в Англии. Их подразделяют на две группы: длинношерстные и короткошерстные. Из длинношерстных пород в России разводят линкольнскую, ромни-марш, куйбышевскую. северокавказскую мясо-шерстную и др.; из короткошерстных - шропширскую, гемпширскую, горьковскую, латвийскую темноголовую, литовскую черноголовую и ряд других.
Линкольнская порода. Выведена более 200 лет назад в Англии в графстве Линкольн путем скрещивания мясных грубошерстных овец, отличавшихся крупным ростом, с длинношерстными лейстерскими баранами. Порода создавалась в условиях почти круглогодичного содержания животных на богатых пастбищах.
Линкольнская порода овец - одна из крупнейших в мире. Масса баранов достигает 130-160 кг, маток - 80-90 кг. На выставке в Англии был представлен баран массой около 300 кг. Наряду с большой массой и скороспелостью линкольны отличаются высокой шерстной продуктивностью. С баранов настригают по 9-10 кг шерсти, с маток - по 6-6,5 кг. Шерсть линкольнов в основном 36-44-го качества, длина волокна 25-30 см с правильной волнистостью и шелковистым блеском (люстрой). Выход чистой шерсти 65-70%.
Туловище у овец длинное, широкое и глубокое, ляжки сильно развиты. Ягнята очень быстро растут: масса туши в 4-месячном возрасте 25-28 кг. Однако линкольны плохо акклиматизируются в сухом жарком климате, требовательны к условиям кормления и содержания.
Полугрубошерстное направление продуктивности
Сараджинская порода(с полугрубой шерстью). Выведена методом народной селекции в Туркмении, где производили отбор местных курдючных овец с большим содержание в шерсти пуха.
Овцы этой породы имеют крепкую конституцию, хорошо выраженные мясо-сальные качества, хвост жирный в виде слабо подтянутого курдюка. Живая масса небольшая - у баранов 80-90 кг у маток 55-60 кг. Животные скороспелы (у ягнят от рождения до отбивки среднесуточный прирост достигает 200 г. Шерстяная продуктивность относительно высокая - годовой настриг шерсти у баранов 4-5 кг, у маток 3-4 кг при выходе мытой шерсти 55-60%.
Шерсть в основном белого цвета, состоит из большого количества длинного руна (75-85%), переходною волоса (13-15%) и не слишком грубой шерсти (6%).
Масть новорожденных ягнят коричневая с различными оттенками. К годовалому возрасту они становятся белыми, за исключением головы и ног.
Таджикская порода выведена (1948-1963 гг.) в экспериментальном хозяйстве Научно-исследовательского института животноводства Таджикской ССР. При создании породы применяли сложное воспроизводительное скрещивание баранов сараджинской породы с матками гиссарской породы. Начиная с первого поколения, помесей желательного типа разводили «в себе». Кроме того, использовались линкольн-гиссарские помесные матки, которых спаривали с сараджино-гиссарскими баранами. Из полученного потомства баранчиков выбраковывали, а ярок включали в основное стадо сараджино-гиссарских маток.
Овцы таджикской породы сочетают высокие мясо-сальные качества материнской (гиссарской) породы с хорошими показателями полугрубой шерсти, свойственной отцовской (сараджинской) породе. По телосложению, крепости конституции, а также по скороспелости таджикские овцы весьма сходны с гиссарскими. Они, подобно гиссарским, крупные, с хорошо развитым курдюком. Бараны весят 120-130 кг, матки - 70-80 кг. Шерсть таджикских овец, как и у сараджинских, полугрубая, белого цвета, с хорошим блеском, средняя длина косиц при весенней стрижке у баранов равна 20 см, длина пуха - 10 см, у маток соответственно 16 и 7,5 см. Настриг шерсти с баранов составляет 4,85 кг, с маток- 2,9-3,1 кг при выходе мытой шерсти 68-72 %. По совокупности признаков и свойств шерсть овец таджикской породы - ценное сырье для изготовления ковров.
Грубошерстное направление продуктивности
Романовская порода (грубошерстные, шубные овцы). Отличительные черты - из них получаются лучшие в мире овчины. Овцы этой породы сравнительно некрупные - бараны весят 60-65 кг, матки - 45-50 кг. Годовой настриг шерсти за 3-4 кратную стрижку - у баранов - 2-2,5 кг, у маток - 1,5-1,8 кг. Овец отличает большая плодовитость (до 300%) и полиэстричность (за один год матки могут ягниться 2 раза в год или 3 раза в два года). Различают 3 типа романовских овец: грубый, нежный и нормальный.
Овцы грубого типа характеризуются грубым телосложением, сильно развитыми рогами у баранов и наличием рогов у маток. Цвет шерсти у этого типа темный, что обусловлено большим содержанием ости (соотношение ости и пуха 164) и значительно большой длиной ости по отношению к пуху. Косицы шерсти почти прямые. У баранов они образуют гриву. Мездра у овчин толстая, грубая, что делает ее тяжелой.
Нежный тип овец имеет утонченный костяк, формы туловища узкие. Кожа тонкая. Бараны и матки комолы. У шерсти содержится иного пуха в 10 раз больше, чем ости, причем, пух всегда перерастает ость. Овчины очень красивые, светло - голубого цвета, но не прочные в носке, т.к шерсть на них быстро сваливаегся. Овцы нормального типа имеют крепкую конституцию, туловище бочкообразное. Голова небольшая, сухая, с небольшой горбоносостью (особенно у баранов). Цвет кроющего волоса на голове черный с широкой белой полосой. Животные бывают рогатые и комолые.
В шерсти содержание пуха 2 раза больше, чем ости, и пух перерастает ость. Благодаря этому шерсть не сваливается, имеет красивый серый цвет с голубоватым оттенком. Самые хорошие овчины получаются их ягнят 4-6 месячного возраста. Однако так как животные в это время продолжают расти, то забивают их в возрасте 8-9 месяцев, когда ость после первой стрижки отрастает на 2,5-З см, а пух - на 4-6 см.
Каракульская порода(смушково-молочная). Одна из самых ценных пород смушкового направления (самые лучшие шапки, воротники, пальто и др). Ценятся смушки за оригинальную извитость шерстного покрова, которая придает им исключительно красивый рисунок.
Порода ценится еще за сычуг - сырье для изготовления сычужного фермента и лекарственных препаратов.
Особенность породы - многообразие типов овец: по окраске, расцветкам, смушковым и конституционным свойствам. Главный критерий разделения - окраска (масть) овец, которая является не только экономически обоснованной, но и коррелятивно связана с конституциональными особенностями овец. Главный, наиболее многочисленный тип - черные овцы. Есть серый тип, бухарский, сурхандарьинский, каракалпакский сур и даже розовой окраски. Различают породные типы и заводские типы (выведенные в заводах).
Убой ягнят производят в 1 или 2 день после рождения. Животные не крупные - масса баранов - до 70 кг, маток - до 46 кг.
От взрослых особей еще получают шерсть, которая является ценным сырьем для текстильной, ковровой, валяльной промышленности; мясо за счет выбраковки из стада; молоко от маток, у которых ягнят направили на смушку (до 30 л от одной матки за сезон).
Гиссарские овцы (мясо-сальные). В значительной степени это обособившаяся группа курдючных овец (Локайская долина Таджикистана).
Гиссарские овцы самые крупные из всех пород. И самые выносливые. Легко совершают 400-500 км - переходы с высокогорных пастбищ Альп. Масть черная или темно-рыжая.
Они имеют сильный, крепкий костяк, длинное, широкое и глубокое туловище, и очень сильно развитый, приподнятый курдюк. Голова массивная, горбоносая, уши длинные, свислые, шея короткая, мускулистая. Живая масса баранов 130-140 кг, маток - 70-80 кг. Убойный выход овец - 58-60%. Курдюк до 23-30 кг и более.
Молодняк исключительно скороспелый: при отбивке баранчики весят 47-50 кг, ярки -46-48 кг.
При хороших мясо-сальных качествах гиссарские овцы дают очень мало шерсти, причем, самой грубой из всех отродий курдючных овец: средний настриг шерсти у баранов - 1,3-1,6 кг, у маток 1-1,3 кг., в шерсти много ости и мертвого волоса (до 34%).
Выбор породы
Если хозяин хочет получить высококачественную баранину, а шерсть его интересует меньше, и он ее намерен сдавать в заготконтору, то этим целям в… При отборе овец любой породы для разведения следует обращать внимание на…Вопрос 3. Откорм овец
Важной отраслью продуктивного животноводства является овцеводство. По количеству пород и разнообразию продукции оно превосходит другие отрасли.… Подсчитано, что на протяжении всей истории цивилизации сохранилось…Кормление и уход за овцами
Летом, напротив, овец выпускают на пастбища рано. Пастьба на росистой (но не бобовой) траве в летние дни не вредна, а, наоборот, полезна, так как,… Летом пастьбу можно начинать с восхода солнца и продолжать до 11 часов дня, а… В весенний и летний период и очень важно не допускать пастьбы овец после дождя или по обильной росе на участках…Кормление овцематок
В нашей области ягнение проходит зимой и весной. Зимнее ягнение предпочтительно проводить в том случае, если имеется достаточно кормов высокого… Приблизительно через час после ягнения у овцематки появляется жажда, поэтому… К осени ранние ягнята обычно имеют высокую живую массу, нагуливаясь на дешевых кормах. Сверхремонтный молодняк после…Тема 14. Птицеводство
1. Биологические особенности птицы.
2. Основные породы кур яичного (леггорн), мясного (корниш, плимутрок) и яично-мясного (род-айланд, нью-гемпшир, московская черная) направлений продуктивности.
3. Породы уток (пекинская), гусей (холмогорская), индеек (северокавказская, бронзовая широкогрудая).
4. Производство мяса птицы.
Вопрос 1. Биологические особенности птицы. Выращивание.
Куры яичных пород весят 1,7-2,2, петухи - 2,5-3 кг; мясо-яичных - соответственно 2,3-2,8 и 3,4-3,8 кг; мясных - 3,0-3,5 и 3,8-4,5 кг. Половая… Инстинкт насиживания у кур яичных пород утрачен, у мясо-яичных и мясных… Продолжительность жизни кур 10-12 лет.Продукция птицеводства
Яйцо - один из наиболее ценных пищевых продуктов. По питательности 1 яйцо равняется 40 г мяса или 120 г молока. Оно обеспечивает на 5% суточную… Мясо домашней птицы отличается высокими диетическими и кулинарными качествами,… Перо и пух птицы - незаменимое сырье для изготовления перин и подушек.Разведение птицы и выращивание молодняка
Выведенному молодняку надо дать обсохнуть под наседкой, а затем отсадить в коробку в теплое место с температурой 26-28°С до окончания вывода.… Инкубировать куриные яйца необходимо в следующем режиме: 1-2-й день -… В промышленных инкубаторах всё автоматизировано.Выращивание цыплят-бройлеров
В приусадебных хозяйствах цыплят обычно выращивают на мясо двумя способами. Первый (интенсивный), когда молодняк выращивают в любое время года,… На 1 м2 площади пола размещают 12 цыплят в суточном возрасте. Такая плотность… При выращивании мясных цыплят (бройлеров) в помещении поддерживают влажность в пределах 60-70% и температуру: в 1-5…Содержание кур-несушек
Практикой определены оптимальные сроки использования птицы: для кур яичных пород - 12 месяцев; мясо-яичных- 10; мясных - 8 месяцев.
Если в хозяйстве нет условий для выгула птицы и ограничены размеры помещения, целесообразно содержать кур в клетках. Для этих целей можно использовать деревянные или металлические конструкции. Пол в клетках делают решетчатыми из проволоки толщиной 3-5 мм, размер ячейки 2,5х5,0 см. Устанавливают его с уклоном 7-9° к передней стенке. Для сбора помета под пол на расстоянии 10-12 см устанавливают противни из плоского шифера или оцинкованной жести. Двери в клетках делают из металлических или деревянных прутьев с интервалом 8-10 см. Кормушку изготавливают из дерева или жести и крепят с наружной стороны клетки так, чтобы под ней могло прокатиться яйцо. Поилку размещают снаружи или внутри клетки с передней или задней стороны. Высота клетки внутри 40-45 и 60-65 см при совместном содержании кур и петухов. На 1 м2 площади пола можно содержать 15 яичных или 10 мясных кур. Клеточные несушки более требовательны к полноценному кормлению, чем куры на выгульном содержании. Особенно они нуждаются в витамине D, которым птица обеспечивается на выгулах при облучении солнцем.
Условия содержания домашней птицы изменяются в зависимости от времени года и климатических особенностей. Летом птица весь день находится на свежем воздухе, вволю поедает свежую зелень, богатую витаминами. Зимой, когда птица все время находится в помещении, благоприятные условия приходится создавать искусственным путем.
При этом важно, чтобы птичник был заранее хорошо подготовлен к зимнему содержанию птицы. Помещение следует отремонтировать, заделать все щели в стенах, крыше, побелить, привести в порядок оборудование и инвентарь.
Зимой нужно заботиться о том, чтобы птица как можно больше находилась на воздухе. При наружной температуре минус 10° кур, уток, индеек и гусей желательно выпускать на выгулы, очищенные от снега и застланные подстилкой. Утки, гуси и индейки могут некоторое время переносить температуру до 20° мороза без ущерба для продуктивности.
Принудительно выгонять птицу на выгул не следует. Зимой в солярий можно привлечь птицу кормушками с зерном, которые следует ставить на расчищенную от снега площадку. Если недалеко от птичника есть водоем, рекомендуется во время оттепели делать в нем проруби и выпускать в них гусей и уток. Вокруг проруби нужно положить подстилку.
Содержание уток
Разводить уток в домашнем хозяйстве очень выгодно. В короткий срок (55 дней) утки достигают 2,4 кг веса. Утиное мясо калорийно, с прекрасными вкусовыми качествами. Утки хорошо оплачивают корма: на 1 кг привеса молодняка затраты корма составляют 3-4 кг. Уток можно выращивать в помещениях, на водоемах и на сухопутных выгулах. Лучше всего выращивать уток до 55 дней. В возрасте 60-70 дней у них начинается линька (смена пера), которая обычно сопровождается потерей веса, а при обработке тушки невозможно удалить появившиеся пеньки, что несколько снижает качество тушки.
Утки относятся к всеядным птицам, так как поедают разнообразные корма растительного и животного происхождения. Они легко находят корма в водоемах и на сухих выгулах. Утки отличаются высокой жизнеспособностью, они устойчивы к инфекционным заболеваниям. Сохранность молодняка утят самая высокая по сравнению с цыплятами, гусятами и индюшатами.
Особенно удобно разводить уток в домашнем хозяйстве, если вблизи имеется водоем. В водоемах живет много мелких насекомых, богатая растительность, которая служит для уток хорошей пищей. Умелое использование естественных водоемов позволяет сократить расход концентрированных кормов для уток на 30-40%.
При отсутствии водных выгулов уток можно содержать круглогодично в помещениях. В помещении для выращивания уток поддерживают температуру около 18°С. В первые дни жизни утята особенно чувствительны к колебаниям температуры. С 30-дневного возраста утят можно выращивать в неотапливаемых помещениях даже в осенне-зимний период. Особенно чувствительны утята к колебаниям температуры в первые дни жизни. Плохо переносят утята и чрезмерную плотность посадки. Максимальная плотность посадки утят на 1 м2 составляет с 1 до 15-дневного возраста - 16-20 голов, с 16 до 30-дневного возраста - 12-14 голов, а с 31 до 55-дневного возраста - 6-8 голов.
В качестве подстилки для утят до 10-дневного возраста лучше использовать древесную стружку или измельченную солому. Стелят ее два раза в сутки - утром и вечером. В первые 3-4 дня не следует применять в качестве подстилочного материала опилки, так как молодняк клюет их и заболевает.
Продолжительность светового дня в первые дни жизни должна быть 14-16 часов, чтобы утята успевали съесть данный им корм. Помещения, где содержатся утята, необходимо хорошо вентилировать, не допускать скопления вредных газов и влаги. Летом утят с 10-дневного возраста начинают выпускать на выгул, сначала ненадолго, а затем время увеличивают до полного светового дня. На выгулах устраивают теневые навесы для защиты утят от солнца и дождя. Зимой утят тоже можно выпускать на выгул, начиная с 30-дневного возраста, при температуре воздуха не ниже - КУС. На водоем молодняк выпускают в 20-дневном возрасте, температура воды должна быть не менее 14,198С.
Выращивание гусей
В птичнике необходимо иметь хорошую вентиляцию, влажность воздуха поддерживается в пределах 65-75%. В первую неделю выращивания рекомендуется… Молодняк выпускают на выгул в теплую солнечную погоду. Первые пять дней - на… В домашних хозяйствах широко распространено выращивание гусят с использованием естественных пастбищ. Этот способ…Содержание индеек
Разводить индеек в домашнем хозяйстве исключительно интересно и очень выгодно. Если птицевод-любитель приобрел весной 50-60 индюшат, то к концу лета он получит дополнительно 300- 350 кг индюшиного мяса, которое отличается высокой питательностью. Для этого вида птицы свойственна высокая интенсивность роста. Если в суточном возрасте средний вес индюшат составляет 60 г, то к годовалому возрасту он достигает 12 кг, т. е. увеличивается в 200 раз. Вес других видов птицы увеличивается к этому возрасту в 60-100 раз. Индюшат можно вывести в собственном хозяйстве и дорастить под наседкой или приобрести подрощенными 20-30-дневного возраста, что существенно облегчит их выращивание.
К выгулу индюшат можно приучать постепенно с 10 дня. Первый раз в солярий их выпускают на 10-15 минут, затем время пребывания их в солярии увеличивают. К 15 дню молодняк весь день может находиться на воздухе. С 20 дня его выпускают на пастбища, прилегающие к птичнику. В первые дни индюшат кормят с фанерных лотков. В поилки лучше наливать теплую воду. С 45 дня молодняк приучают к насестам, что помогает предотвратить скученность индюшат в ночное время.
При интенсивном методе выращивания в 120-дневном возрасте индюшата весят 4 кг и более. При этом на 1 кг привеса затрачивают 3,5 кг корма. Если молодняк выращивают с использованием выпаса, срок откорма увеличивается до 180 дней.
Кормление птицы
Основу рационов домашней птицы составляют зерновые корма (55-75%). Из злаковых зерновых наиболее широко используют кукурузу, ячмень, пшеницу… Зимой домашней птице хорошо скармливать проращенное зерно ячменя или овса.… Дрожжуют корма для увеличения содержания в них витаминов группы В. При дрожжевании на 1 кг сухой смеси из молотых…Кормление молодняка птицы
Для кормления гусят в первые дни жизни готовят увлажненные мещанки из вареных яиц, зерномучнистых кормов (ячменя, овса, пшена, пшеничных отрубей),… С 45-60-дневного возраста при пастьбе на богатых кормами выгулах (сады после… Индюшата хорошо поедают лук, что способствует лучшему перевариванию кормов и служит профилактической мерой против…Кормление кур
Взрослую птицу кормят 3-4 раза в сутки. Утром дают зерновую смесь, днем - один или два раза влаженку, вечером - сухое зерно. Сухая мучная смесь… Не следует перекармливать картофелем переярых кур, особенно мясо-яичных пород,… В кормушке для минеральных кормов всегда должны быть ракушка или мел, древесная зола, гравий, в поилках - свежая и…Кормление гусей
Для кормления гусят в первые дни жизни готовят увлажненные мешанки из вареных яиц, зерномучнистых кормов (ячменя, овса, пшена, пшеничных отрубей),… В зимний период гусятам можно скармливать в значительном количестве грубые,…Кормление уток
Сухую мучную смесь утки едят плохо и много ее рассыпают, поэтому мешанка должна быть влажной, но рассыпчатой. Кормить уток в зимний период надо 3…Кормление индеек
Рацион для индеек должен состоять из зерно-мучных кормов, сочной зелени, картофеля, пекарских или кормовых дрожжей, ракушки, костной муки и… В зимний период индеек следует кормить 3 раза в день: утром и днем скармливают… Вопрос 2. Основные породы кур яичного (леггорн), мясного (корниш, плимутрок) и яично-мясиого(род-айланд, нью-гемпшир,…Яичные породы
Леггорн -порода кур яичного направления. Выведена в 19 в. в США скрещиванием итальянских белых кур с минорками, испанскими, бойцовскими и другими…Кроссы яичных пород
Кросс Ломан-Браун (коричневый) – цыплята достигают половой зрелости в 135 дней, когда у них появляется первое яйцо. Уже в 150 дней яйценоскость… Кросс Тетра СЛ. Высокопродуктивный. Средняя яйценоскость за 52 недели… Изобраун – результат многолетний работы французских селекционеров. Широко распространен в мире. Хорошо…Мясные породы
Корниш (Корнуэльские куры). Порода кур мясного направления. Выведена в Великобритании (графство Корнуолл) в 19 в. скрещиванием бойцовских кур старой… Плимутрок белый, полосатый(англ. Plymouth Rock), порода кур мясо-яичного…Яично-мясные породы
Род-айланд(род–айленд), порода кур мясо-яичного направления. Выведена в США (штаты Род-Айленд и Массачусетс) во 2-й пол. 19 в. скрещиванием местных… Нью-гемпшир -порода кур мясо-яичного направления. Выведена в США (штат… Московскаяпорода кур мясо-яичного направления. Выведена в хозяйствах Московской области скрещиванием юрловских кур с…Декоративные куры
К этой группе кур принадлежат породы с оригинальной постановкой тела, рожковидным гребнем, хохлатые и карликовые. Из карликовых кур наиболее распространена порода бентам.
Вопрос 3. Породы уток (пекинская), гусей(холмогорская), индеек(северокавказская, бронзовая широкогрудая).
Породы уток
Средняя живая масса уток пекинской породы - 3 кг, селезней - 3,5 кг. Утята к 55-дневному возрасту достигают живой массы 2,5 кг. Яйцекладку утки… Птица имеет белое с желтоватым оттенком оперение, длинное, слегка приподнятое… В 1971 году в нашу страну завезены из Англии утки пекинской породы двухлинейного кросса «Черри-Велли». Утки этого…Породы гусей
Туловище гусей этой породы широкое, массивное с кожной складкой на животе. У белой разновидности клюв крепкий, довольно сильно изогнутый,…Породы индеек
Бронзовые широкогрудые.Выведены в США. Туловище массивное, широкое спереди, к хвосту постепенно суживается; голова небольшая; грудь очень широкая,…Вопрос 4. Производство мяса птицы.
Бройлер (англ. Broiler, от broil - жарить на огне), мясной цыпленок, отличающийся интенсивным ростом, скороспелостью, низкими затратами корма,…Убой птицы, обработка и хранение тушек
Убитых кур и индеек ощипывают сразу же после убоя, пока тушки еще теплые. Гусей и уток лучше ощипывать после предварительного охлаждения тушек в… Ощипывать индеек, кур и цыплят легче и быстрее, когда тушки убитой птицы… При полупотрошении вскрывают брюшную полость тушки и извлекают весь кишечник от зоба до прямой кишки. При потрошении…А) Основная
2. Вракин В.Ф., Сидорова М.В. Морфология сельскохозяйственных животных. М.: Агропромиздат, 1991.- 528 с. 3. Писменская В.Н., Ленченко Е.М. Гистология. Функциональная морфология… 4. Яров И.И., Васютенкова Н.С. Основы животноводства: Учебник. М.:Агропромиздат,1986.-335 с.Б) Дополнительная
2. Алмазов И.Е., Сутулов Л.С. Атлас по гистологии и эмбриологии: Учебное пособие. М.: Медицина, 1978.- 544 с. 3. Антипчук Ю.П. Гистология с основами эмбриологии: Учебное пособие. М.:… 4. Животноводство: Учебник/ Под редакцией Е.А. Арзуманяна. М.: Колос, 1976.- 436 с.МОДУЛЬ I
1. Введение
2. Общая характеристика строения и функций системы органон движения.
3. Система внутренних органов и органов крово- и лимфообращения.
4. Нервная система, железы внутренней секреции и анализаторы.
5. Анатомия домашней птицы.
МОДУЛЬ II
6. Клетки и ткани как основные структурные элементы организма животного.
7. Мышечная ткань.
8. Кожа и её производные. Особенности строения некоторых органов пищеварения.
МОДУЛЬ III
9. Выведение и разведение животных.
10. Кормление животных.
11. Скотоводство.
12. Свиноводство.
13. Овцеводство.
14. Птицеводство.